In Vitro Synergism of Colistin and N-acetylcysteine against Stenotrophomonas maltophilia

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Colistin Susceptibility of S. maltophilia Strains Included in the Study
2.2. In Vitro Synergism of Colistin/NAC Combinations against S. maltophilia Strains Grown in Planktonic Phase
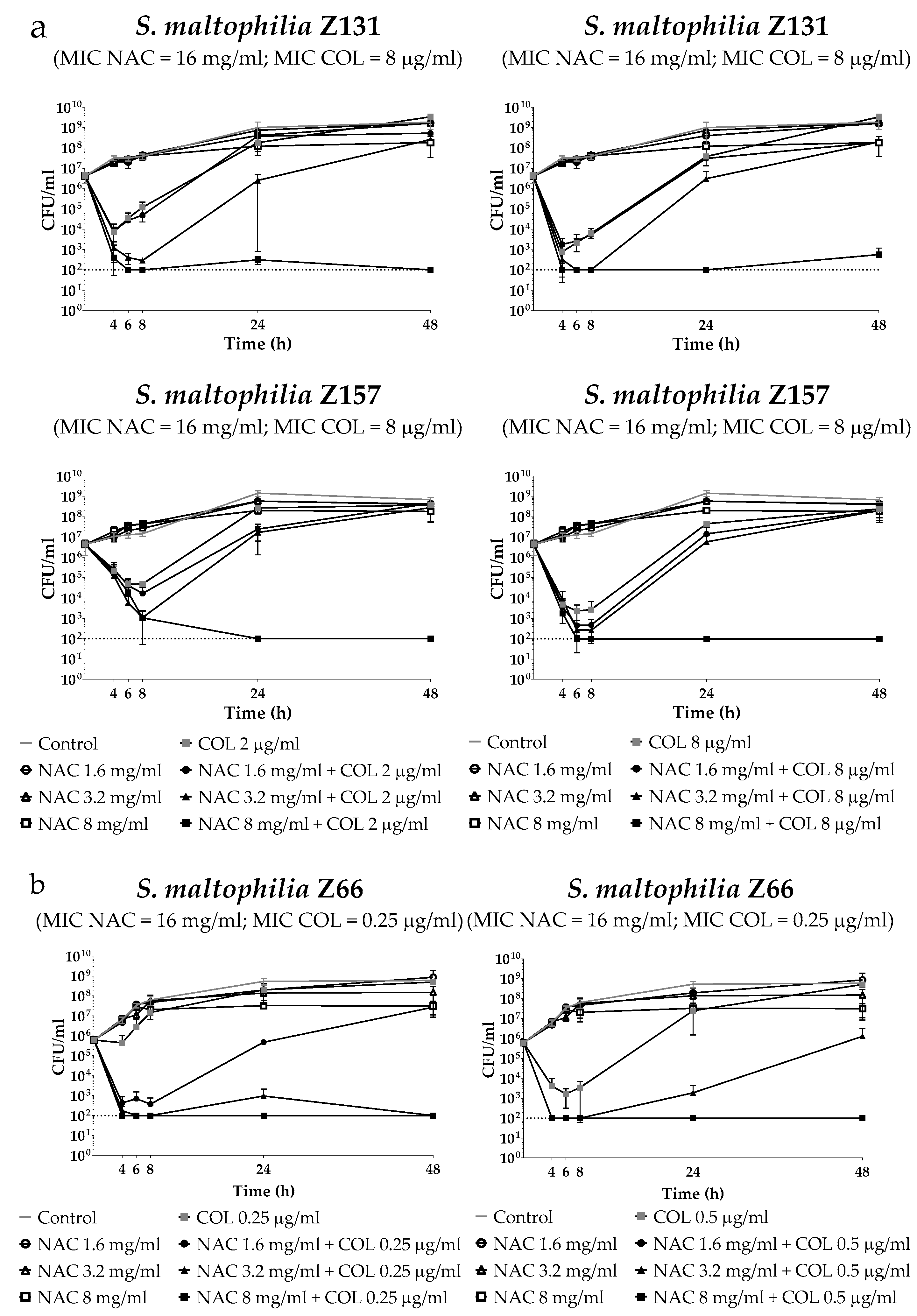
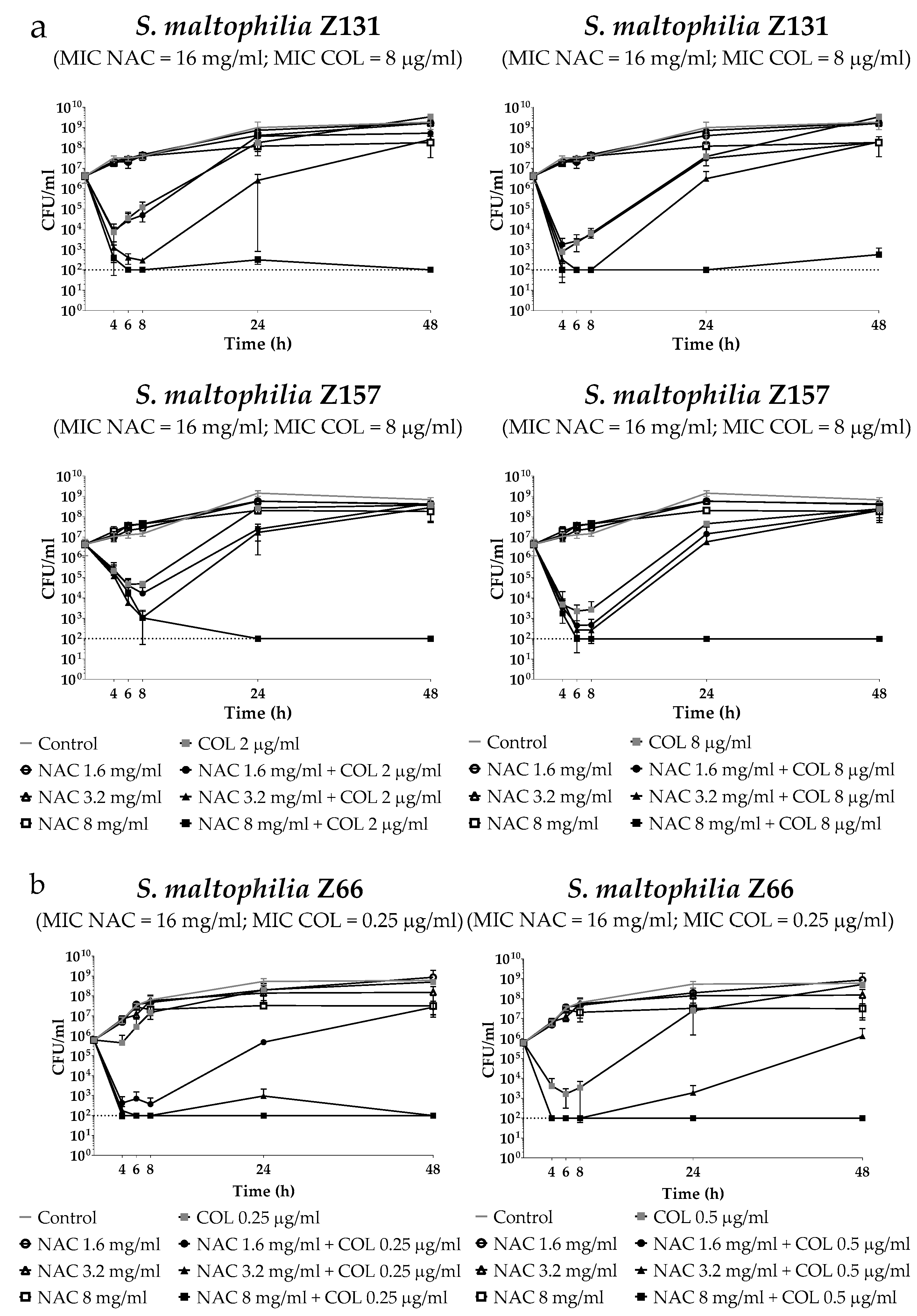
2.3. Time-Kill Assays of Colistin/NAC Combinations against Three Selected S. Maltophilia Strains Grown in Planktonic Phase
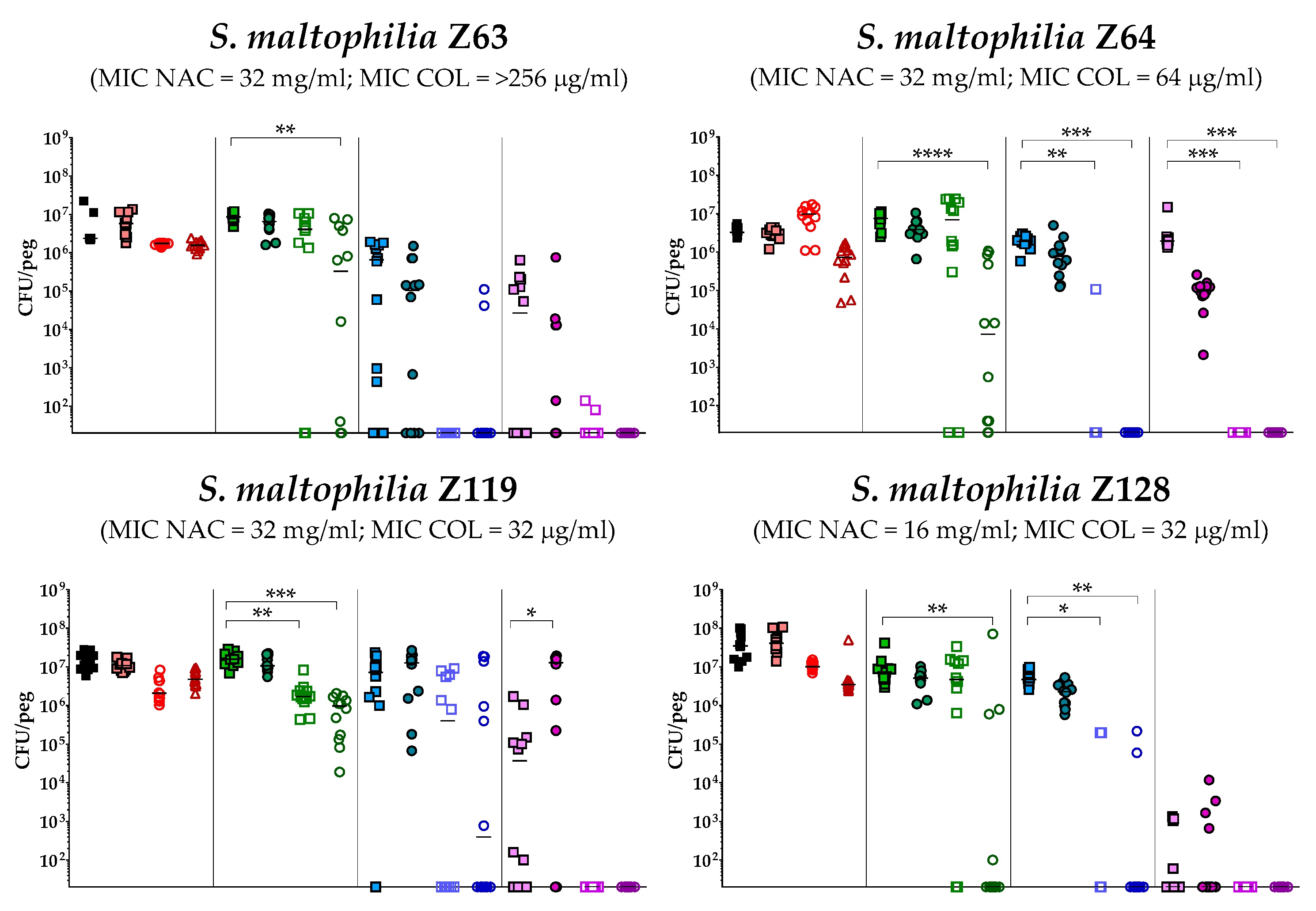
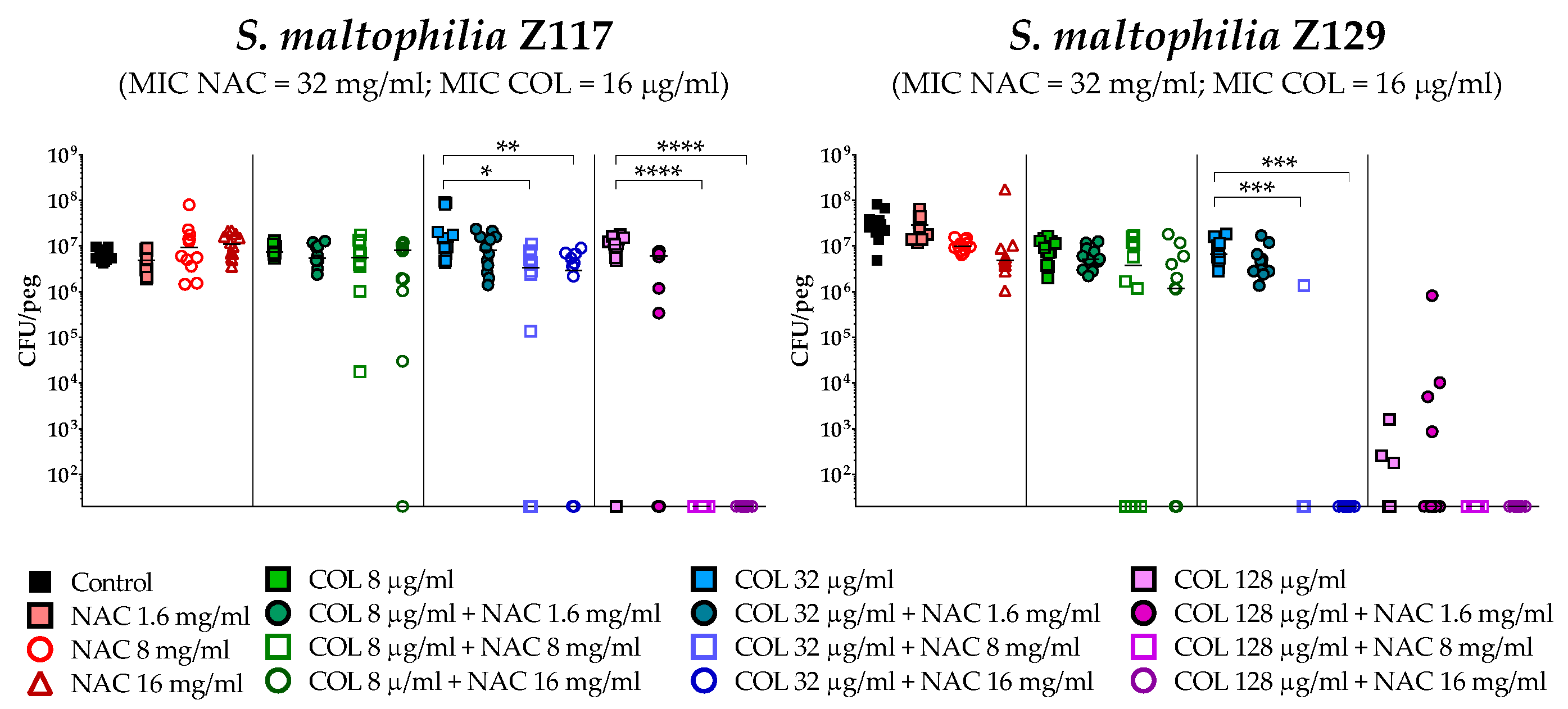
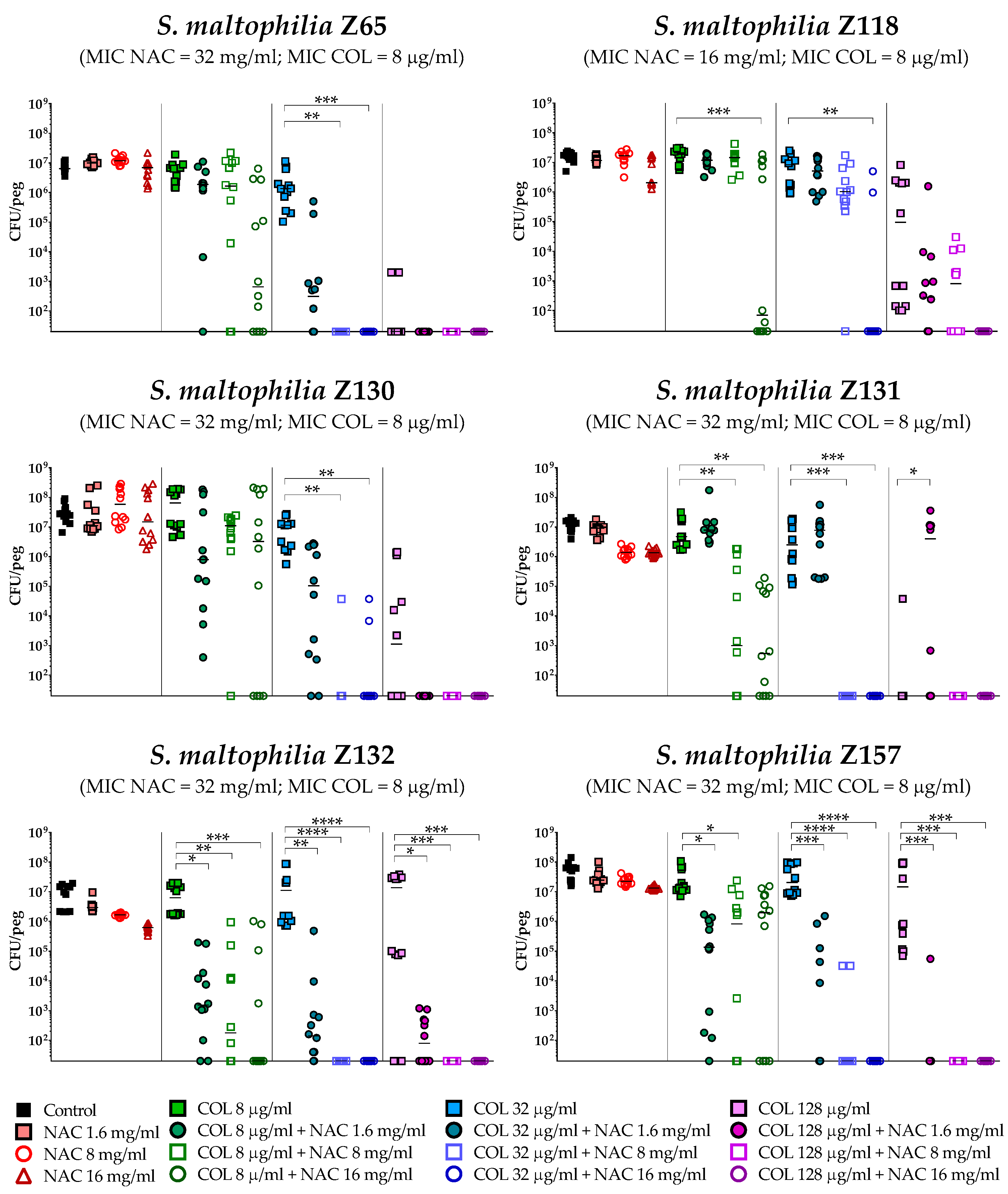
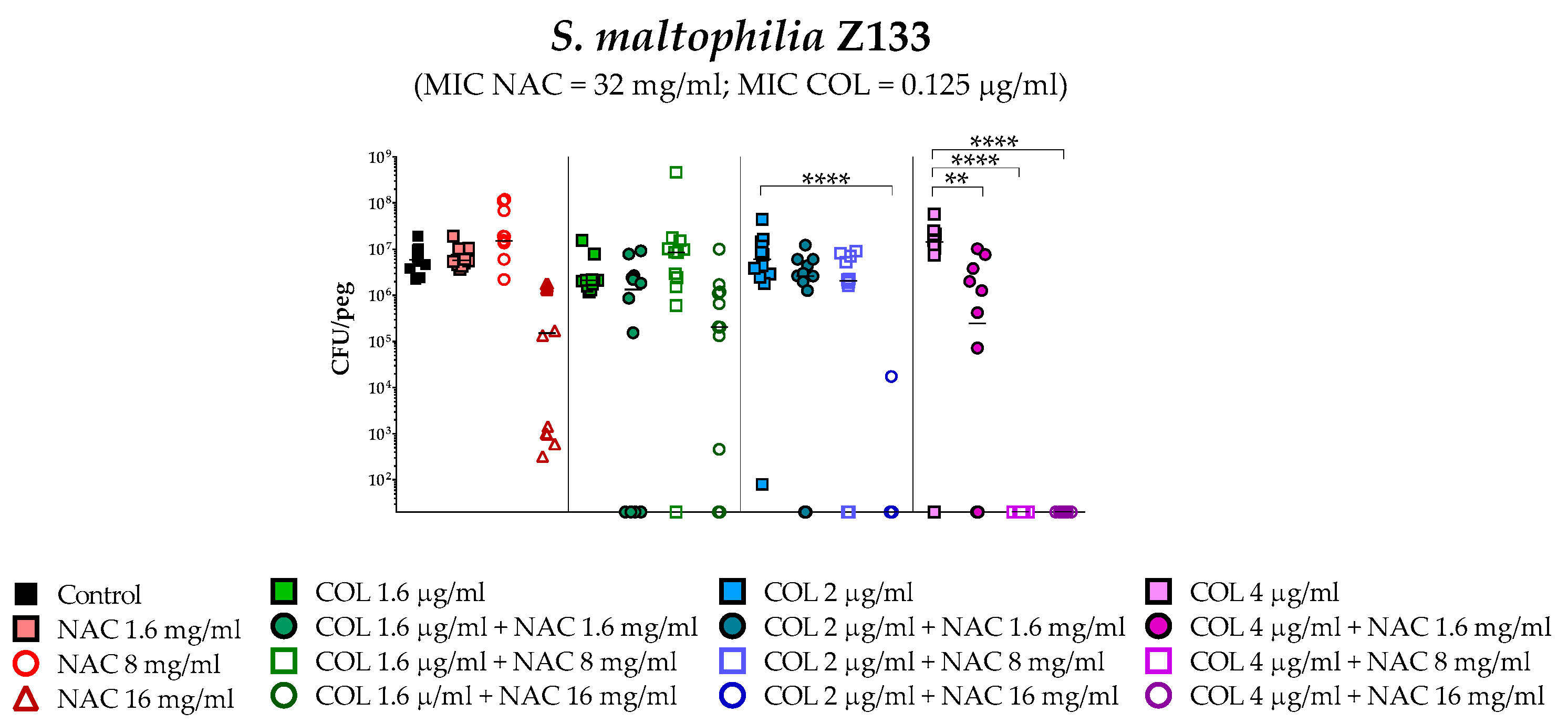
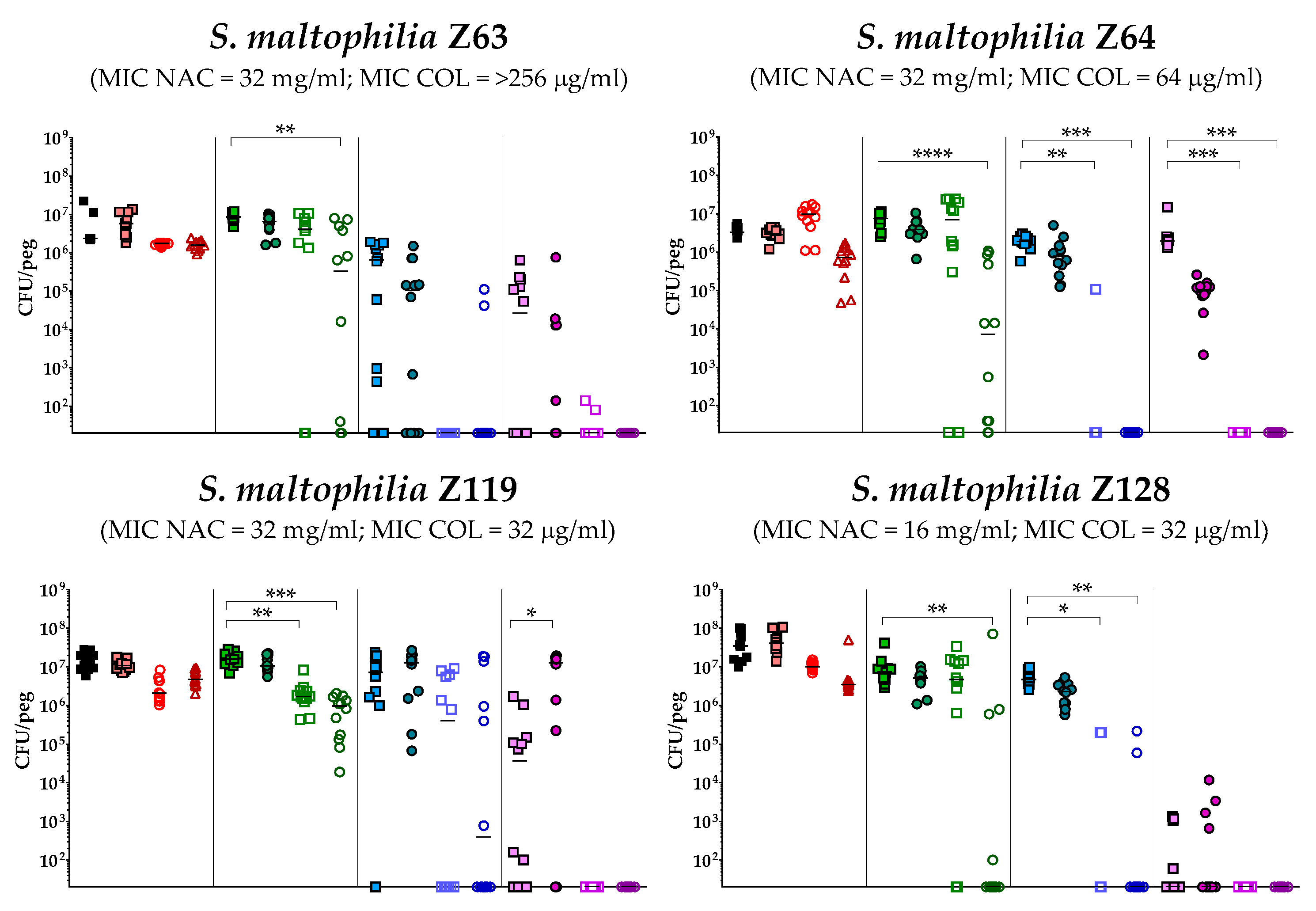
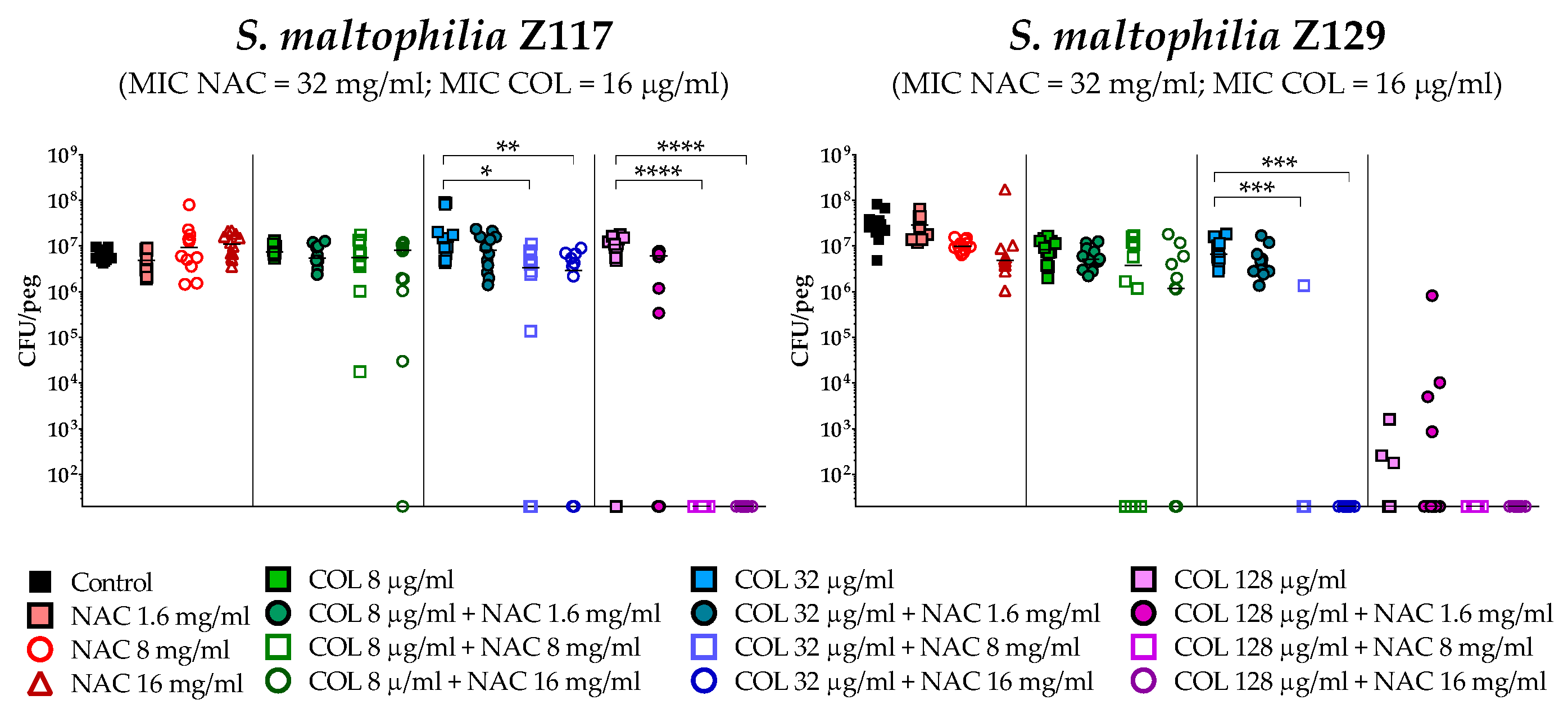
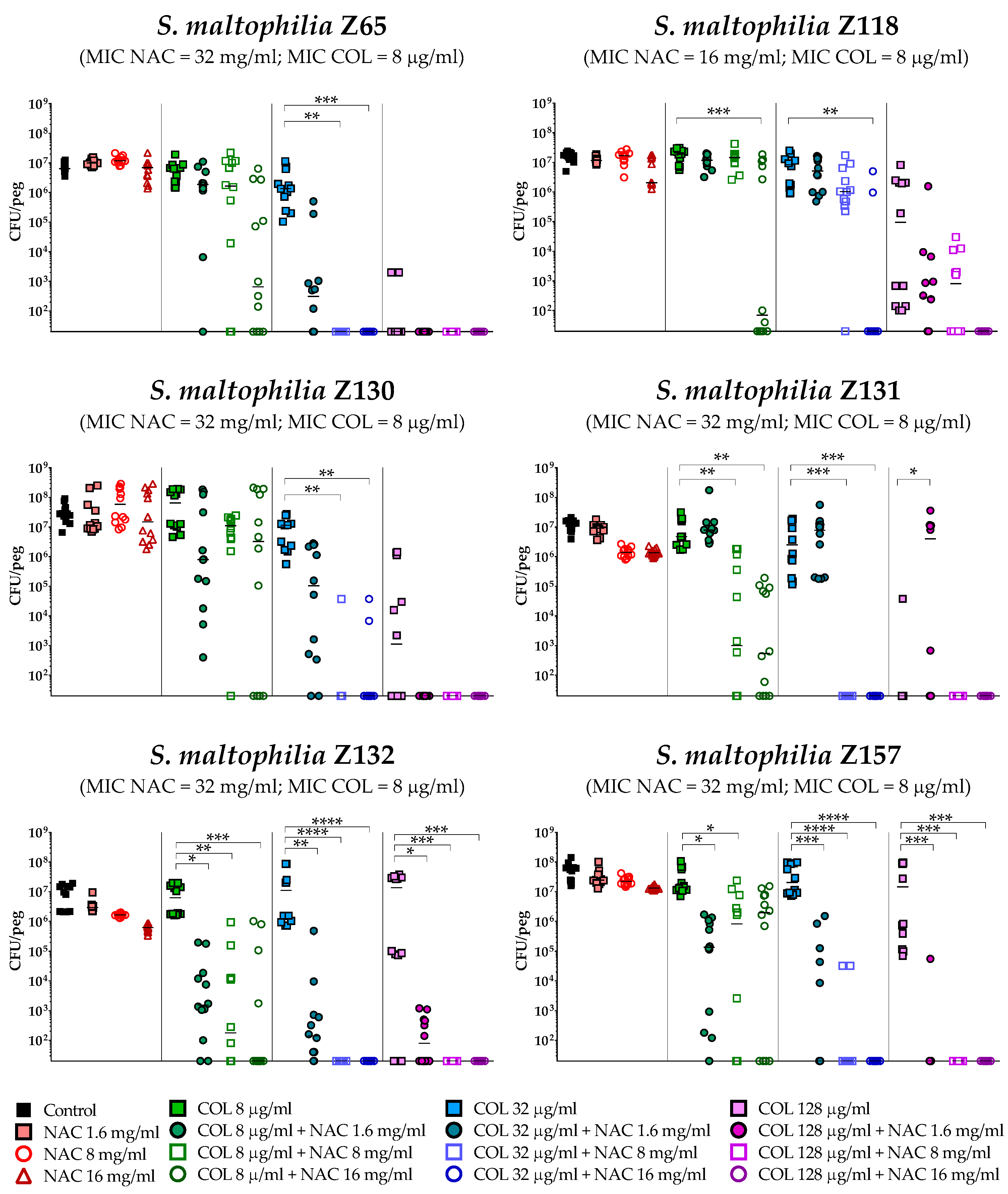
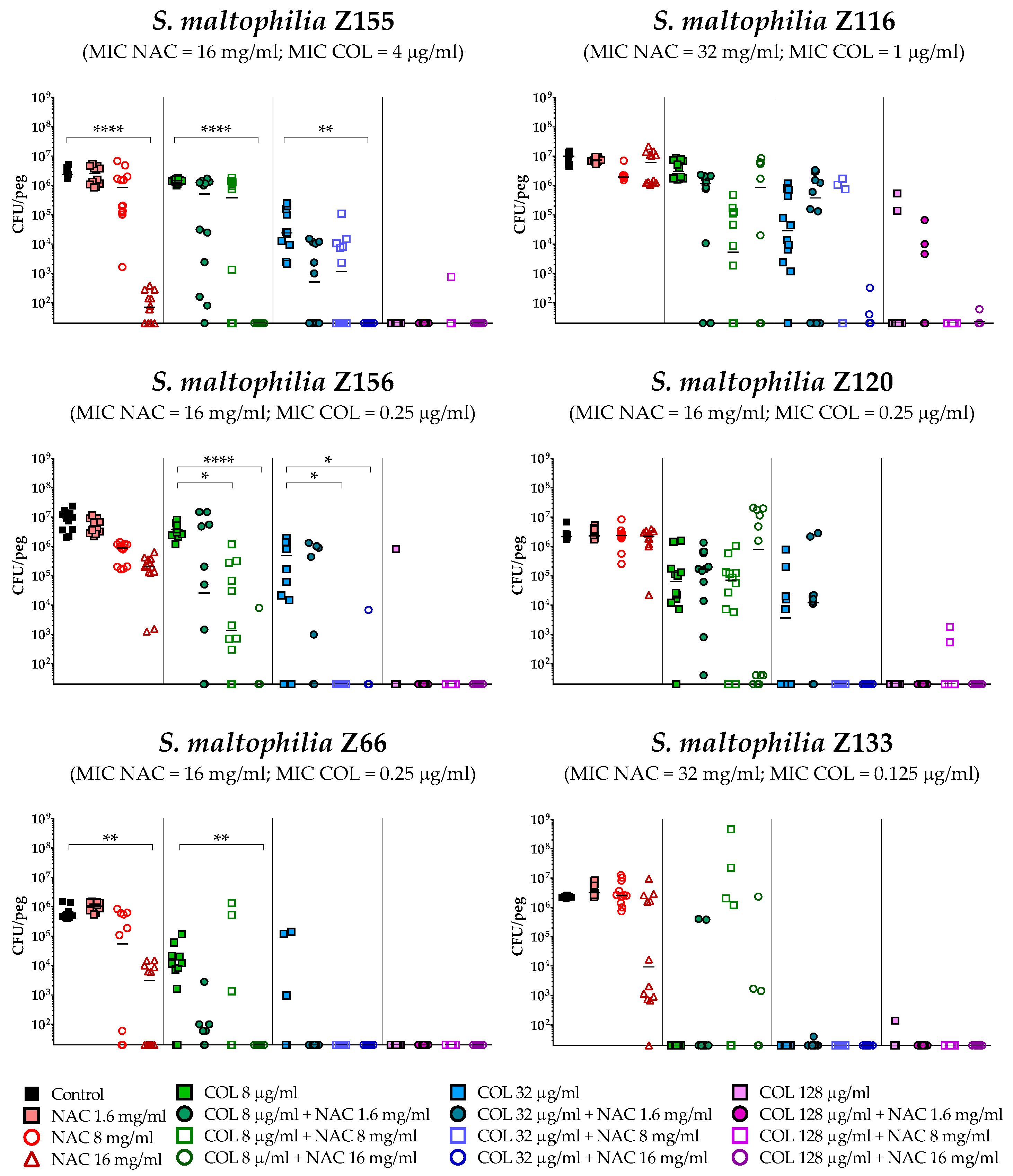
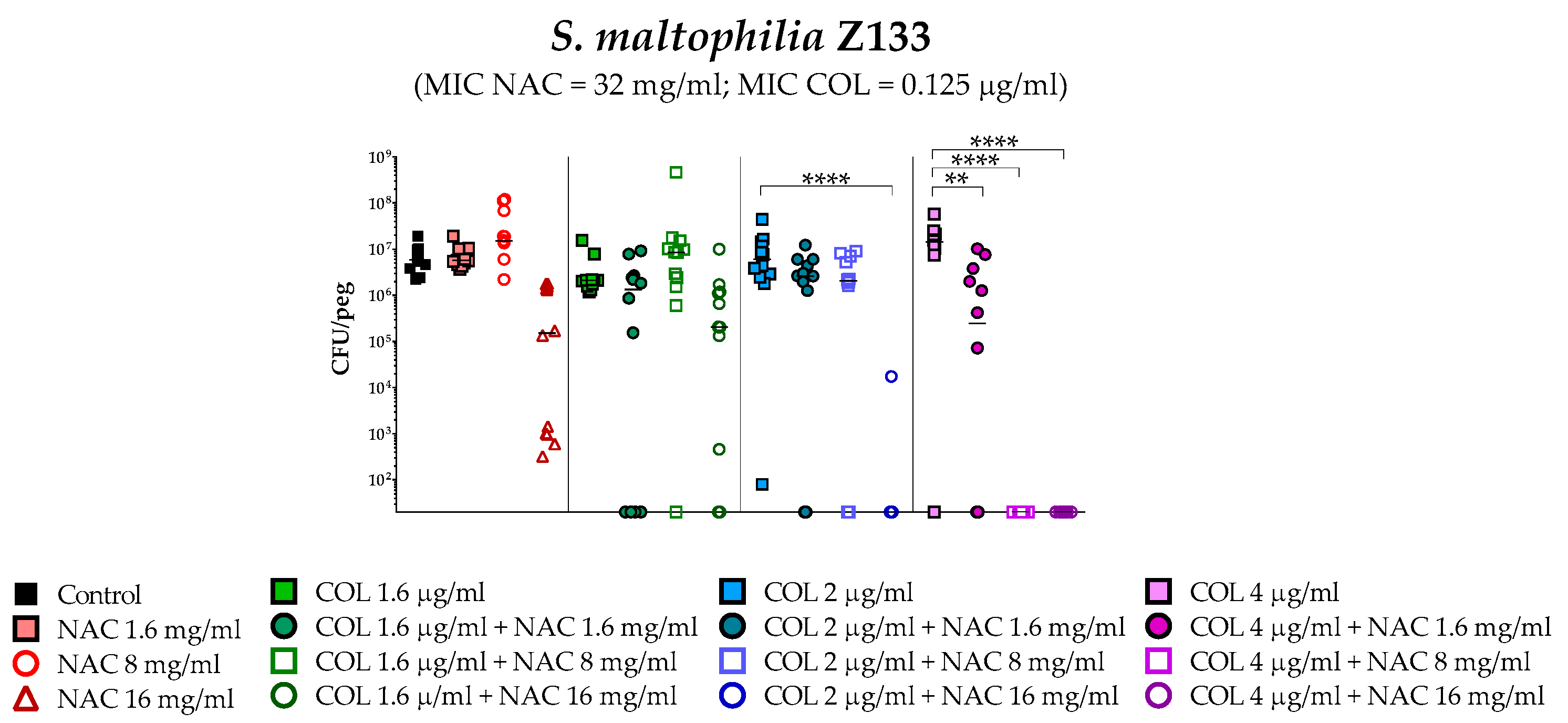
2.4. In Vitro Activity of Colistin/NAC Combinations against S. maltophilia Biofilms
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains Tested
4.2. Preparation of NAC-Containing Medium
4.3. Checkerboard Assays
4.4. Time–Kill Assays of Colistin/NAC Combinations Against Planktonic Cultures
4.5. In Vitro Biofilm Susceptibility Testing
4.6. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Conflicts of Interest
References
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed]
- Giordani, B.; Amato, A.; Majo, F.; Ferrari, G.; Quattrucci, S.; Minicucci, L.; Padoan, R.; Floridia, G.; Puppo Fornaro, G.; Taruscio, D.; et al. Italian Cystic Fibrosis Registry. Report 2011–2014. Epidemiol. Prev. 2018, 42, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Amin, R.; Waters, V. Antibiotic treatment for Stenotrophomonas maltophilia in people with cystic fibrosis. Cochrane. Database. Syst. Rev. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, B.P.; Freedman, S.D. Cystic fibrosis. Lancet 2009, 373, 1891–1904. [Google Scholar] [CrossRef]
- Esposito, A.; Pompilio, A.; Bettua, C.; Crocetta, V.; Giacobazzi, E.; Fiscarelli, E.; Jousson, O.; Di Bonaventura, G. Evolution of Stenotrophomonas maltophilia in cystic fibrosis lung over chronic infection: A genomic and phenotypic population study. Front. Microbiol. 2017, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Barsky, E.E.; Williams, K.A.; Priebe, G.P.; Sawicki, G.S. Incident Stenotrophomonas maltophilia infection and lung function decline in cystic fibrosis. Pediatr. Pulmonol. 2017, 52, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Waters, V.; Atenafu, E.G.; Lu, A.; Yau, Y.; Tullis, E.; Ratjen, F. Chronic Stenotrophomonas maltophilia infection and mortality or lung transplantation in cystic fibrosis patients. J. Cyst. Fibros. 2013, 12, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Díez-Aguilar, M.; Ekkelenkamp, M.; Morosini, M.I.; Merino, I.; de Dios Caballero, J.; Jones, M.; van Westreenen, M.; Tunney, M.M.; Cantón, R.; Fluit, A.C. Antimicrobial susceptibility of non-fermenting Gram-negative pathogens isolated from cystic fibrosis patients. Int. J. Antimicrob. Agents. 2019, 53, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, A.A.; Stenström, T.A.; Okoh, A.I. Stenotrophomonas maltophilia as an emerging ubiquitous pathogen: Looking beyond contemporary antibiotic therapy. Front. Microbiol. 2017, 8, 2276. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Tolker-Nielsen, T.; Jensen, P.Ø.; Wang, H.; Høiby, N. Antimicrobial resistance, respiratory tract infections and role of biofilms in lung infections in cystic fibrosis patients. Adv. Drug. Deliv. Rev. 2015, 85, 7–23. [Google Scholar] [CrossRef]
- Chang, Y.T.; Lin, C.Y.; Chen, Y.H.; Hsueh, P.R. Update on infections caused by Stenotrophomonas maltophilia with particular attention to resistance mechanisms and therapeutic options. Front. Microbiol. 2015, 6, 893. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters. Version 9.0. 2019. Available online: http://www.eucast.org (accessed on 18 February 2019).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Maselli, D.J.; Keyt, H.; Restrepo, M.I. Inhaled antibiotic therapy in chronic respiratory diseases. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Fiel, S.B. Aerosolized antibiotics in cystic fibrosis: An update. Expert. Rev. Respir. Med. 2014, 8, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Blasi, F.; Page, C.; Rossolini, G.M.; Pallecchi, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. The effect of N-acetylcysteine on biofilms: Implications for the treatment of respiratory tract infections. Respir. Med. 2016, 117, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Pollini, S.; Di Pilato, V.; Landini, G.; Di Maggio, T.; Cannatelli, A.; Sottotetti, S.; Cariani, L.; Aliberti, S.; Blasi, F.; Sergio, F.; et al. In vitro activity of N-acetylcysteine against Stenotrophomonas maltophilia and Burkholderia cepacia complex grown in planktonic phase and biofilm. PloS. One. 2018, 13, e0203941. [Google Scholar] [CrossRef] [PubMed]
- Vardakas, K.Z.; Voulgaris, G.L.; Samonis, G.; Falagas, M.E. Inhaled colistin monotherapy for respiratory tract infections in adults without cystic fibrosis: A systematic review andmeta-analysis. Int. J. Antimicrob. Agents. 2018, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Boisson, M.; Grégoire, N.; Cormier, M.; Gobin, P.; Marchand, S.; Couet, W.; Mimoz, O. Pharmacokinetics of nebulized colistin methanesulfonate in critically ill patients. J. Antimicrob. Chemother. 2017, 72, 2607–2612. [Google Scholar] [CrossRef]
- Boisson, M.; Jacobs, M.; Grégoire, N.; Gobin, P.; Marchand, S.; Couet, W.; Mimoz, O. Comparison of intrapulmonary and systemic pharmacokinetics of colistin methanesulfonate (CMS) and colistin after aerosol delivery and intravenous administration of CMS in critically ill patients. Antimicrob. Agents. Chemother. 2014, 58, 7331–7339. [Google Scholar] [CrossRef]
- Abbott, I.J.; Peleg, A.Y. Stenotrophomonas, Achromobacter, and nonmelioid Burkholderia species: Antimicrobial resistance and therapeutic strategies. Semin. Respir. Crit. Care. Med. 2015, 36, 99–110. [Google Scholar] [CrossRef]
- Pollini, S.; Boncompagni, S.; Di Maggio, T.; Di Pilato, V.; Spanu, T.; Fiori, B.; Blasi, F.; Aliberti, S.; Sergio, F.; Rossolini, G.M.; et al. In vitro synergism of colistin in combination with N-acetylcysteine against Acinetobacter baumannii grown in planktonic phase and in biofilms. J. Antimicrob. Chemother. 2018, 73, 2388–2395. [Google Scholar] [CrossRef]
- Martínez-Servat, S.; Yero, D.; Huedo, P.; Marquez, R.; Molina, G.; Daura, X.; Gibert, I. Heterogeneous colistin-resistance phenotypes coexisting in Stenotrophomonas maltophilia isolates influence colistin susceptibility testing. Front. Microbiol. 2018, 9, 2871. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Stremick, C.A.; Turner, R.J.; Allan, N.D.; Olson, M.E.; Ceri, H. Microtiter susceptibility testing of microbes growing on peg lids: A miniaturized biofilm model for high-throughput screening. Nat. Protoc. 2010, 5, 1236–1254. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar] [CrossRef] [PubMed]
- Kukut Hatipoglu, M.; Hickey, A.J.; Garcia–Contreras, L. Pharmacokinetics and pharmacodynamics of high doses of inhaled dry powder drugs. Int. J. Pharm. 2018, 549, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Murgia, X.; Siebenbürger, L.; Börger, C.; Schwarzkopf, K.; Sewald, K.; Häussler, S.; Braun, A.; Lehr, C.M.; Hittinger, M.; et al. Human airway mucus alters susceptibility of Pseudomonas aeruginosa biofilms to tobramycin, but not colistin. J. Antimicrob. Chemother. 2018, 73, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Conole, D.; Keating, G.M. Colistimethate sodium dry powder for inhalation: A review of its use in the treatment of chronic Pseudomonas aeruginosa infection in patients with cystic fibrosis. Drugs 2014, 74, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Mata, A.F.; Sarnaik, A.A. Bronchoscopy with N–acetylcysteine lavage in severe respiratory failure from pertussis infection. Pediatrics 2013, 132, e1418–e1423. [Google Scholar] [CrossRef] [PubMed]
- Akdag Cayli, Y.; Sahin, S.; Buttini, F.; Balducci, A.G.; Montanari, S.; Vural, I.; Oner, L. Dry powders for the inhalation of ciprofloxacin or levofloxacin combined with a mucolytic agent for cystic fibrosis patients. Drug. Dev. Ind. Pharm. 2017, 43, 1378–1389. [Google Scholar] [CrossRef]
- Manniello, M.D.; Del Gaudio, P.; Aquino, R.P.; Russo, P. Clarithromycin and N-acetylcysteine co-spray-dried powders for pulmonary drug delivery: A focus on drug solubility. Int. J. Pharm. 2017, 533, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Zhao, J.; Maifiah, M.H.M.; Velkov, T.; Schreiber, F.; Li, J. Metabolic Responses to Polymyxin Treatment in Acinetobacter baumannii ATCC 19606: Integrating Transcriptomics and Metabolomics with Genome-Scale Metabolic Modeling. mSystems 2019, 4, e00157-18. [Google Scholar] [CrossRef] [PubMed]
- Deris, Z.Z.; Akter, J.; Sivanesan, S.; Roberts, K.D.; Thompson, P.E.; Nation, R.L.; Li, J.; Velkov, T. A secondary mode of action of polymyxins against Gram-negative bacteria involves the inhibition of NADH-quinone oxidoreductase activity. J. Antibiot. 2014, 67, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents. Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhu, Y.; Fu, J.; Qiu, J.; Yin, J. Enhanced NADH Metabolism Involves Colistin-Induced Killing of Bacillus subtilis and Paenibacillus polymyxa. Molecules 2019, 24, 387. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Bonapace, C.R.; Bosso, J.A.; Friedrich, L.V.; White, R.L. Comparison of methods of interpretation of checkerboard synergy testing. Diagn. Microbiol. Infect. Dis. 2002, 44, 363–366. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guideline, 1st ed.; M026-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 1999. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MLST | Origin | Antibiotics | ||||
|---|---|---|---|---|---|---|---|
| MIC (μg/mL) | |||||||
| COL | SXT | CAZ | LVX | MIN | |||
| Z63 | NA | BSI | >256 | 0.5 | 2 | ≤0.25 | 0.125 |
| Z64 | NA | BSI | 64 | 2 | 64 | 2 | 2 |
| Z119 | NA | LRTI | 32 | 0.5 | 32 | 2 | 0.5 |
| Z128 | NA | LRTI | 32 | ≤0.25 | 4 | 1 | 0.25 |
| Z117 | NA | LRTI | 16 | 0.5 | 64 | 0.5 | 0.25 |
| Z129 | NA | LRTI | 16 | ≤0.25 | 4 | 1 | 0.25 |
| Z65 | NA | IAI | 8 | 1 | 64 | 2 | 1 |
| Z118 | ST162 | LRTI | 8 | 0.5 | 8 | 2 | 0.25 |
| Z130 | NA | IAI | 8 | 0.5 | 16 | 16 | 2 |
| Z131 | NA | BSI | 8 | >8 | 64 | 32 | 1 |
| Z132 | NA | LRTI | 8 | 1 | 2 | 16 | 1 |
| Z157 | NA | CF | 8 | 0.5 | 4 | 2 | 1 |
| Z155 | ST335 | CF | 4 | >8 | 32 | 4 | 2 |
| Z116 | NA | LRTI | 1 | 0.5 | 16 | 2 | 0.25 |
| Z156 | NA | CF | 0.25 | 1 | 16 | 2 | 0.25 |
| Z120 | ST334 | LRTI | 0.25 | 0.5 | 32 | 1 | 0.5 |
| Z66 | NA | LRTI | 0.25 | 0.5 | ≤1 | 1 | 0.25 |
| Z133 | NA | LRTI | 0.125 | 1 | 2 | 1 | 0.25 |
| Strain | MIC | NAC Concentrations (mg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NAC (mg/mL) | COL (µg/mL) | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | ||||||||
| COL (µg/mL) | FICI | COL (µg/mL) | FICI | COL (µg/mL) | FICI | COL (µg/mL) | FICI | COL (µg/mL) | FICI | COL (µg/mL) | FICI | COL (µg/mL) | FICI | |||
| Z63 | 32 | >256 | 32 | 0.14 | 16 | 0.09 | 2 | 0.07 | 1 | 0.13 | 0.5 | 0.25 | ≤0.25 | 0.50 | ≤0.25 | 1.00 |
| Z64 | 32 | 64 | 64 | 0.02 | 32 | 0.53 | 16 | 0.31 | 1 | 0.14 | 1 | 0.27 | 0.5 | 0.51 | 0.25 | 1.00 |
| Z119 | 32 | 32 | 32 | 1.02 | 16 | 0.53 | 8 | 0.31 | 4 | 0.25 | 1 | 0.28 | 1 | 0.53 | ≤0.25 | 1.01 |
| Z128 | 16 | 32 | 32 | 1.03 | 32 | 1.06 | 16 | 0.63 | 2 | 0.31 | 1 | 0.53 | ≤0.25 | 1.01 | ≤0.25 | 2.01 |
| Z117 | 32 | 16 | 32 | 2.02 | 32 | 2.03 | 16 | 1.06 | 1 | 0.19 | 1 | 0.31 | ≤0.25 | 0.52 | ≤0.25 | 1.02 |
| Z129 | 32 | 16 | 32 | 2.02 | 16 | 1.03 | 16 | 1.06 | 2 | 0.25 | 1 | 0.31 | 0.5 | 0.53 | ≤0.25 | 1.02 |
| Z65 | 32 | 8 | 16 | 2.02 | 8 | 1.03 | 4 | 0.56 | 1 | 0.25 | ≤0.25 | 0.28 | ≤0.25 | 0.53 | ≤0.25 | 1.03 |
| Z118 | 16 | 8 | 2 | 0.28 | 2 | 0.31 | 2 | 0.38 | 1 | 0.38 | ≤0.25 | 0.53 | ≤0.25 | 1.03 | ≤0.25 | 2.03 |
| Z130 | 32 | 8 | 16 | 2.02 | 8 | 1.03 | 2 | 0.31 | 1 | 0.25 | 1 | 0.38 | 0.5 | 0.56 | ≤0.25 | 1.03 |
| Z131 | 32 | 8 | 16 | 2.02 | 8 | 1.03 | 4 | 0.56 | 2 | 0.38 | 1 | 0.38 | 0.5 | 0.56 | ≤0.25 | 1.03 |
| Z132 | 32 | 8 | 16 | 2.02 | 8 | 1.03 | 2 | 0.31 | 1 | 0.25 | 1 | 0.38 | 0.5 | 0.56 | ≤0.25 | 1.03 |
| Z157 | 32 | 8 | 32 | 4.02 | 32 | 4.03 | 8 | 1.06 | 2 | 0.38 | 1 | 0.38 | 1 | 0.63 | ≤0.25 | 1.03 |
| Z155 | 16 | 4 | 4 | 1.03 | 4 | 1.06 | 2 | 0.63 | 1 | 0.50 | 1 | 0.75 | ≤0.25 | 1.06 | ≤0.25 | 2.06 |
| Z116 | 32 | 1 | 1 | 1.02 | 1 | 1.03 | 1 | 1.06 | 1 | 1.13 | 0.5 | 0.75 | 0.25 | 0.75 | ≤0.003 | 1.00 |
| Z156 | 16 | 0.25 | 0.5 | 2.03 | 0.5 | 2.06 | 0.25 | 1.13 | 0.25 | 1.25 | 0.06 | 0.75 | ≤0.003 | 1.02 | ≤0.003 | 2.02 |
| Z120 | 16 | 0.25 | 0.5 | 2.03 | 0.5 | 2.06 | 0.25 | 1.13 | 0.25 | 1.25 | 0.06 | 0.75 | ≤0.003 | 1.02 | ≤0.003 | 2.02 |
| Z66 | 16 | 0.25 | 0.25 | 1.03 | 0.25 | 1.06 | 0.25 | 1.13 | 0.125 | 0.75 | 0.125 | 1.00 | ≤0.003 | 1.02 | ≤0.003 | 2.02 |
| Z133 | 32 | 0.125 | 0.25 | 2.03 | 0.25 | 2.06 | 0.125 | 1.13 | 0.125 | 1.25 | 0.125 | 1.50 | ≤0.003 | 1.03 | ≤0.003 | 2.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciacci, N.; Boncompagni, S.; Valzano, F.; Cariani, L.; Aliberti, S.; Blasi, F.; Pollini, S.; Rossolini, G.M.; Pallecchi, L. In Vitro Synergism of Colistin and N-acetylcysteine against Stenotrophomonas maltophilia. Antibiotics 2019, 8, 101. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030101
Ciacci N, Boncompagni S, Valzano F, Cariani L, Aliberti S, Blasi F, Pollini S, Rossolini GM, Pallecchi L. In Vitro Synergism of Colistin and N-acetylcysteine against Stenotrophomonas maltophilia. Antibiotics. 2019; 8(3):101. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030101
Chicago/Turabian StyleCiacci, Nagaia, Selene Boncompagni, Felice Valzano, Lisa Cariani, Stefano Aliberti, Francesco Blasi, Simona Pollini, Gian Maria Rossolini, and Lucia Pallecchi. 2019. "In Vitro Synergism of Colistin and N-acetylcysteine against Stenotrophomonas maltophilia" Antibiotics 8, no. 3: 101. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030101