
Aerosolized Hypertonic Saline Hinders Biofilm Formation to Enhance Antibiotic Susceptibility of Multidrug-Resistant Acinetobacter baumannii

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
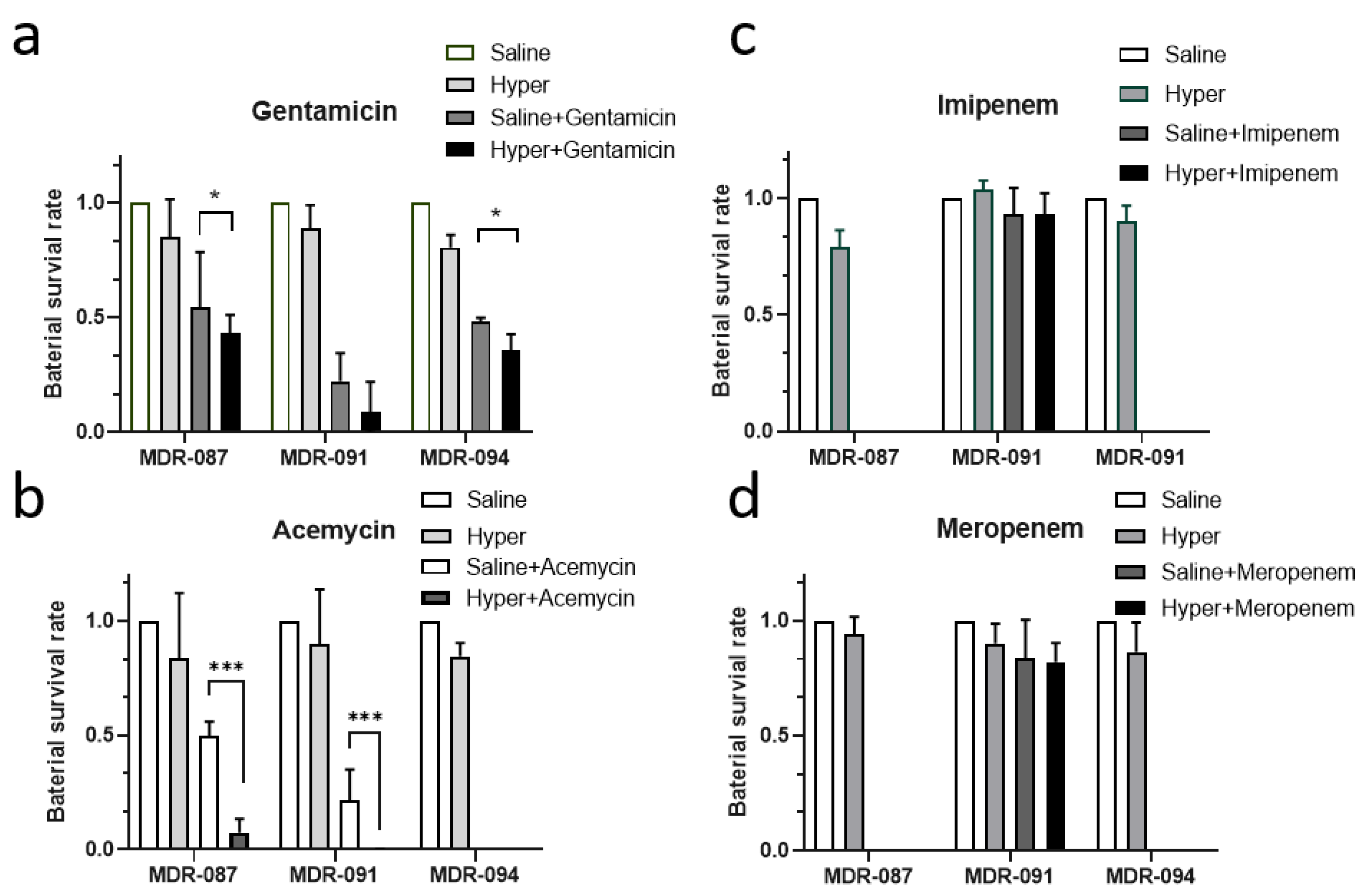{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Culture and Drug Resistance Test
2.2. Aerosol Generation and Aerosol Particle Size Distribution
2.3. Biofilm Quantitative Measurement and Scanning Electron Microscopy
2.4. The Spatial Distribution of Protein and Carbohydrate in Biofilm
2.5. The Dynamic of Bacteria Swarming
2.6. Statistical Analysis
3. Results and Discussion
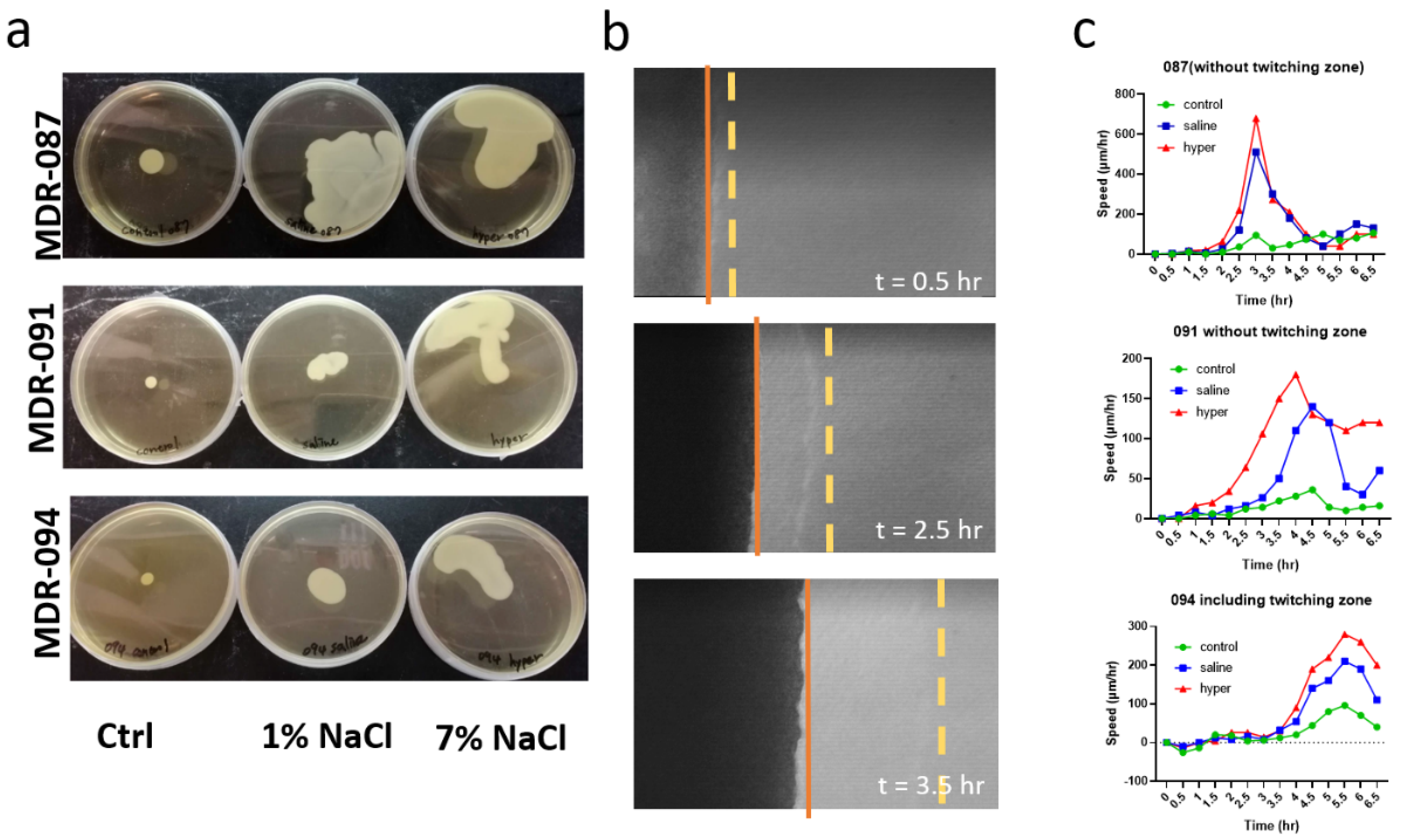
3.1. Hypertonic Saline Hinders the Biofilm Formation of A. baumannii
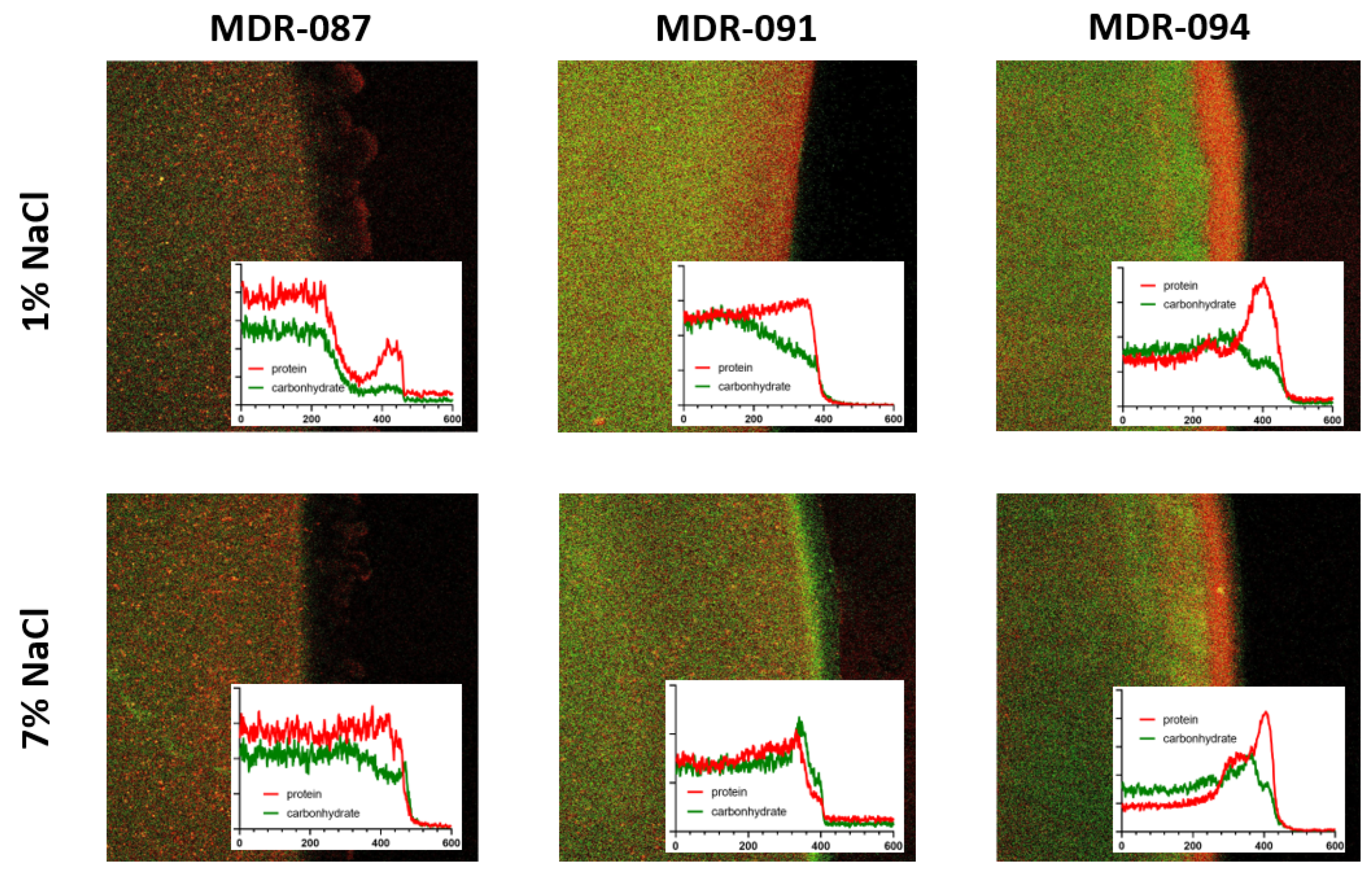
3.2. Hypertonic Saline Stimulates Bacterial Swarming and Alters the Distribution of EPS
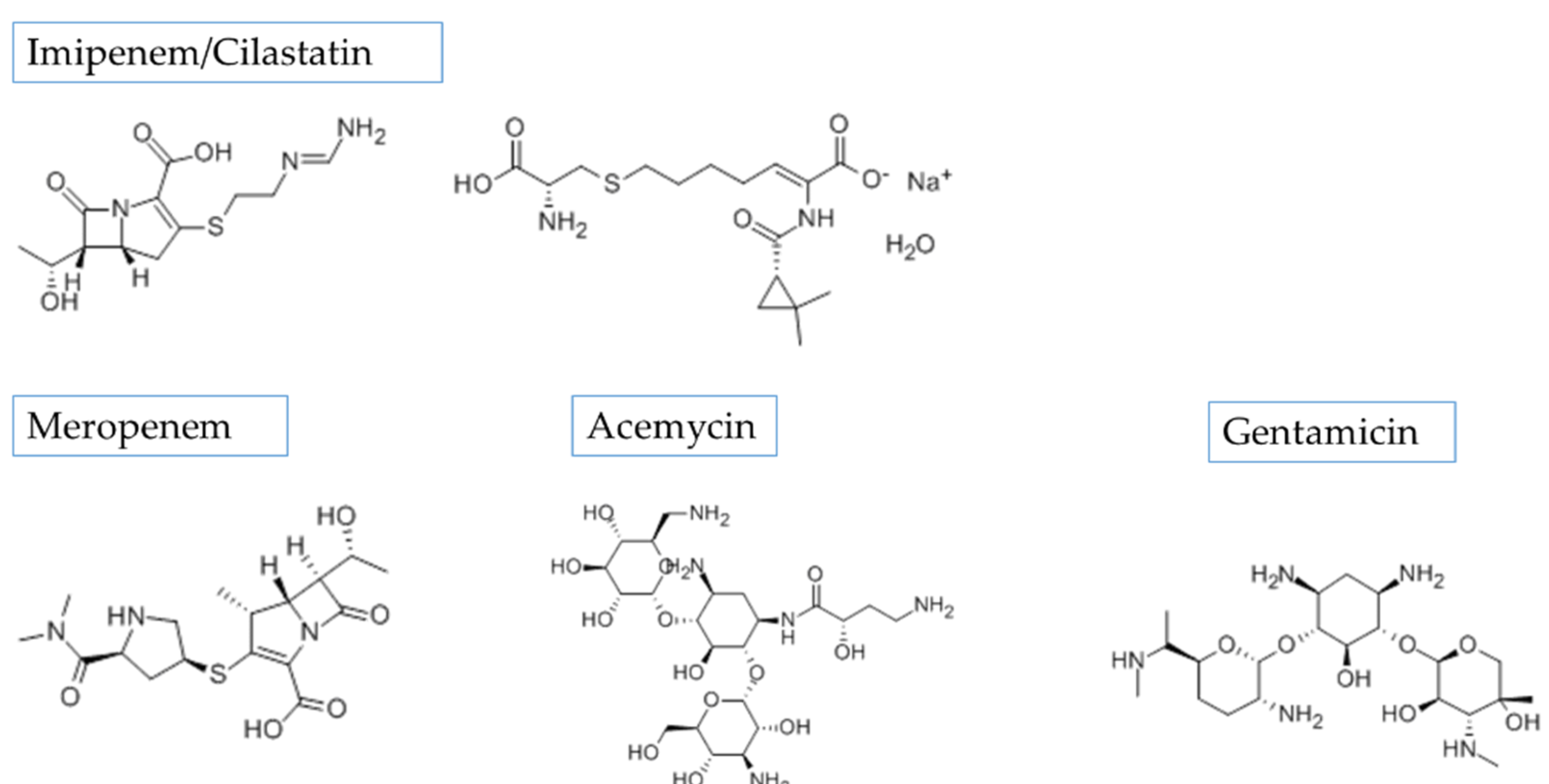
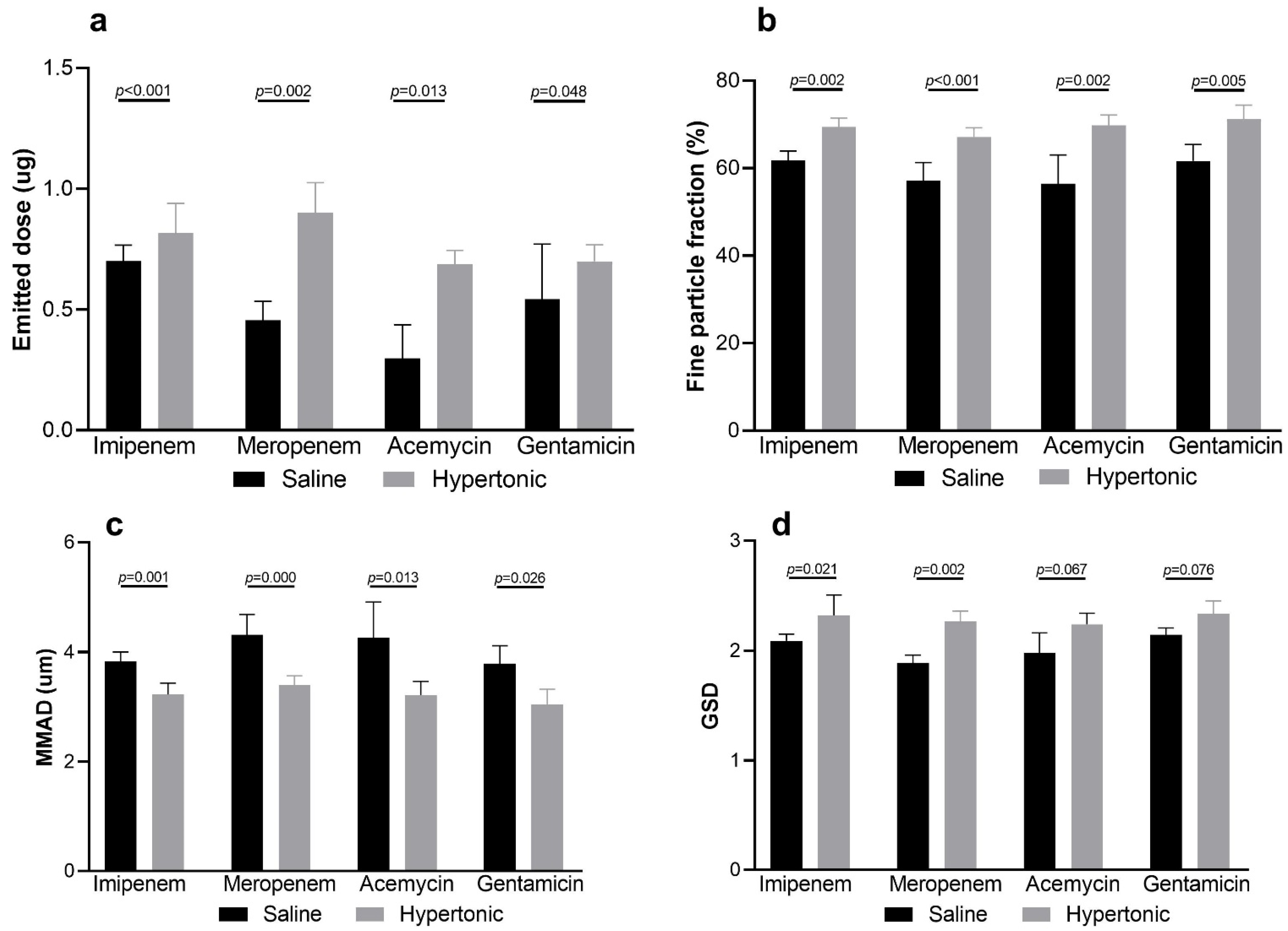
3.3. Influence of Hypertonic Saline to Aerosol Characteristics and Delivered Dose
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Laessig, K.A. End points in hospital-acquired pneumonia and/or ventilator-associated pneumonia clinical trials: Food and drug administration perspective. Clin. Infect. Dis. 2010, 51 (Suppl. 1), S117–S119. [Google Scholar] [CrossRef]
- Moubareck, C.A.; Halat, D.H. Insights into Acinetobacter baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Cillóniz, C.; Dominedò, C.; Torres, A. Multidrug Resistant Gram-Negative Bacteria in Community-Acquired Pneumonia. Crit. Care 2019, 23, 79. [Google Scholar] [CrossRef] [Green Version]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Kurihara, M.N.L.; Sales, R.O.; Silva, K.E.D.; Maciel, W.G.; Simionatto, S. Multidrug-resistant Acinetobacter baumannii outbreaks: A global problem in healthcare settings. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200248. [Google Scholar] [CrossRef]
- Skariyachan, S.; Taskeen, N.; Ganta, M.; Krishna, B.V. Recent perspectives on the virulent factors and treatment options for multidrug-resistant Acinetobacter baumannii. Crit. Rev. Microbiol. 2019, 45, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.C.; Swanson, J.M. An Update on Aerosolized Antibiotics for Treating Hospital-Acquired and Ventilator-Associated Pneumonia in Adults. Ann. Pharmacother. 2017, 51, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Flume, P.A.; VanDevanter, D.R. Clinical applications of pulmonary delivery of antibiotics. Adv. Drug Deliv. Rev. 2015, 85, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.T.; Leung, S.S.; Tang, P.; Parumasivam, T.; Loh, Z.H.; Chan, H.K. Inhaled formulations and pulmonary drug delivery systems for respiratory infections. Adv. Drug Deliv. Rev. 2015, 85, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Wenzler, E.; Fraidenburg, D.R.; Scardina, T.; Danziger, L.H. Inhaled Antibiotics for Gram-Negative Respiratory Infections. Clin. Microbiol. Rev. 2016, 29, 581–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gappa, M.; Steinkamp, G.; Tümmler, B.; von der Hardt, H. Long-term tobramycin aerosol therapy of chronic Pseudomonas aeruginosa infection in patients with cystic fibrosis. Scand. J. Gastroenterol. Suppl. 1988, 143, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Fang, W.F.; Kao, H.C.; Chen, H.-C.; Tsai, Y.-C.; Shen, L.-S.; Li, C.-L.; Chang, H.-C.; Huang, K.-T.; Lin, M.-C.; et al. Influencing factors of successful eradication of multidrug-resistant Acinetobacter baumannii in the respiratory tract with aerosolized colistin. Biomed. J. 2014, 37, 314–320. [Google Scholar]
- Lu, Q.; Luo, R.; Bodin, L.; Yang, J.; Zahr, N.; Aubry, A.; Golmard, J.-L.; Rouby, J.-J.; Nebulized Antibiotics Study Group. Efficacy of high-dose nebulized colistin in ventilator-associated pneumonia caused by multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Anesthesiology 2012, 117, 1335–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badave, G.K.; Kulkarni, D. Biofilm Producing Multidrug Resistant Acinetobacter baumannii: An Emerging Challenge. J. Clin. Diagn. Res. 2015, 9, DC08–DC10. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Lee, H.W.; Koh, Y.M.; Kim, J.; Lee, J.-C.; Lee, Y.-C.; Seol, S.-Y.; Cho, D.-T. Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm and adhere to epithelial cell surfaces. Clin. Microbiol. Infect. 2008, 14, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B. Approaches to Prevention, Removal and Killing of Biofilms; International Biodeterioration & Biodegradation: London, UK, 2003; Volume 51, pp. 249–253. [Google Scholar]
- Zapotoczna, M.; O’Neill, E.; O’Gara, J.P. Untangling the Diverse and Redundant Mechanisms of Staphylococcus aureus Biofilm Formation. PLoS Pathog. 2016, 12, e1005671. [Google Scholar] [CrossRef]
- Chen, C.S.; Shiu, R.F.; Hsieh, Y.Y.; Xu, C.; Vazquez, C.I.; Cui, Y.; Hsu, I.C.; Quigg, A.; Santschi, P.H.; Chin, W.-C. Stickiness of extracellular polymeric substances on different surfaces via magnetic tweezers. Sci. Total Environ. 2021, 757, 143766. [Google Scholar] [CrossRef]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef] [Green Version]
- Tsuneda, S.; Aikawa, H.; Hayashi, H.; Yuasa, A.; Hirata, A. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface. FEMS Microbiol. Lett. 2003, 223, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.S.; Furrer, J.M.; Kadilak, A.L.; Hinestroza, H.F.; Gage, D.; Cho, Y.K.; Shor, L.M. Bacterial Extracellular Polymeric Substances Amplify Water Content Variability at the Pore Scale. Front. Environ. Sci. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.L.; Chiu, L.C.; Wan, G.H.; Huang, C.-C.; Lee, Z.-T.; Lin, Y.-T.; Wu, S.-R.; Chen, C.-S. Hypertonic saline enhances the efficacy of aerosolized gentamicin against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 4325. [Google Scholar] [CrossRef] [PubMed]
- Harimawan, A.; Rajasekar, A.; Ting, Y.P. Bacteria attachment to surfaces–AFM force spectroscopy and physicochemical analyses. J. Colloid Interface Sci. 2011, 364, 213–218, Erratum in 2013, 15, 392–470. [Google Scholar] [CrossRef] [PubMed]
- Klemmer, A.; Krämer, I.; Kamin, W. Physicochemical compatibility of nebulizable drug admixtures containing budesonide and colistimethate or hypertonic saline. Int. J. Pharm. Compd. 2013, 17, 254–261. [Google Scholar]
- Carro, L.M.; Martínez-García, M.A. Use of Hyaluronic Acid (HA) in Chronic Airway Diseases. Cells 2020, 9, 2210. [Google Scholar] [CrossRef]
- Reeves, E.P.; Williamson, M.; O’Neill, S.J.; Greally, P.; McElvaney, N.G. Nebulized hypertonic saline decreases IL-8 in sputum of patients with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2011, 183, 1517–1523. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.-L.; Chiang, C.-E.; Lin, M.-C.; Kau, M.-L.; Lin, Y.-T.; Chen, C.-S. Aerosolized Hypertonic Saline Hinders Biofilm Formation to Enhance Antibiotic Susceptibility of Multidrug-Resistant Acinetobacter baumannii. Antibiotics 2021, 10, 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091115
Lin H-L, Chiang C-E, Lin M-C, Kau M-L, Lin Y-T, Chen C-S. Aerosolized Hypertonic Saline Hinders Biofilm Formation to Enhance Antibiotic Susceptibility of Multidrug-Resistant Acinetobacter baumannii. Antibiotics. 2021; 10(9):1115. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091115
Chicago/Turabian StyleLin, Hui-Ling, Chen-En Chiang, Mei-Chun Lin, Mei-Lan Kau, Yun-Tzu Lin, and Chi-Shuo Chen. 2021. "Aerosolized Hypertonic Saline Hinders Biofilm Formation to Enhance Antibiotic Susceptibility of Multidrug-Resistant Acinetobacter baumannii" Antibiotics 10, no. 9: 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091115