
Whole-Genome Characterisation of ESBL-Producing E. coli Isolated from Drinking Water and Dog Faeces from Rural Andean Households in Peru

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Setting and Design
2.2. Households’ Characteristics
2.3. Microbiological Identification and Antimicrobial Susceptibility Testing
2.4. Phenotypical and Molecular Identification of ESBL-Producing Ability
2.5. Genome Sequencing and Bioinformatic Analysis
3. Results
3.1. Microbiological Identification and Antimicrobial Susceptibility Testing of the E. coli Isolates
3.2. Detection of ESBL-Encoding Genes by PCR and Whole-Genome Sequencing Analysis (WGS)
3.3. Pathogenicity, Virulence Genes, and Type of Plasmids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- D’Agostino, M.; Cook, N. Foodborne Pathogens. In Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 83–86. [Google Scholar]
- Guentzel, M.N. Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacter, and Proteus. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2021, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance-the Need for Global Solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- Bonelli, R.R.; Moreira, B.M.; Picão, R.C. Antimicrobial Resistance among Enterobacteriaceae in South America: History, Current Dissemination Status and Associated Socioeconomic Factors. Drug Resist. Updat. 2014, 17, 24–36. [Google Scholar] [CrossRef]
- Mendes Oliveira, V.R.; Paiva, M.C.; Lima, W.G. Plasmid-Mediated Colistin Resistance in Latin America and Caribbean: A Systematic Review. Travel Med. Infect. Dis. 2019, 31, 101459. [Google Scholar] [CrossRef]
- Rapoport, M.; Faccone, D.; Pasteran, F.; Ceriana, P.; Albornoz, E.; Petroni, A.; Corso, A.; Maldonado, M.L.; Lucero, C.; Amalfa, F.; et al. First Description of Mcr-1-Mediated Colistin Resistance in Human Infections Caused by Escherichia coli in Latin America. Antimicrob. Agents Chemother. 2016, 60, 4412–4413. [Google Scholar] [CrossRef] [Green Version]
- Medina-Pizzali, M.L.; Hartinger, S.M.; Salmon-Mulanovich, G.; Larson, A.; Riveros, M.; Mäusezahl, D. Antimicrobial Resistance in Rural Settings in Latin America: A Scoping Review with a One Health Lens. Int. J. Environ. Res. Public Health 2021, 18, 9837. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global Trends in Antimicrobial Resistance in Animals in Low- And Middle-Income Countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental Antimicrobial Resistance and Its Drivers: A Potential Threat to Public Health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar] [CrossRef]
- Larson, A.; Hartinger, S.M.; Riveros, M.; Salmon-Mulanovich, G.; Hattendorf, J.; Verastegui, H.; Huaylinos, M.L.; Mäusezahl, D. Antibiotic-Resistant Escherichia coli in Drinking Water Samples from Rural Andean Households in Cajamarca, Peru. Am. J. Trop. Med. Hyg. 2019, 100, 1363–1368. [Google Scholar] [CrossRef]
- Pitout, J.D.D. Extraintestinal Pathogenic Escherichia coli: An Update on Antimicrobial Resistance, Laboratory Diagnosis and Treatment. Expert Rev. Anti Infect. Ther. 2012, 10, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Sawa, T.; Kooguchi, K.; Moriyama, K. Molecular Diversity of Extended-Spectrum β-Lactamases and Carbapenemases, and Antimicrobial Resistance. J. Intensive Care 2020, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Mohamudha, P.R.; Harish, B.N.; Parija, S.C. AmpC Beta Lactamases among Gram Negative Clinical Isolates from a Tertiary Hospital, South India. Braz. J. Microbiol. 2010, 41, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colquechagua Aliaga, F.; Andrade Sevilla, C.; Escalante Gonzales, E. Extended-Spectrum Beta-Lactamase (ESBL)-producing Enterobacteriaceae in Fecal Samples at the National Institute of Child Health, Peru. Rev. Peru Med. Exp. Salud Publica 2015, 32, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loyola, S.; Gutierrez, L.R.; Horna, G.; Petersen, K.; Agapito, J.; Osada, J.; Rios, P.; Lescano, A.G.; Tamariz, J. Extended-Spectrum β-Lactamase–Producing Enterobacteriaceae in Cell Phones of Health Care Workers from Peruvian Pediatric and Neonatal Intensive Care Units. Am. J. Infect. Control 2016, 44, 910–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, C.; Hinostroza, N.; Astocondor, L.; Ochoa, T.; Jacobs, J. Characterization of ESBL-Producing Salmonella Enterica Serovar Infantis Infection in Humans, Lima, Peru. Am. J. Trop. Med. Hyg. 2019, 101, 746–748. [Google Scholar] [CrossRef]
- León-Luna, D.; Fajardo-Loyola, A.; Yareta-Yareta, J.; Burgos-Espejo, A.; Peralta-Siesquen, C.; Galarza-Pérez, M.; Marcos-Carbajal, P. Molecular Characterization of Multi-Resistant Enterobacteria in Two Departments of the Peruvian Jungle. Biomedica 2021, 41, 180–187. [Google Scholar] [CrossRef]
- Benavides, J.A.; Shiva, C.; Virhuez, M.; Tello, C.; Appelgren, A.; Vendrell, J.; Solassol, J.; Godreuil, S.; Streicker, D.G. Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in Common Vampire Bats Desmodus Rotundus and Livestock in Peru. Zoonoses Public Health 2018, 65, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Hartinger, S.M.; Medina-Pizzali, M.L.; Salmon-Mulanovich, G.; Larson, A.J.; Pinedo-Bardales, M.; Verastegui, H.; Riberos, M.; Mäusezahl, D. Antimicrobial Resistance in Humans, Animals, Water and Household Environs in Rural Andean Peru: Exploring Dissemination Pathways through the One Health Lens. Int. J. Environ. Res. Public Health 2021, 18, 4604. [Google Scholar] [CrossRef]
- Quino, W.; Hurtado, C.V.; Escalante-Maldonado, O.; Flores-León, D.; Mestanza, O.; Vences-Rosales, F.; Zamudio, M.L.; Gavilán, R.G. Multidrug Resistance of Salmonella infantis in Peru: A Study through next Generation Sequencing. Rev. Peru Med. Exp. Salud Publica 2019, 36, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavides, J.A.; Godreuil, S.; Opazo-Capurro, A.; Mahamat, O.O.; Falcon, N.; Oravcova, K.; Streicker, D.G.; Shiva, C. Long-Term Maintenance of Multidrug-Resistant Escherichia coli Carried by Vampire Bats and Shared with Livestock in Peru. Sci. Total Environ. 2022, 810, 152045. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Salvatierra, G.; Dávila-Barclay, A.; Ayzanoa, B.; Castillo-Vilcahuaman, C.; Huang, M.; Pajuelo, M.J.; Lescano, A.G.; Cabrera, L.; Calderón, M.; et al. Market Chickens as a Source of Antibiotic-Resistant Escherichia coli in a Peri-Urban Community in Lima, Peru. Front. Microbiol. 2021, 12, 635871. [Google Scholar] [CrossRef] [PubMed]
- Clinical Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Lezameta, L.; Gonzáles-Escalante, E.; Tamariz, J.H. Comparison of four phenotypic methods to detect Extended-Spectrum Betalactamases. Rev. Peru Med. Exp. Salud Publica 2010, 27, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 28 February 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, F.M.; de Carli, S.; Lopes, C.E.; Machado, L.; Vieira, T.R.; Pöppl, G.; Cardoso, M.R.I.; Zaha, A. Non-Lactose-Fermenting Uropathogenic Escherichia coli from Dogs: Virulence Profile Characterization. Lett. Appl. Microbiol. 2021, 72, 596–603. [Google Scholar] [CrossRef]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Burián, K. Differential Epidemiology and Antibiotic Resistance of Lactose-Fermenting and Non-Fermenting Escherichia coli: Is It Just a Matter of Taste? Biol. Futur. 2020, 71, 175–182. [Google Scholar] [CrossRef]
- Food and Drug Administration. Chapter 4A: Diarrheagenic Escherichia coli. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-4a-diarrheagenic-escherichia-coli (accessed on 1 January 2022).
- Ori, E.L.; Takagi, E.H.; Andrade, T.S.; Miguel, B.T.; Cergole-Novella, M.C.; Guth, B.E.C.; Hernandes, R.T.; Dias, R.C.B.; Pinheiro, S.R.S.; Camargo, C.H.; et al. Diarrhoeagenic Escherichia coli and Escherichia Albertii in Brazil: Pathotypes and Serotypes over a 6-Year Period of Surveillance. Epidemiol. Infect. 2019, 147, e10. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; van der Ploeg, C.; Rogé, A.; Licona-Moreno, D.; Delgado, G.; Morales-Espinosa, R.; Cravioto, A.; Eslava, C. Diversity of Potentially Pathogenic Escherichia coli O104 and O9 Serogroups Isolated before 2011 from Fecal Samples from Children from Different Geographic Regions. Microorganisms 2021, 9, 2227. [Google Scholar] [CrossRef]
- Spanu, T.; Sanguinetti, M.; Tumbarello, M.; D’Inzeo, T.; Fiori, B.; Posteraro, B.; Santangelo, R.; Cauda, R.; Fadda, G. Evaluation of the New VITEK 2 Extended-Spectrum Beta-Lactamase (ESBL) Test for Rapid Detection of ESBL Production in Enterobacteriaceae Isolates. J. Clin. Microbiol. 2006, 44, 3257–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, I.; Geiss, H.K.; Mack, D.; Stürenburg, E.; Seifert, H. Detection of Extended-Spectrum Beta-Lactamases among Enterobacteriaceae by Use of Semiautomated Microbiology Systems and Manual Detection Procedures. J. Clin. Microbiol. 2007, 45, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Condamine, B.; Dion, S.; Gordon, D.M.; Denamur, E. The E Phylogroup of Escherichia coli Is Highly Diverse and Mimics the Whole E. Coli Species Population Structure. Environ. Microbiol. 2021, 23, 7139–7151. [Google Scholar] [CrossRef] [PubMed]
- De Castro Stoppe, N.; Silva, J.S.; Carlos, C.; Sato, M.I.Z.; Saraiva, A.M.; Ottoboni, L.M.M.; Torres, T.T. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front. Microbiol. 2017, 8, 2512. [Google Scholar] [CrossRef] [Green Version]
- Coura, F.M.; Diniz, S.D.A.; Silva, M.X.; Mussi, J.M.S.; Barbosa, S.M.; Lage, A.P.; Heinemann, M.B. Phylogenetic Group Determination of Escherichia coli Isolated from Animals Samples. Sci. World J. 2015, 2015, 258424. [Google Scholar] [CrossRef] [Green Version]
- Alberca, V.; León, D.; Falcón, N. Keeping Backyard Animals and Evaluation of Knowledge and Practices Associated with Exposure to Zoonotic Agents in La Coipa, Cajamarca, Peru. Rev. Inv. Vet. Perú 2020, 31, e18733. [Google Scholar] [CrossRef]
- Benavides, J.A.; Streicker, D.G.; Gonzales, M.S.; Rojas-Paniagua, E.; Shiva, C. Knowledge and Use of Antibiotics among Low-Income Small-Scale Farmers of Peru. Prev. Vet. Med. 2021, 189, 105287. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. OIE List Of Antimicrobial Agents Of Veterinary Importance; World Organisation for Animal Health: Paris, France, 2007. [Google Scholar]
- Tanfous, F.B.; Alonso, C.A.; Achour, W.; Ruiz-Ripa, L.; Torres, C.; Hassen, A.B. First Description of KPC-2-Producing Escherichia coli and ST15 OXA-48-Positive Klebsiella Pneumoniae in Tunisia. Microb. Drug Resist. 2017, 23, 365–375. [Google Scholar] [CrossRef]
- Beyrouthy, R.; Robin, F.; Dabboussi, F.; Mallat, H.; Hamzé, M.; Bonnet, R. Carbapenemase and Virulence Factors of Enterobacteriaceae in North Lebanon between 2008 and 2012: Evolution via Endemic Spread of OXA-48. J. Antimicrob. Chemother. 2014, 69, 2699–2705. [Google Scholar] [CrossRef]
- Peng, Z.; Hu, Z.; Li, Z.; Zhang, X.; Jia, C.; Li, T.; Dai, M.; Tan, C.; Xu, Z.; Wu, B.; et al. Antimicrobial Resistance and Population Genomics of Multidrug-Resistant Escherichia coli in Pig Farms in Mainland China. Nat. Commun. 2022, 13, 1116. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli Isolates That Carry Vat, Fyua, Chua, and Yfcv Efficiently Colonize the Urinary Tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales-Rodriguez, A.O.; Varillas, S.F.I.; Pastor, H.J.B.; Mitma, Y.L.; Canales, D.H.; Chero, P.A.W.; Gutierrez, C.; Cunza, S.S. Immunological and Biochemical Response from Older Adults with Urinary Tract Infection to Uropathogenic Escherichia coli Virulence Factors. Rev. Peru Med. Exp. Salud Publica 2020, 37, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Berberov, E.M.; Freeling, J.; He, D.; Moxley, R.A.; Francis, D.H. Significance of Heat-Stable and Heat-Labile Enterotoxins in Porcine Colibacillosis in an Additive Model for Pathogenicity Studies. Infect. Immun. 2006, 74, 3107–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, S.; Luo, J.; Li, X.; Zhuo, C.; Wu, A.; Chen, X.; Huang, L.S. Molecular Epidemiology and Characteristics of CTX-M-55 Extended-Spectrum β-Lactamase-Producing Escherichia coli From Guangzhou, China. Front. Microbiol. 2021, 12, 730012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Ding, X.M.; Lin, X.L.; Sun, R.Y.; Lu, Y.W.; Cai, R.M.; Webber, M.A.; Ding, H.Z.; Jiang, H.X. The Emergence of Chromosomally Located BlaTX-M-55 in Salmonella from Foodborne Animals in China. Front. Microbiol. 2019, 10, 1268. [Google Scholar] [CrossRef] [PubMed]
- Moffat, J.; Chalmers, G.; Reid-Smith, R.; Mulvey, M.R.; Agunos, A.; Calvert, J.; Cormier, A.; Ricker, N.; Weese, J.S.; Boerlin, P. Resistance to Extended-Spectrum Cephalosporins in Escherichia coli and Other Enterobacterales from Canadian Turkeys. PLoS ONE 2020, 15, e0236442. [Google Scholar] [CrossRef]
- Norizuki, C.; Wachino, J.I.; Suzuki, M.; Kawamura, K.; Nagano, N.; Kimura, K.; Arakawa, Y. Specific BlaCTX-M-8/IncI1 Plasmid Transfer among Genetically Diverse Escherichia coli Isolates between Humans and Chickens. Antimicrob. Agents Chemother. 2017, 61, e00663-17. [Google Scholar] [CrossRef] [Green Version]
- National Library of Medicine (US). Reference Gene Catalog; National Library of Medicine: Bethesda, MD, USA, 2022.
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517. [Google Scholar] [CrossRef]
- Adator, E.H.; Walker, M.; Narvaez-Bravo, C.; Zaheer, R.; Goji, N.; Cook, S.R.; Tymensen, L.; Hannon, S.J.; Church, D.; Booker, C.W.; et al. Whole Genome Sequencing Differentiates Presumptive Extended Spectrum Beta-Lactamase Producing Escherichia coli along Segments of the One Health Continuum. Microorganisms 2020, 8, 448. [Google Scholar] [CrossRef] [Green Version]
- Tekele, S.G.; Teklu, D.S.; Tullu, K.D.; Birru, S.K.; Legese, M.H. Extended-Spectrum Beta-Lactamase and AmpC Beta-Lactamases Producing Gram Negative Bacilli Isolated from Clinical Specimens at International Clinical Laboratories, Addis Ababa, Ethiopia. PLoS ONE 2020, 15, e0241984. [Google Scholar] [CrossRef]
- Guzmán-Blanco, M.; Labarca, J.A.; Villegas, M.V.; Gotuzzo, E. Extended Spectrum β-Lactamase Producers among Nosocomial Enterobacteriaceae in Latin America. Braz. J. Infect. Dis. 2014, 18, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuga, B.; Sellera, F.P.; Cerdeira, L.; Esposito, F.; Cardoso, B.; Fontana, H.; Moura, Q.; Cardenas-Arias, A.; Sano, E.; Ribas, R.M.; et al. WHO Critical Priority Escherichia coli as One Health Challenge for a Post-Pandemic Scenario: Genomic Surveillance and Analysis of Current Trends in Brazil. Microbiol. Spectr. 2022, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Escandón-Vargas, K.; Reyes, S.; Gutiérrez, S.; Villegas, M.V. The Epidemiology of Carbapenemases in Latin America and the Caribbean. Expert Rev. Anti Infect. Ther. 2017, 15, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, H.; Du, H. Carbapenemases in Enterobacteriaceae: Detection and Antimicrobial Therapy. Front. Microbiol. 2019, 10, 1823. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). CRE Technical Information. Available online: https://www.cdc.gov/hai/organisms/cre/technical-info.html (accessed on 19 February 2022).
- Temkin, E.; Adler, A.; Lerner, A.; Carmeli, Y. Carbapenem-Resistant Enterobacteriaceae: Biology, Epidemiology, and Management. Ann. NY Acad. Sci. 2014, 1323, 22–42. [Google Scholar] [CrossRef]
- Marimuthu, K.; Ng, O.T.; Cherng, B.P.Z.; Fong, R.K.C.; Pada, S.K.; De, P.P.; Ooi, S.T.; Smitasin, N.; Thoon, K.C.; Krishnan, P.U.; et al. Antecedent Carbapenem Exposure as a Risk Factor for Non-Carbapenemase-Producing Carbapenem-Resistant Enterobacteriaceae and Carbapenemase-Producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e00845-19. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Plasmids and the Spread of Resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance Plasmid Families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Ramsamy, Y.; Mlisana, K.P.; Amoako, D.G.; Abia, A.L.K.; Ismail, A.; Allam, M.; Mbanga, J.; Singh, R.; Essack, S.Y. Mobile Genetic Elements-Mediated Enterobacterales-Associated Carbapenemase Antibiotic Resistance Genes Propagation between the Environment and Humans: A One Health South African Study. Sci. Total Environ. 2022, 806, 150641. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [Green Version]

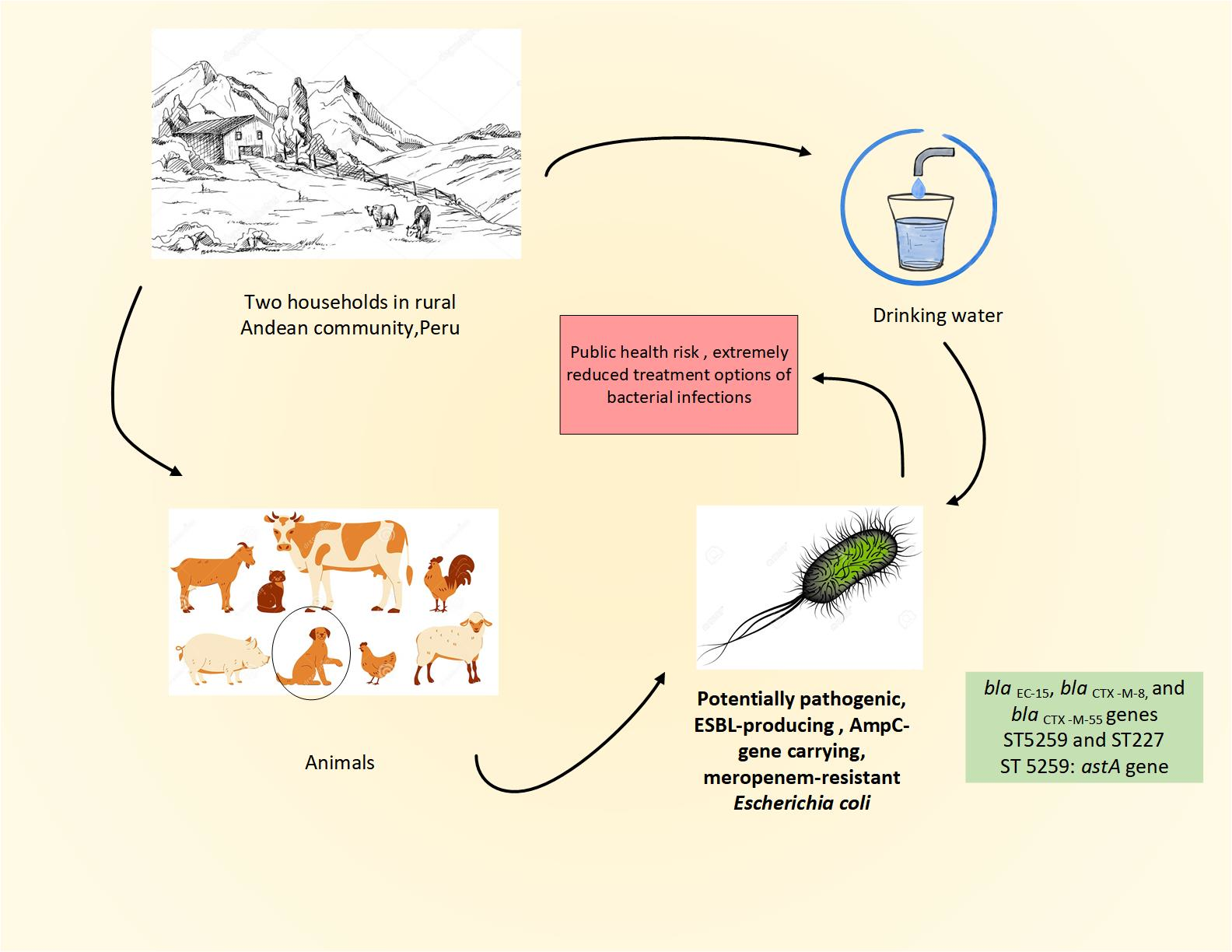{kind=link}
| ID E. coli Strains | Source | ST/CC | Serotype | Phylogroup | Virulence Genes | Resistance Phenotype | Resistance Genotype | Plasmid Type |
|---|---|---|---|---|---|---|---|---|
| 1143 | dog faeces | 5259/- | -:H46 | E | astA, chuA, cia, gad, ompT, terC | SAM, TZP, KZ, CXM, CTX, CAZ, FEP, MEM, SXT | aac(3)-IId, aph(3″)-Ib, aph(6)-Id, aadA1, aadA2, floR, blaTEM-1, blaEC-15, blaCTX-M-8, tet(M), tet(B) dfrA12 | IncI1-IAlpha, IncR, IncFIB(AP001918), IncFIC(FII) |
| 1144 | drinking water | 227/10 | O9:H10 | A | capU, cea, cia, fyuA, iutA, sitA, irp2, iroN, terC | SAM, TZP, KZ, CXM, CTX, MEM, CN, CIP | aph(3′)-IIa, cmlA1, blaCTX-M-55, blaEC-15, fosA3, qnrB19, sul3, tet(B) | IncFIB, Col440I, IncFII, IncI1-IAlpha, IncFII(pHN7A8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Pizzali, M.L.; Venkatesh, A.; Riveros, M.; Cuicapuza, D.; Salmon-Mulanovich, G.; Mäusezahl, D.; Hartinger, S.M. Whole-Genome Characterisation of ESBL-Producing E. coli Isolated from Drinking Water and Dog Faeces from Rural Andean Households in Peru. Antibiotics 2022, 11, 692. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050692
Medina-Pizzali ML, Venkatesh A, Riveros M, Cuicapuza D, Salmon-Mulanovich G, Mäusezahl D, Hartinger SM. Whole-Genome Characterisation of ESBL-Producing E. coli Isolated from Drinking Water and Dog Faeces from Rural Andean Households in Peru. Antibiotics. 2022; 11(5):692. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050692
Chicago/Turabian StyleMedina-Pizzali, Maria Luisa, Apoorva Venkatesh, Maribel Riveros, Diego Cuicapuza, Gabriela Salmon-Mulanovich, Daniel Mäusezahl, and Stella M. Hartinger. 2022. "Whole-Genome Characterisation of ESBL-Producing E. coli Isolated from Drinking Water and Dog Faeces from Rural Andean Households in Peru" Antibiotics 11, no. 5: 692. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11050692