Effectiveness of Efflux Pump Inhibitors as Biofilm Disruptors and Resistance Breakers in Gram-Negative (ESKAPEE) Bacteria


Abstract
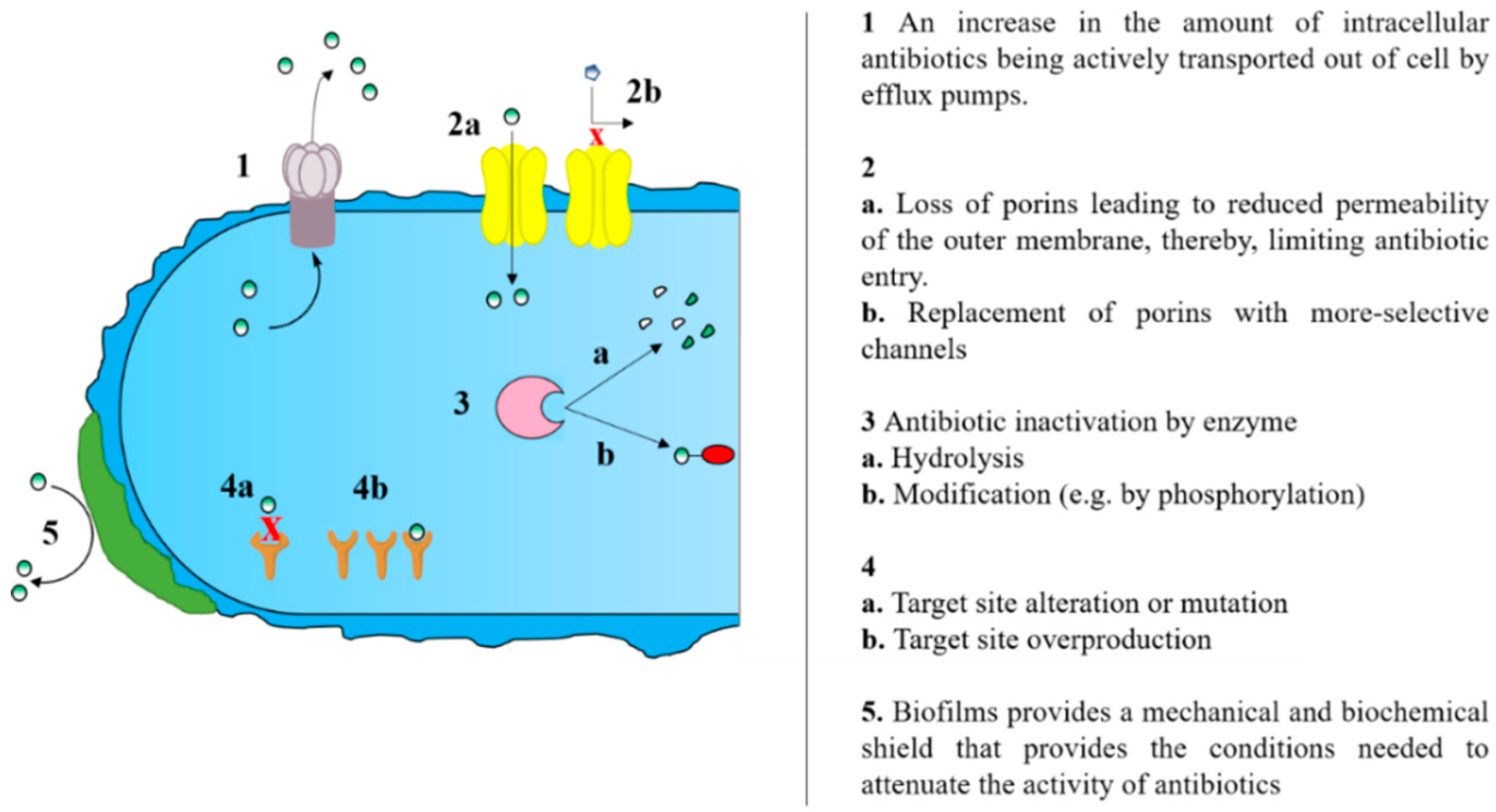
:1. Background
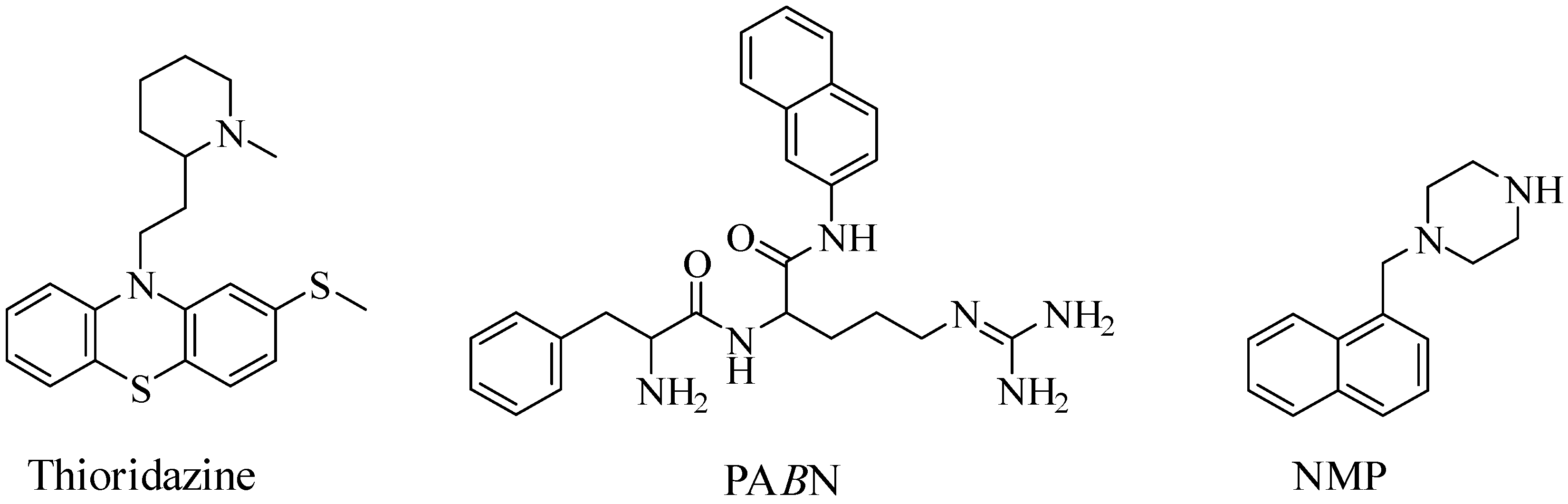
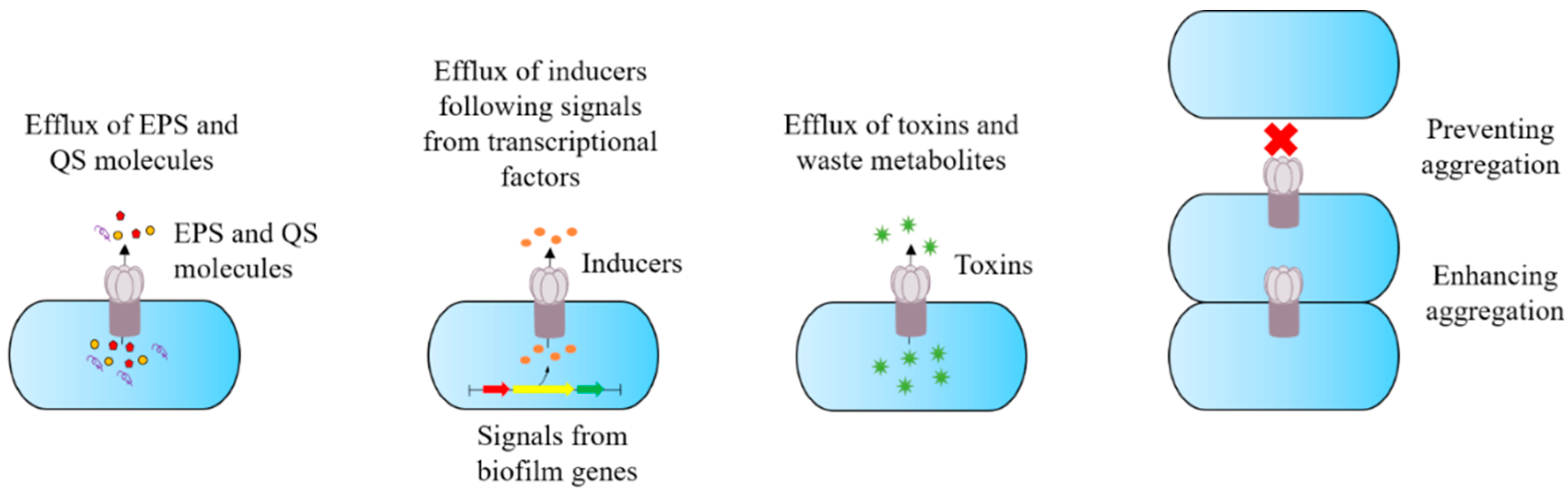
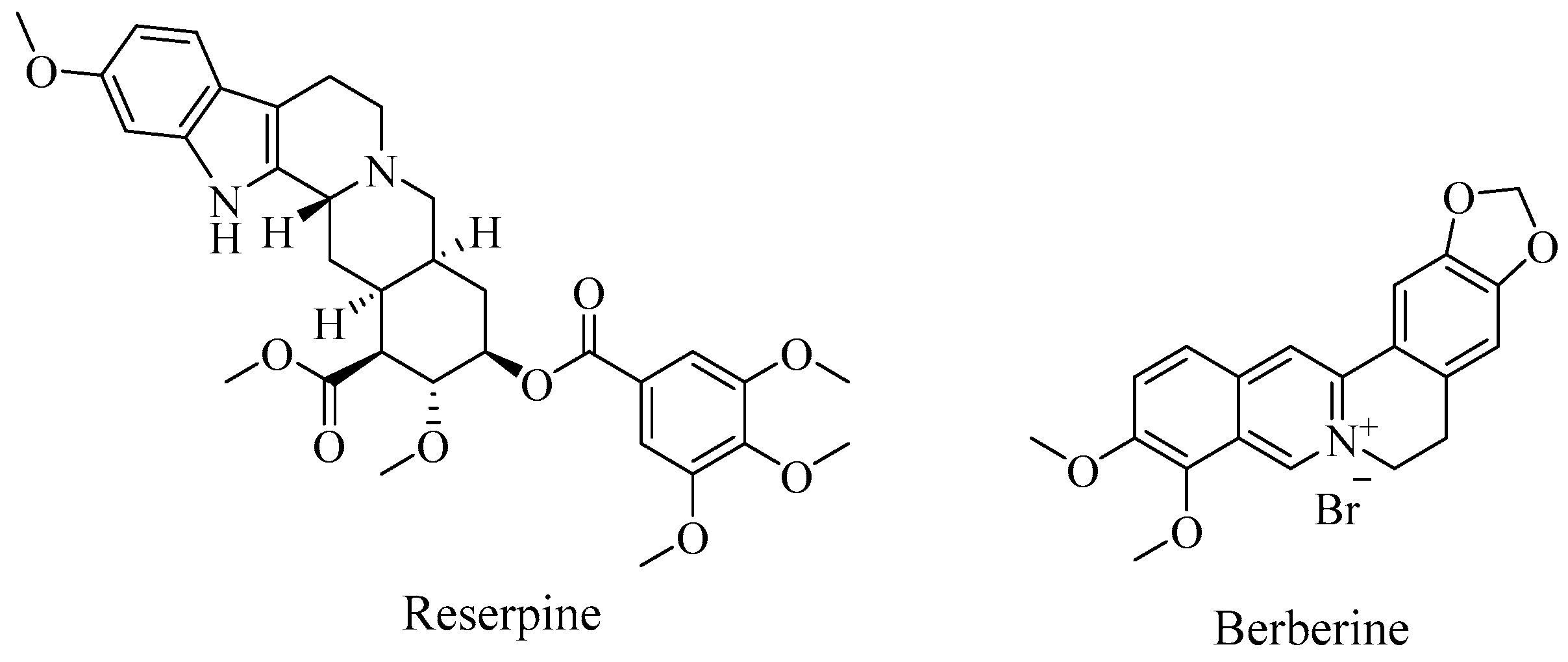
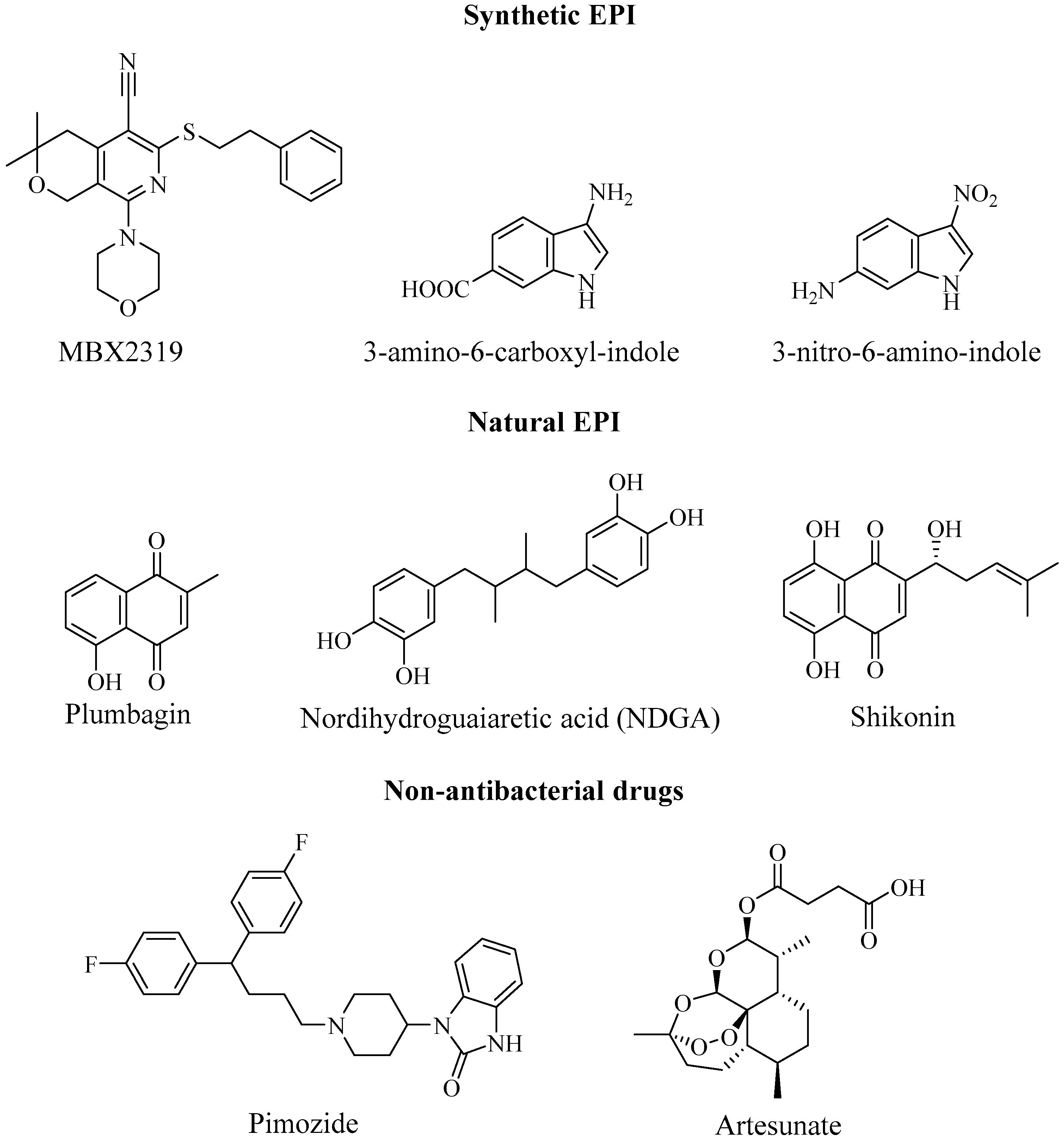
2. Efflux Pump Inhibitors and Antibiotic Potentiation
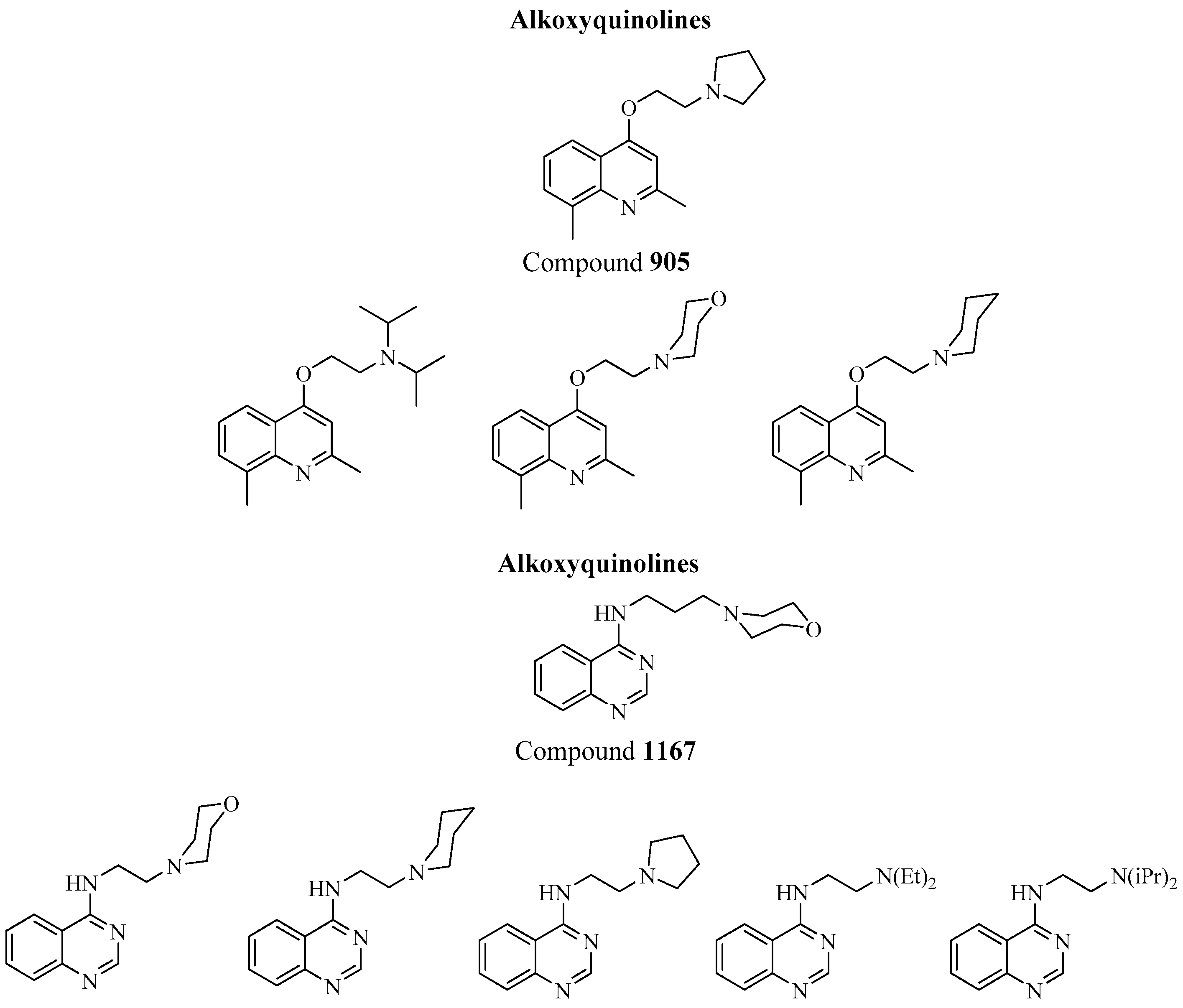
3. Klebsiella pneumoniae
EPIs and Their Potential Role in Biofilm Disruption in K. Pneumoniae
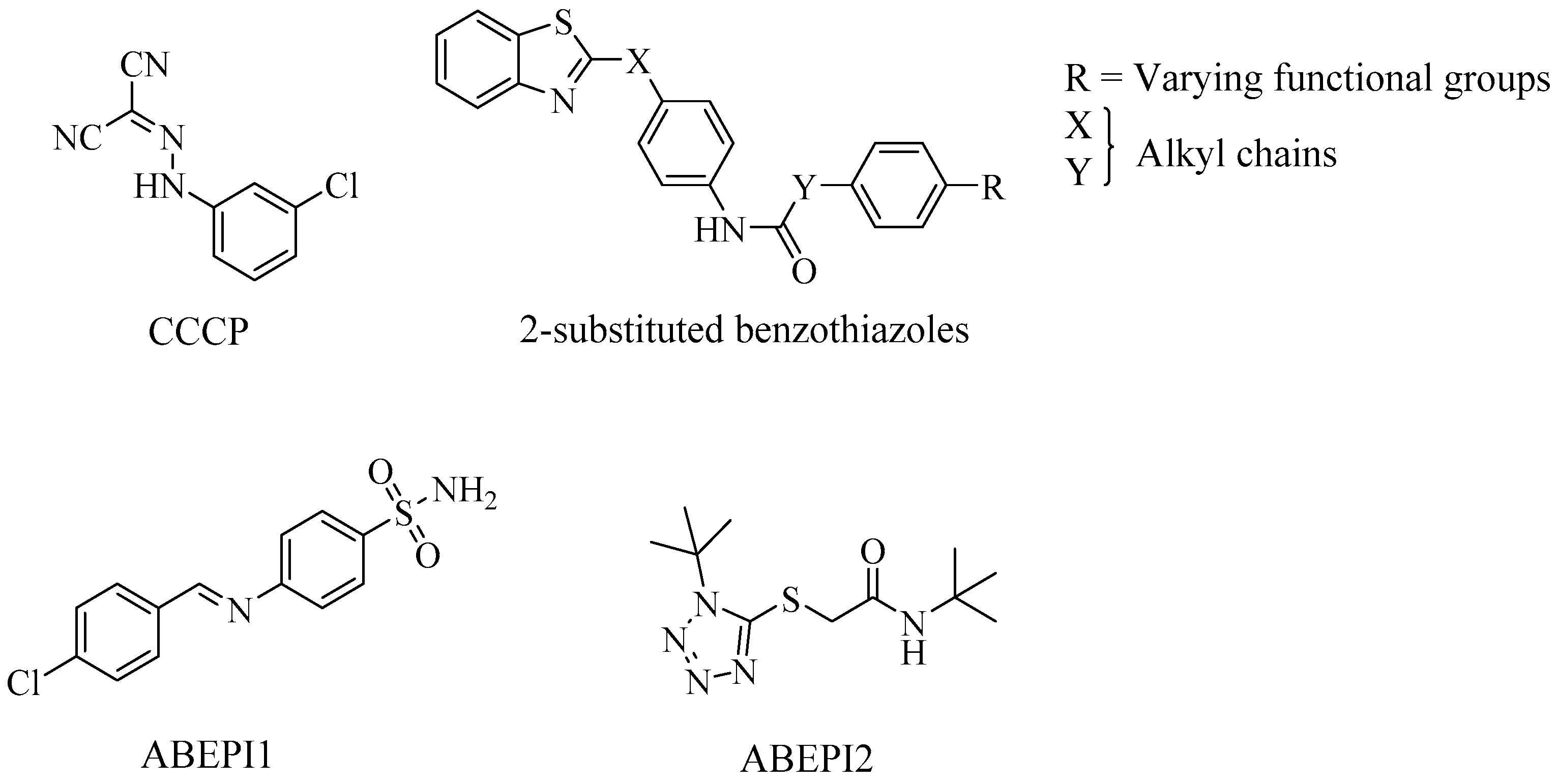
4. Acinetobacter Baumannii
EPIs and Their Potential Role in Biofilm Disruption in A. Baumannii
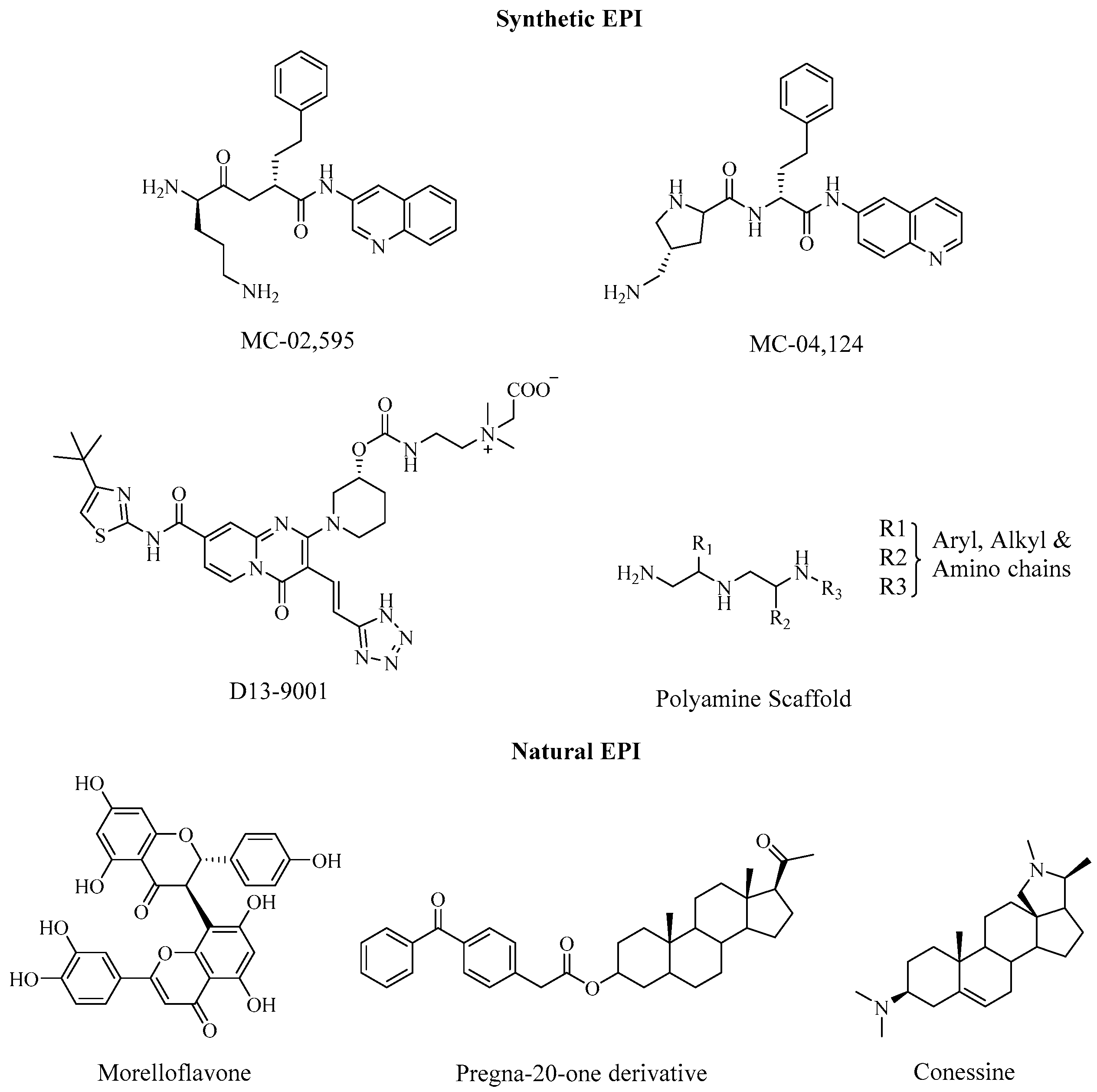
5. Pseudomonas Aeruginosa
5.1. EPIs and Their Potential Role in Biofilm Disruption in P. Aeruginosa
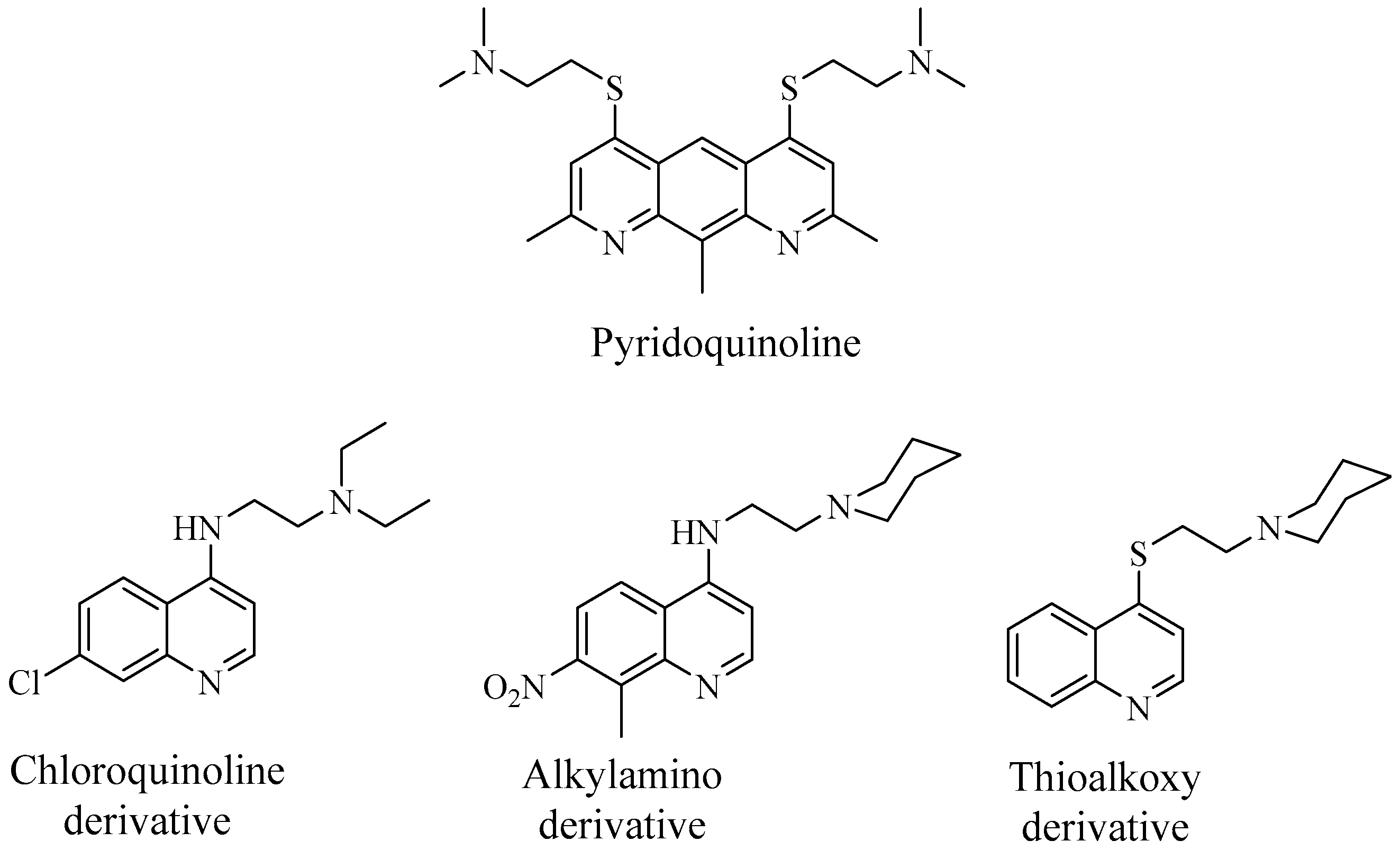
5.2. Enterobacter Spp.
5.3. EPIs and Their Potential Role in Biofilm Disruption in Enterobacter Spp.
6. Escherichia coli
EPIs and Their Potential Role in Biofilm Disruption in E. coli
7. Future Prospective and Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Piddock, L.J. Reflecting on the final report of the O’Neill Review on Antimicrobial Resistance. Lancet Infect. Dis. 2016, 16, 767–768. [Google Scholar] [CrossRef]
- WHO. High Levels of Antibiotic Resistance Found Worldwide, New Data Shows. Available online: http://www.who.int/news-room/detail/29-01-2018-high-levels-of-antibiotic-resistance-found-worldwide-new-data-shows (accessed on 27 August 2018).
- Poole, K. Mechanisms of bacterial biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 55S–64S. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Mech. Antibiot. Resist. 2016, 4, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Soto, S.M. Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm. Virulence 2015, 5594, 223–229. [Google Scholar]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. Apmis 2013, 121, 1–51. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.Z.; Høiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Sayen, S.; Al-Dhabi, N. Biofilm Control and Antimicrobial Agents, 1st ed.; Taylor & Francis Group: Oakville, ON, Canada, 2014. [Google Scholar]
- Karatan, E.; Watnick, P. Signals, Regulatory Networks, and Materials that Build and Break Bacterial Biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310–347. [Google Scholar] [CrossRef]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2000, 44, 1818–1824. [Google Scholar] [CrossRef]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Pagès, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm. Pharmacol. Ther. 2008, 21, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Abee, T.; Kovács, Á.T.; Kuipers, O.P.; van der Veen, S. Biofilm formation and dispersal in Gram-positive bacteria. Curr. Opin. Biotechnol. 2011, 22, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Malik, A.; Ahmad, J.; Rizvi, M. A study of biofilm production by gram negative organisms isolated from diabetic foot ulcer patients. Biol. Med. 2011, 3, 147–157. [Google Scholar]
- Hoen, B.; Duval, X. Infective Endocarditis. N. Engl. J. Med. 2013, 368, 1425–1433. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A.; Murray, B.E. Enterococcal endocarditis: Can we win the war? Curr. Infect. Dis. Rep. 2012, 14, 339–349. [Google Scholar] [CrossRef]
- Fernández Guerrero, M.L.; González López, J.J.; Goyenechea, A.; Fraile, J.; de Górgolas, M. Endocarditis Caused by Staphylococcus aureus. Medicine 2009, 88, 1–22. [Google Scholar] [CrossRef]
- Chu, V.H.; Cabell, C.H.; Abrutyn, E.; Corey, G.R.; Hoen, B.; Miro, J.M.; Olaison, L.; Stryjewski, M.E.; Pappas, P.; Anstrom, K.J.; et al. Native valve endocarditis due to coagulase-negative staphylococci: Report of 99 episodes from the International Collaboration on Endocarditis Merged Database. Clin. Infect. Dis. 2004, 39, 1527–1530. [Google Scholar] [CrossRef]
- Silva Filho, L.V.R.F.; Ferreira, F.D.A.; Reis, F.J.C.; Britto, M.C.A.D.; Levy, C.E.; Clark, O.; Ribeiro, J.D. Pseudomonas aeruginosa infection in patients with cystic fibrosis: Scientific evidence regarding clinical impact, diagnosis, and treatment. J. Bras. Pneumol. 2013, 39, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.F. Editorial Commentary: The Many Faces of Pseudomonas aeruginosa in Chronic Obstructive Pulmonary Disease. Clin. Infect. Dis. 2008, 47, 1534–1536. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, N.P.; Alexander, M.; Burns, L.A.; Dellinger, E.P.; Garland, J.; Heard, S.O.; Lipsett, P.A.; Masur, H.; Mermel, L.A.; Pearson, M.L.; et al. Guidelines for the prevention of intravascular catheter-related infections. Clin. Infect. Dis. 2011, 52, e162–e193. [Google Scholar] [CrossRef] [PubMed]
- Gahlot, R.; Nigam, C.; Kumar, V.; Yadav, G.; Anupurba, S. Catheter-related bloodstream infections. Int. J. Crit. Illn. Inj. Sci. 2014, 4, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Saadatian-Elahi, M.; Teyssou, R.; Vanhems, P. Staphylococcus aureus, the major pathogen in orthopaedic and cardiac surgical site infections: A literature review. Int. J. Surg. 2008, 6, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Uçkay, I.; Harbarth, S.; Ferry, T.; Lübbeke, A.; Emonet, S.; Hoffmeyer, P.; Pittet, D. Meticillin resistance in orthopaedic coagulase-negative staphylococcal infections. J. Hosp. Infect. 2011, 79, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Nolazco, J.I.; González, M.I.; Favre, G.; Guegilo, G.; Tejerizo, J. A case of emphysematous cystitis caused by Klebsiella pneumoniae. Can. J. Urol. 2017, 24, 8932–8933. [Google Scholar]
- Pour, N.K.; Dusane, D.H.; Dhakephalkar, P.K.; Zamin, F.R.; Zinjarde, S.S.; Chopade, B.A. Biofilm formation by Acinetobacter baumanniistrains isolated from urinary tract infection and urinary catheters. FEMS Immunol. Med. Microbiol. 2011, 62, 328–338. [Google Scholar] [CrossRef]
- Kauffman, C.A.; Fisher, J.F.; Sobel, J.D.; Newman, C.A. Candida urinary tract infections-Diagnosis. Clin. Infect. Dis. 2011, 52, S452–S456. [Google Scholar] [CrossRef]
- Swaminathan, S.; Alangaden, G.J. Treatment of resistant enterococcal urinary tract infections. Curr. Infect. Dis. Rep. 2010, 12, 455–465. [Google Scholar] [CrossRef]
- Pallett, A.; Hand, K. Complicated urinary tract infections: Practical solutions for the treatment of multiresistant gram-negative bacteria. J. Antimicrob. Chemother. 2010, 65, 25–33. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.; Caroleo, B.; Amato, B.; de Gallelli, F.S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti. Infect. Ther. 2015, 13, 605–6013. [Google Scholar] [CrossRef]
- Kvist, M.; Hancock, V.; Klemm, P. Inactivation of efflux pumps abolishes bacterial biofilm formation. Appl. Environ. Microbiol. 2008, 74, 7376–7382. [Google Scholar] [CrossRef]
- Sabatini, S.; Piccioni, M.; Felicetti, T.; De Marco, S.; Manfroni, G.; Pagiotti, R.; Nocchetti, M.; Cecchetti, V.; Pietrella, D. Investigation on the effect of known potent: S. aureus NorA efflux pump inhibitors on the staphylococcal biofilm formation. RSC Adv. 2017, 7, 37007–37014. [Google Scholar] [CrossRef] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 12, 12–13. [Google Scholar] [CrossRef] [Green Version]
- Marquez, B. Bacterial efflux systems and efflux pumps inhibitors. Biochimie 2005, 87, 1137–1147. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Pascolini, C.; Donelli, G.; Balice, M.P.; Libori, M.F.; Tiracchia, V.; Salvia, A.; Varaldo, P.E. Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J. Appl. Microbiol. 2017, 123, 1003–1018. [Google Scholar] [CrossRef]
- De Rosa, F.G.; Corcione, S.; Cavallo, R.; Di Perri, G.; Bassetti, M. Critical issues for Klebsiella pneumoniae KPC-carbapenemase producing K. pneumoniae infections: A critical agenda. Future Microbiol. 2015, 10, 283–294. [Google Scholar] [CrossRef]
- WHO. WHO’s First Global Report on Antibiotic Resistance Reveals Serious, Worldwide Threat to Public Health. Available online: http://www.who.int/mediacentre/news/releases/2014/amr-report/en/ (accessed on 27 August 2018).
- Magesh, H.; Kumar, A.; Alam, A.; Sekar, U. Identification of natural compounds which inhibit biofilm formation in clinical isolates of Klebsiella pneumoniae. Indian, J. Exp. Biol. 2013, 51, 764–772. [Google Scholar]
- Zhou, X.Y.; Ye, X.G.; He, L.T.; Zhang, S.R.; Wang, R.L.; Zhou, J.; He, Z.S. In vitro characterization and inhibition of the interaction between ciprofloxacin and berberine against multidrug-resistant Klebsiella pneumoniae. J. Antibiot. 2016, 69, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Hasdemir, U.O.; Chevalier, J.; Nordmann, P.; Pagès, J.M. Detection and prevalence of active drug efflux mechanism in various multidrug-resistant Klebsiella pneumoniae strains from Turkey. J. Clin. Microbiol. 2004, 42, 2701–2706. [Google Scholar] [CrossRef] [Green Version]
- Mahamoud, A.; Chevalier, J.; Baitiche, M.; Adam, E.; Page, J.; Page, J. An alkylaminoquinazoline restores antibiotic activity in Gram-negative resistant isolates. Microbiology 2011, 157, 566–571. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, J.; Mahamoud, A.; Malle, M.; Barbe, J.; Page, J. Inhibitors of Antibiotic Efflux in Resistant Enterobacter aerogenes and Klebsiella pneumoniae Strains. Antimicrob. Agents Chemother. 2004, 48, 1043–1046. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, J.; Mahamoud, A.; Baitiche, M.; Adam, E.; Viveiros, M.; Smarandache, A.; Militaru, A.; Pascu, M.L.; Amaral, L.; Pagès, J. Quinazoline derivatives are efficient chemosensitizers of antibiotic activity in Enterobacter aerogenes, Klebsiella pneumoniae and Pseudomonas aeruginosa resistant strains. Int. J. Antimicrob. Agents 2010, 36, 164–168. [Google Scholar] [CrossRef] [Green Version]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef]
- Pannek, S.; Higgins, P.G.; Steinke, P.; Jonas, D.; Akova, M.; Bohnert, J.A.; Seifert, H.; Kern, W. Multidrug efflux inhibition in Acinetobacter baumannii: Comparison between 1-(1-naphthylmethyl)-piperazine and phenyl-arginine-β-naphthylamide. J. Antimicrob. Chemother. 2006, 57, 970–974. [Google Scholar] [CrossRef]
- Cortez-cordova, J.; Kumar, A. Activity of the efflux pump inhibitor phenylalanine-arginine B-naphthylamide against the AdeFGH pump of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2011, 37, 420–424. [Google Scholar] [CrossRef]
- Nikasa, P.; Abdi-Ali, A.; Rahmani-Badi, A.; Al-Hamad, A. In vitro Evaluation of Proton Motive Force-Dependent Efflux Pumps Among Multidrug Resistant Acinetobacter baumannii Isolated From Patients at Tehran Hospitals. Jundishapur J. Microbiol. 2013, 6, e6792. [Google Scholar] [CrossRef] [Green Version]
- Ardebili, A.; Talebi, M.; Azimi, L.; Lari, A. Effect of Efflux Pump Inhibitor Carbonyl Cyanide 3-Chlorophenylhydrazone on the Minimum Inhibitory Concentration of Ciprofloxacin in Acinetobacter baumannii Clinical Isolates. Jundishapur J. Microbiol. 2014, 7, e8691. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Zhu, W.; Tian, S.F.; Chu, Y.Z.; Chen, B.Y. Molecular characteristics and resistant mechanisms of imipenem-resistant Acinetobacter baumannii isolates in Shenyang, China. J. Microbiol. 2010, 48, 689–694. [Google Scholar] [CrossRef]
- Park, Y.K.; Ko, K.S. Effect of carbonyl cyanide 3-chlorophenylhydrazone (CCCP) on killing Acinetobacter baumannii by colistin. J. Microbiol. 2015, 53, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Runci, F.; Bonchi, C.; Frangipani, E.; Visaggio, D.; Visca, P. Acinetobacter baumannii biofilm formation in human serum and disruption by gallium. Antimicrob. Agents Chemother. 2017, 61, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchard, C.; Barnett, P.; Perlmutter, J.; Dunman, P.M. Identification of acinetobacter baumannii serum-associated antibiotic efflux pump inhibitors. Antimicrob. Agents Chemother. 2014, 58, 6360–6370. [Google Scholar] [CrossRef] [Green Version]
- Zloh, M.; Kaatz, G.W.; Gibbons, S. Inhibitors of multidrug resistance (MDR) have affinity for MDR substrates. Bioorganic Med. Chem. Lett. 2004, 14, 881–885. [Google Scholar] [CrossRef]
- Yilmaz, S.; Altinkanat-Gelmez, G.; Bolelli, K.; Guneser-Merdan, D.; Over-Hasdemir, M.; Yildiz, I.; Yalcin, I.; Bolelli, K. Pharmacophore generation of 2-substituted benzothiazoles as AdeABC efflux pump inhibitors in A. baumannii. SAR QSAR Environ. Res. 2014, 25, 551–563. [Google Scholar] [CrossRef]
- Mahmood, H.Y.; Jamshidi, S.; Sutton, J.M.; Rahman, K.M. Current Advances in Developing Inhibitors of Bacterial Multidrug Efflux Pumps. Curr. Med. Chem. 2016, 23, 1062–1081. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: Novel agents for combination therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Renau, T.; Leger, R.; Yen, R.; She, M.W.; Renau, T.E.; Le, R.; Flamme, E.M.; Sangalang, J.; Gannon, C.L.; Chamberland, S.; et al. Peptidomimetics of Efflux Pump Inhibitors Potentiate the Activity of Levofloxacin in Pseudomonas aeruginosa. Bioorganic Med. Chem. Lett. 2002, 12, 763–766. [Google Scholar] [CrossRef]
- Renau, T.E.; Filonova, L.; Flamme, E.M.; Wang, M.; Le, R.; Yen, R.; Madsen, D.; Griffith, D.; Chamberland, S.; Dudley, M.N.; et al. Conformationally-Restricted Analogues of Efflux Pump Inhibitors that Potentiate the Activity of Levofloxacin in Pseudomonas aeruginosa. Bioorganic Med. Chem. Lett. 2003, 13, 2755–2758. [Google Scholar] [CrossRef]
- Yoshida, K.; Nakayama, K.; Ohtsuka, M.; Kuru, N.; Yokomizo, Y.; Sakamoto, A.; Takemura, M.; Hoshino, K.; Kanda, H.; Nitanai, H.; et al. MexAB-OprM specific efflux pump inhibitors in Pseudomonas aeruginosa. Part 7: Highly soluble and in vivo active quaternary ammonium analogue D13-9001, a potential preclinical candidate. Bioorg. Med. Chem. 2007, 15, 7087–7097. [Google Scholar] [CrossRef]
- Hirakata, Y.; Kondo, A.; Hoshino, K.; Yano, H.; Arai, K. Efflux pump inhibitors reduce the invasiveness of Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2009, 34, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, L.; Molin, S. Synergistic activities of an efflux pump inhibitor and iron chelators against Pseudomonas aeruginosa growth and biofilm formation. Antimicrob. Agents Chemother. 2010, 54, 3960–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aron, Z.; Opperman, T.J. Optimization of a novel series of pyranopyridine RND efflux pump inhibitors. Curr. Opin. Microbiol. 2016, 33, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleeman, R.M.; Debevec, G.; Antonen, K.; Adams, J.L.; Santos, R.G.; Welmaker, G.S.; Houghten, R.A.; Giulianotti, M.A.; Shaw, L.N. Identification of a novel polyamine scaffold with potent efflux pump inhibition activity toward multi-drug resistant bacterial pathogens. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Mangiaterra, G.; Laudadio, E.; Cometti, M.; Mobbili, G.; Minnelli, C.; Massaccesi, L.; Citterio, B.; Biavasco, F.; Galeazzi, R. Inhibitors of multidrug efflux pumps of Pseudomonas aeruginosa from natural sources: An in silico high-throughput virtual screening and in vitro validation. Med. Chem. Res. 2017, 26, 414–430. [Google Scholar] [CrossRef]
- Aparna, V.; Dineshkumar, K.; Mohanalakshmi, N.; Velmurugan, D.; Hopper, W. Identification of natural compound inhibitors for multidrug efflux pumps of Escherichia coli and Pseudomonas aeruginosa using In Silico high-throughput virtual screening and In Vitro validation. PLoS ONE 2014, 9, e101840. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Singh, A.; Khan, A.U. Nanoparticles as Efflux Pump and Biofilm Inhibitor to Rejuvenate Bactericidal Effect of Conventional Antibiotics. Nanoscale Res. Lett. 2017, 12, 9–11. [Google Scholar] [CrossRef]
- Eugene Sanders, W.E.; Sanders, C.C. Enterobacter spp.: Pathogens poised to flourish at the turn of the century. Clin. Microbiol. Rev. 1997, 10, 220–241. [Google Scholar] [CrossRef]
- Tindall, B.J.; Sutton, G.; Garrity, G.M. Enterobacter aerogenes hormaeche and Edwards 1960 (Approved lists 1980) and Klebsiella mobilis bascomb et al. 1971 (approved lists 1980) share the same nomenclatural type (ATCC 13048) on the approved lists and are homotypic synonyms, with consequences for. Int. J. Syst. Evol. Microbiol. 2017, 67, 502–504. [Google Scholar]
- Guérin, F.; Lallement, C.; Isnard, C.; Dhalluin, A.; Cattoir, V.; Giard, J.C. Landscape of resistance-nodulation-cell division (RND)-type efflux pumps in Enterobacter cloacae complex. Antimicrob. Agents Chemother. 2016, 60, 2373–2382. [Google Scholar] [CrossRef] [Green Version]
- Mezzatesta, M.; Gona, F.; Stefani, S. Enterobacter cloacae complex: Clinical impact and emerging antibiotic resistance. Future Microbiol. 2012, 7, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Bina, X.R.; Philippart, J.A.; Bina, J.E. Effect of the efflux inhibitors 1-(1-naphthylmethyl)-piperazine and phenyl-arginine-β-naphthylamide on antimicrobial susceptibility and virulence factor production in Vibrio cholerae. J. Antimicrob. Chemother. 2009, 63, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahamoud, A.; Chevalier, J.; Davin-regli, A.; Barbe, J.; Pages, J.-M. Quinoline Derivatives as Promising Inhibitors of Antibiotic Efflux Pump in Multidrug Resistant Enterobacter Aerogenes Isolates. Curr. Drug Targets 2006, 7, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Ghisalberti, D.; Mahamoud, A.; Chevalier, J.; Baitiche, M.; Martino, M.; Pagès, J.M.; Barbe, J. Chloroquinolines block antibiotic efflux pumps in antibiotic-resistant Enterobacter aerogenes isolates. Int. J. Antimicrob. Agents 2006, 27, 565–569. [Google Scholar] [CrossRef]
- Chevalier, J.; Atifi, S.; Eyraud, A.; Mahamoud, A.; Barbe, J.; Pagès, J.M. New pyridoquinoline derivatives as potential inhibitors of the fluoroquinolone efflux pump in resistant enterobacter aerogenes strains. J. Med. Chem. 2001, 44, 4023–4026. [Google Scholar] [CrossRef]
- Malléa, M.; Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Brouant, P.; Barbe, J.; Pagès, J.M. Alkylaminoquinolines inhibit the bacterial antibiotic efflux pump in multidrug-resistant clinical isolates. Biochem. J. 2003, 805, 801–805. [Google Scholar] [CrossRef] [Green Version]
- Gallo, S.; Che, J.; Mahamoud, A.; Eyraud, A.; Barbe, J. 4-alkoxy and 4-thioalkoxyquinoline derivatives as chemosensitizers for the chloramphenicol-resistant clinical Enterobacter aerogenes 27 strain. Int. J. Antimicrob. Agents 2003, 22, 270–273. [Google Scholar] [CrossRef]
- Bohnert, J.A.; Kern, W. Selected Arylpiperazines Are Capable of Reversing Multidrug Resistance in Escherichia coli Overexpressing RND Efflux Pumps. Antimicrob. Agents Chemother. 2005, 49, 849–852. [Google Scholar] [CrossRef] [Green Version]
- Willers, C.; Wentzel, J.F.; du Plessis, L.H.; Gouws, C.; Hamman, J.H. Efflux as a mechanism of antimicrobial drug resistance in clinical relevant microorganisms: The role of efflux inhibitors. Expert Opin. Ther. Targets 2017, 21, 23–36. [Google Scholar] [CrossRef]
- Nakashima, R.; Sakurai, K.; Yamasaki, S.; Hayashi, K.; Nagata, C.; Hoshino, K.; Onodera, Y.; Nishino, K.; Yamaguchi, A. Structural basis for the inhibition of bacterial multidrug exporters. Nature 2013, 500, 102. [Google Scholar] [CrossRef]
- Opperman, T.J.; Kwasny, S.M.; Kim, H.S.; Nguyen, S.T.; Houseweart, C.; D’Souza, S.; Walker, G.C.; Peet, N.P.; Nikaido, H.; Bowlin, T.L. Characterization of a novel pyranopyridine inhibitor of the AcrAB efflux pump of Escherichia coli. Antimicrob. Agents Chemother. 2014, 58, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, D.C.; Stremick, C.A.; Slipski, C.J.; Turner, R.J. Secondary multidrug efflux pump mutants alter Escherichia coli biofilm growth in the presence of cationic antimicrobial compounds. Res. Microbiol. 2017, 168, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Wang, H.; Zou, L.; Zhang, A.; Yang, X.; Guan, Z. Evaluation and Target Validation of Indole Derivatives as Inhibitors of the AcrAB-TolC Efflux Pump. Biosci. Biotechnol. Biochem. 2010, 74, 2237–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohene-Agyei, T.; Mowla, R.; Rahman, T.; Venter, H. Phytochemicals increase the antibacterial activity of antibiotics by acting on a drug efflux pump. Microbiologyopen 2014, 3, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Takács, D.; Cerca, P.; Martins, A.; Riedl, Z.; Hajós, G.; Molnár, J.; Viveiros, M.; Couto, I.; Amaral, L. Evaluation of forty new phenothiazine derivatives for activity against intrinsic efflux pump systems of reference Escherichia coli, Salmonella Enteritidis, Enterococcus faecalis and Staphylococcus aureus strains. In Vivo 2011, 25, 719–724. [Google Scholar] [PubMed]
- Amaral, L.; Spengler, G.; Martins, A.; Armada, A.; Handzlik, J.; Kiec-Kononowicz, K.; Molnar, J. Inhibitors of Bacterial Efflux Pumps that also Inhibit Efflux Pumps of Cancer Cells. Anticancer Res. 2012, 2958, 2947–2957. [Google Scholar]
- Bohnert, J.A.; Schuster, S.; Kern, W.V. Pimozide Inhibits the AcrAB-TolC Efflux Pump in Escherichia coli. Open Microbiol. J. 2013, 7, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yao, Q.; Pan, X.C.; Wang, N.; Zhang, R.; Li, J.; Ding, G.; Liu, X.; Wu, C.; Ran, D.; et al. Artesunate enhances the antibacterial effect of β-lactam antibiotics against Escherichia coli by increasing antibiotic accumulation via inhibition of the multidrug efflux pump system AcrAB-TolC. J. Antimicrob. Chemother. 2011, 66, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Subhadra, B.; Kim, D.H.; Woo, K.; Surendran, S.; Choi, C.H. Control of Biofilm Formation in Healthcare: Recent Advances Exploiting Quorum-Sensing Interference Strategies and Multidrug Efflux. Materials 2018, 11, 1679. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, E.P.; Beis, K.; Cameron, A.D.; Iwata, S. Overcoming the challenges of membrane protein crystallography. Curr. Opin. Struct. Biol. 2008, 18, 581–586. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm Infections Site | Clinical Manifestations | Bacteria Species | References |
|---|---|---|---|
| Endocarditis (heart) | Patients with or without prosthetic heart valves or pacemaker, who have intermittent fever and bacteraemia with an identical pathogen. | Streptococcus species Enterococcus species Staphylococcus aureus Coagulase-negative Staphylococci | [19] [20] [21] [22] |
| Biofilm infection in CF/chronic obstructive pulmonary disease (COPD) (lungs) | Presence of bacteria detected in sputum. | Pseudomonas aeruginosa | [23] (CF) [24] (COPD) |
| Intravenous catheter | Patients with central venous catheter or haemodialysis catheter, who exhibit recurrent bacteraemia with an identical pathogen. | Escherichia coli Coagulase-negative Staphylococci Klebsiella pneumoniae Pseudomonas aeruginosa Enterococcus aerogenes | [25] [26] [25] [25] [25] |
| Orthopaedic infections (musculoskeletal system) | Local chronic pain and sign of prostheses loosening in patients with joint prostheses or orthopaedic fixation devices. | Staphylococcus aureus Coagulase-negative Staphylococci | [27] [28] |
| Urinary catheter | Recurrent urinary tract infections with the same pathogens in patients’ urinary catheter. | Klebsiella pneumonia Acinetobacter baumannii Candida species Enterococcus species Enterobacter species | [29] [30] [31] [32] [33] |
| Chronic wounds | Patients with chronic wounds suffering from recurrent wound infections. | Staphylococcus aureus Pseudomonas. aeruginosa | [34] [34] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reza, A.; Sutton, J.M.; Rahman, K.M. Effectiveness of Efflux Pump Inhibitors as Biofilm Disruptors and Resistance Breakers in Gram-Negative (ESKAPEE) Bacteria. Antibiotics 2019, 8, 229. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040229
Reza A, Sutton JM, Rahman KM. Effectiveness of Efflux Pump Inhibitors as Biofilm Disruptors and Resistance Breakers in Gram-Negative (ESKAPEE) Bacteria. Antibiotics. 2019; 8(4):229. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040229
Chicago/Turabian StyleReza, Akif, J. Mark Sutton, and Khondaker Miraz Rahman. 2019. "Effectiveness of Efflux Pump Inhibitors as Biofilm Disruptors and Resistance Breakers in Gram-Negative (ESKAPEE) Bacteria" Antibiotics 8, no. 4: 229. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040229