Antimicrobial Activity of Quinoline-Based Hydroxyimidazolium Hybrids

 , , , , , ,
, , , , , ,
Abstract
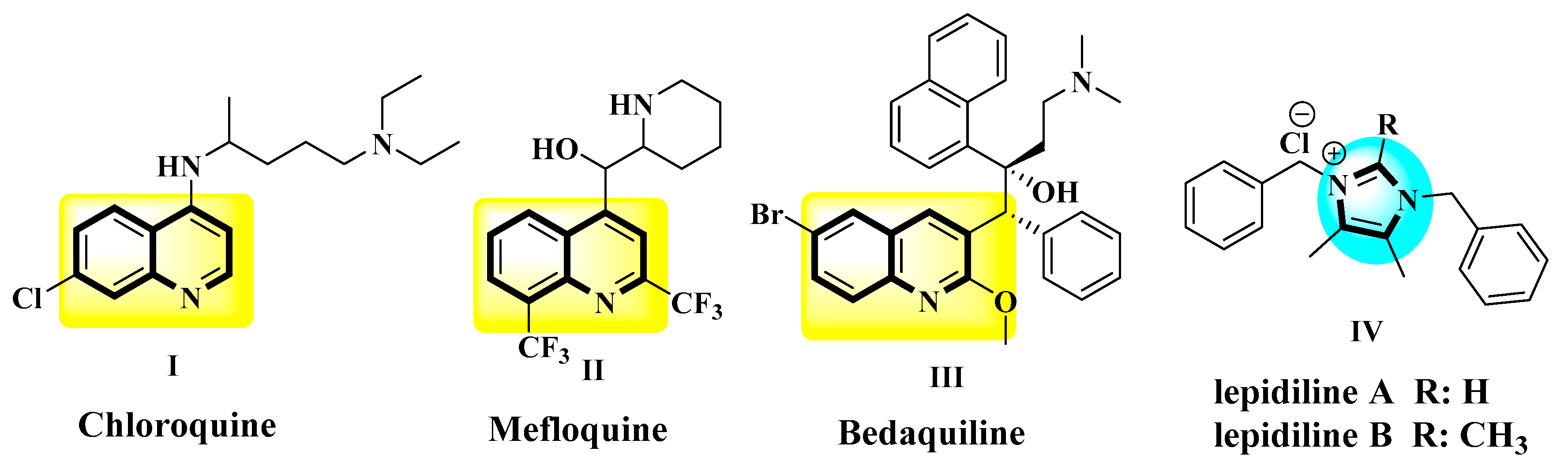
:1. Introduction
2. Results and Discussion
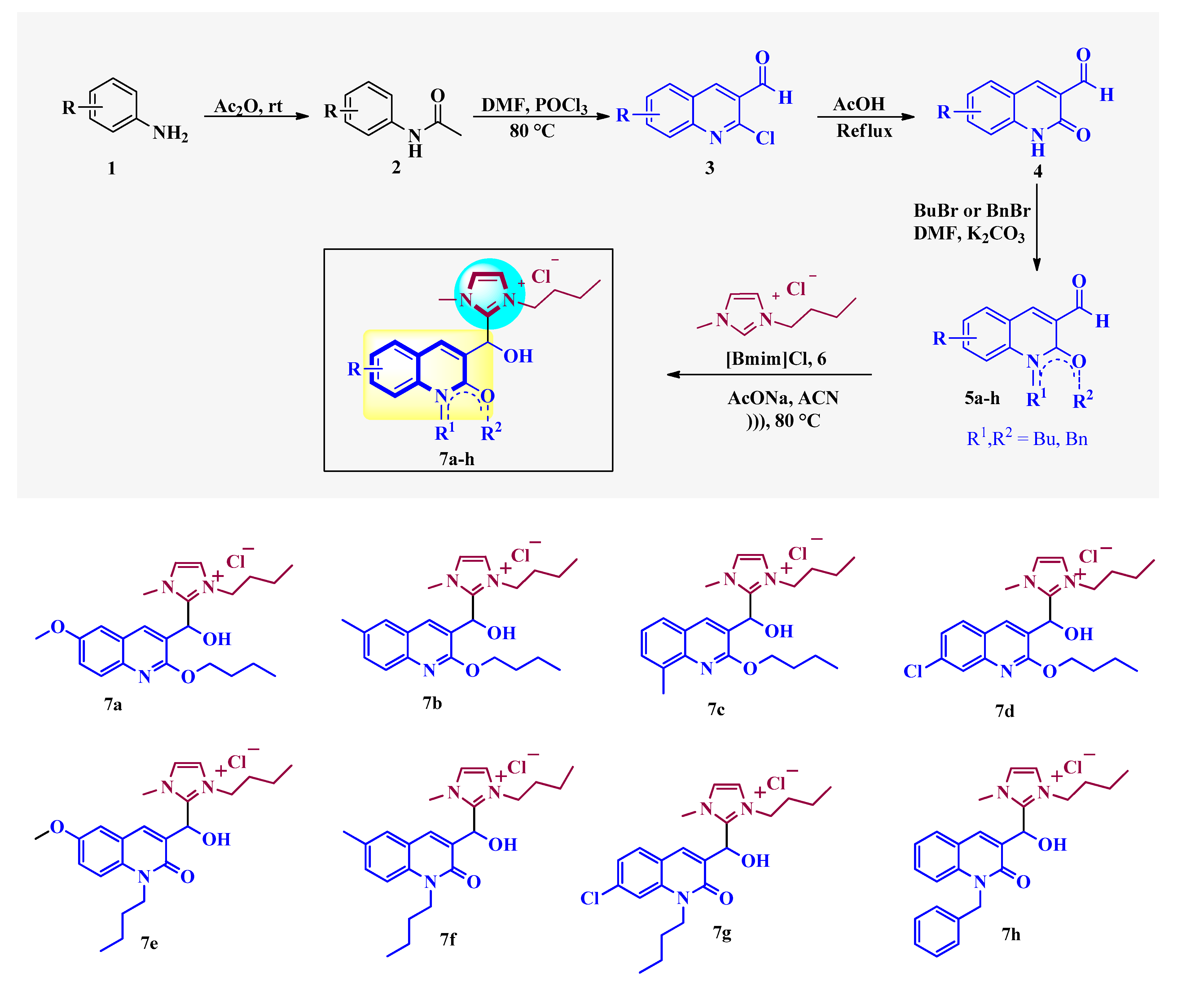
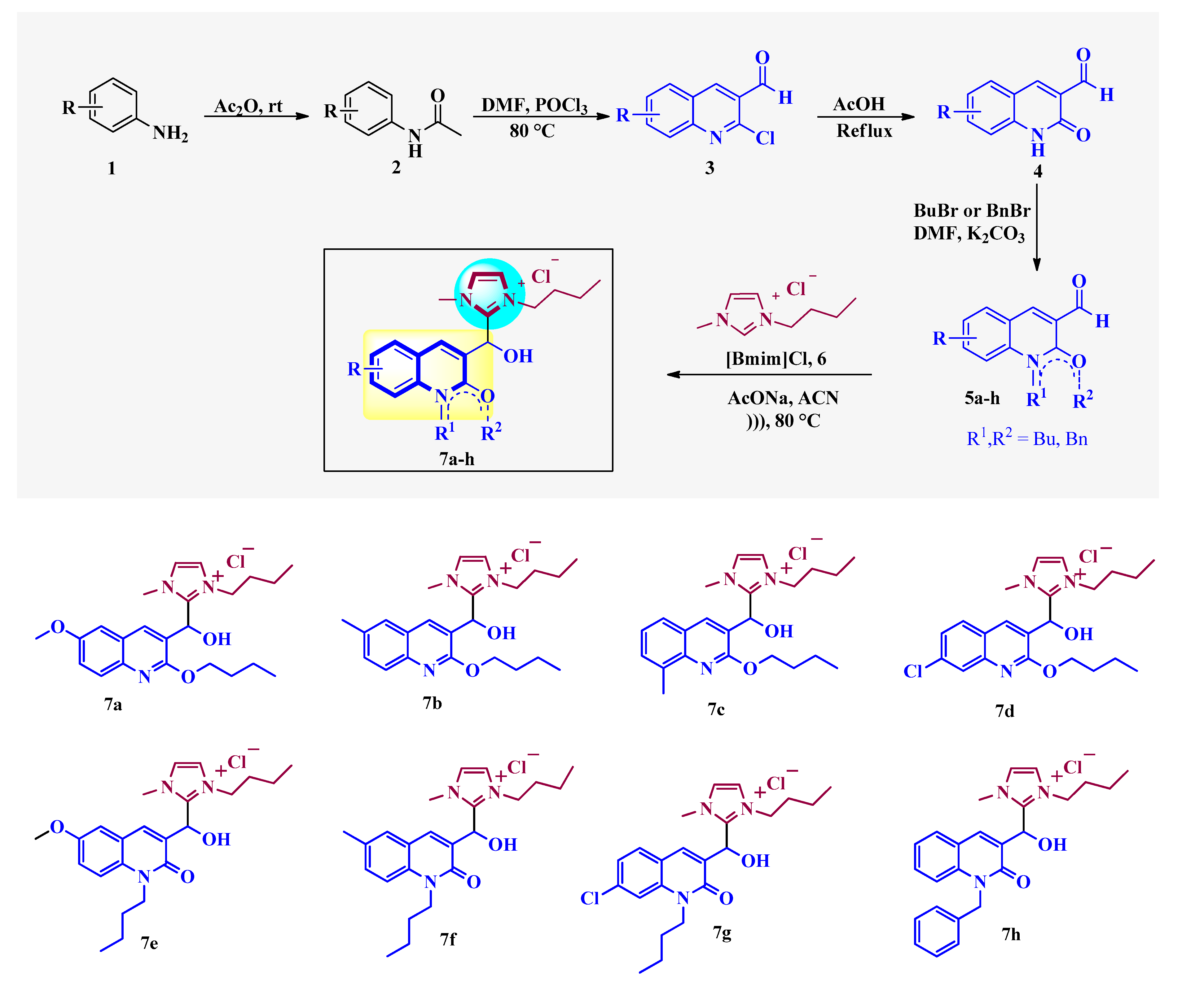
2.1. Chemistry
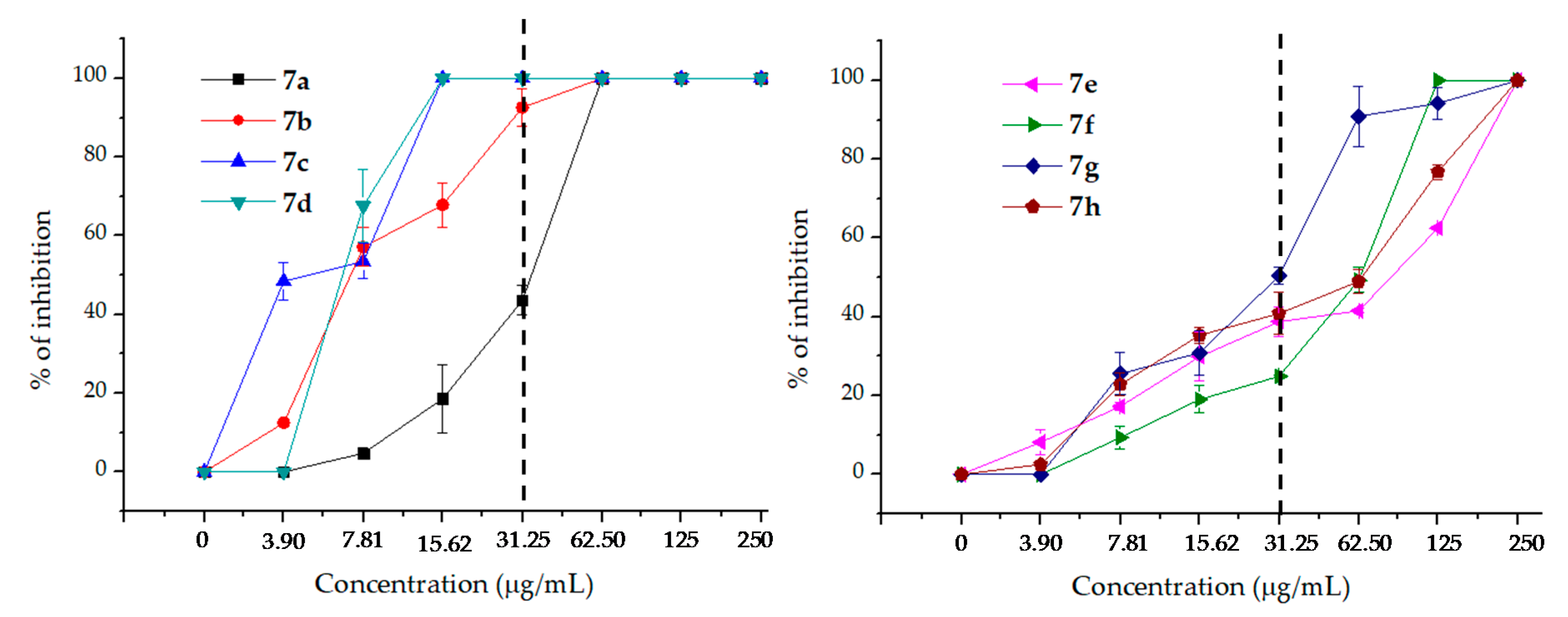
2.2. Antifungal Activity
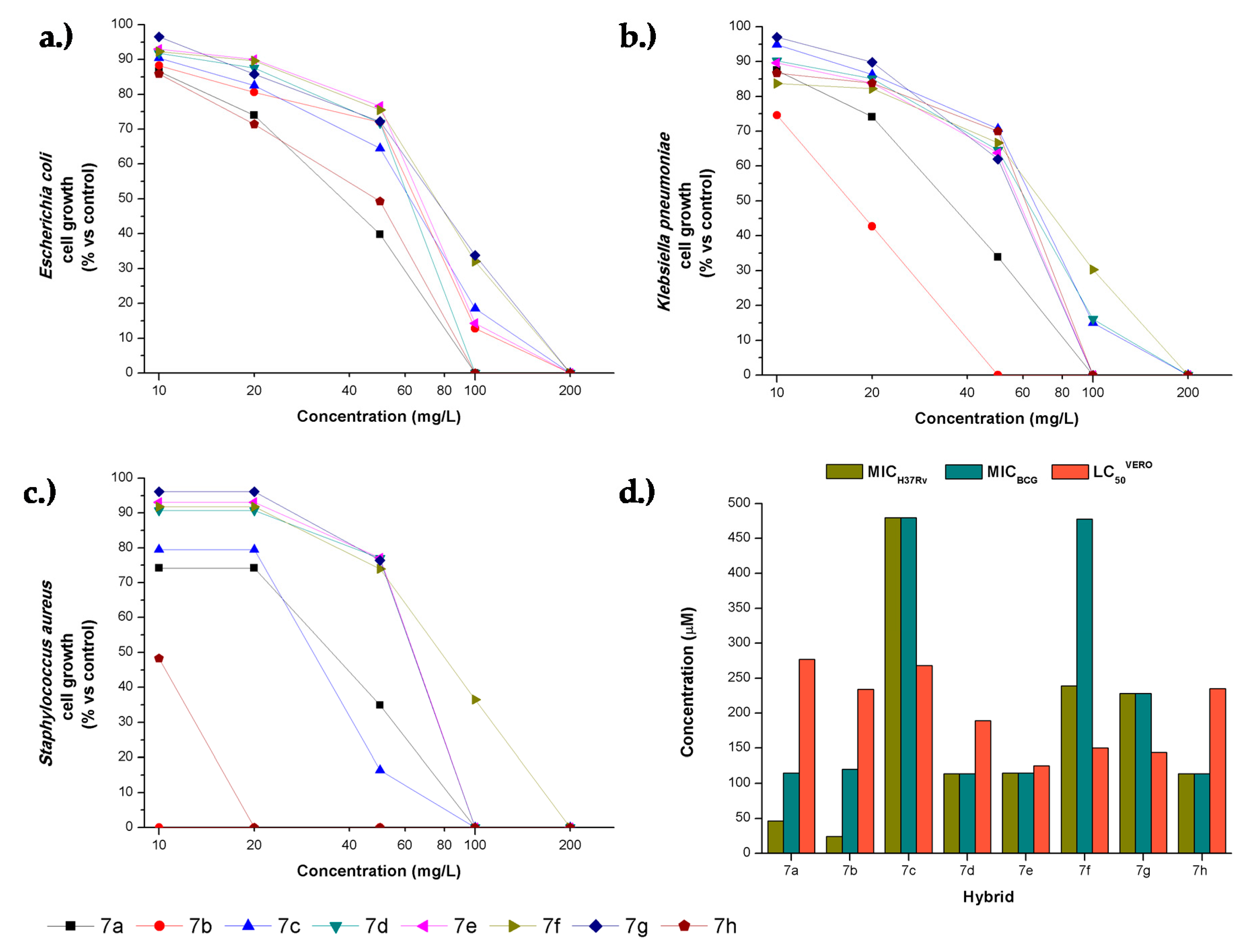
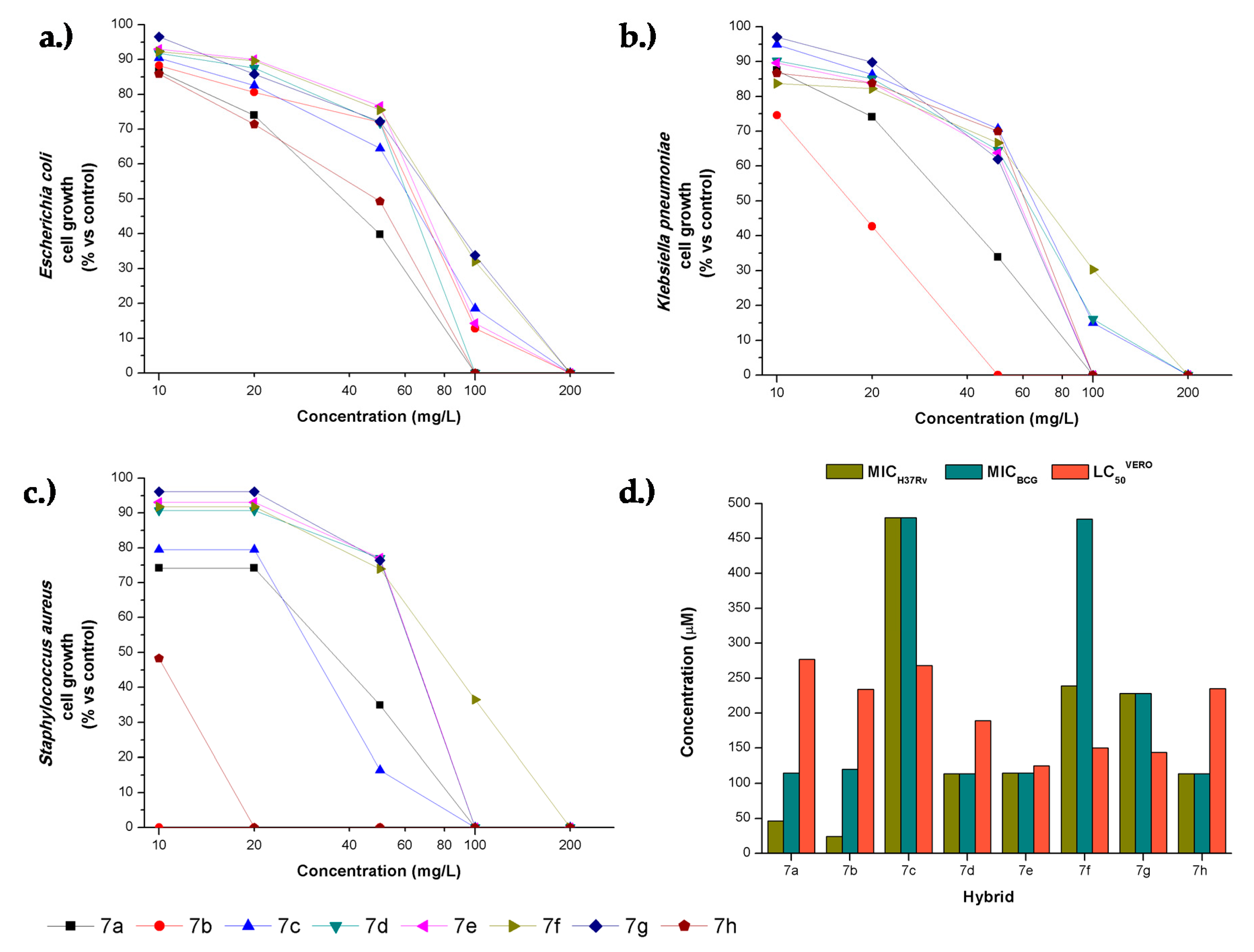
2.3. Antibacterial Activity
2.4. Cytotoxicity and Selective Index
3. Materials and Methods
3.1. Antifungal Activity
3.1.1. Microorganisms and Media
3.1.2. Antifungal Susceptibility Testing
3.1.3. Fungal Growth Inhibition Percentage Determination
3.2. Antituberculosis and Antibacterial Activity
3.3. Cytotoxicity on VERO Cell Line
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andersson, D.I.; Diarmaid, H. Antibiotic resistance and its cost: is it possible to reverse resistance? Nat. Microbiol. Rev. 2010, 8, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Geddes-McAlister, J.; Shapiro, R.S. New pathogens, new tricks: Emerging, drug-resistant fungal pathogens and future prospects for antifungal therapeutics. Ann. N. Y. Acad. Sci. 2019, 1435, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, K.; Bhagat, J.; Gupta, M.K.; Singh, J.V.; Gulati, H.K.; Singh, A.; Kaur, K.; Kaur, G.; Sharma, S.; Rana, A.; et al. Design, synthesis, antimicrobial evaluation, and molecular modeling studies of novel indolinedione–coumarin molecular hybrids. ACS Omega 2019, 4, 8720–8730. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Trivedi, A.; Pandit, U.; Dodiya, A.; Kameswara Rao, V.; Desai, P. Hybrid bioactive heterocycles as potential antimicrobial agents: a review. Mini Rev. Med. Chem. 2016, 16, 1500–1526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, S.; Ba, Y.; Xu, Z. 1,2,4-Triazole-quinoline/quinolone hybrids as potential anti-bacterial agents. Eur. J. Med. Chem. 2019, 174, 1–8. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129–145. [Google Scholar]
- World Health Organization. Guidelines for the Treatment of Malaria; World Health Organization: Geneva, Switzerland, 2015; ISBN 92-4-154912-2. [Google Scholar]
- Bisacchi, G.S. Origins of the quinolone class of antibacterials: An expanded “discovery story”. J. Med. Chem. 2015, 58, 4874–4882. [Google Scholar] [CrossRef]
- Hooper, D.C.; Wolfson, J.S. Fluoroquinolone antimicrobial agents. N. Engl. J. Med. 1991, 324, 384–394. [Google Scholar]
- Koul, A.; Vranckx, L.; Dhar, N.; Göhlmann, H.W.H.; Özdemir, E.; Neefs, J.-M.; Schulz, M.; Lu, P.; Mørtz, E.; McKinney, J.D.; et al. Delayed bactericidal response of Mycobacterium tuberculosis to bedaquiline involves remodelling of bacterial metabolism. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Datta, S.; He, G.; Tomilov, A.; Sahdeo, S.; Denison, M.S.; Cortopassi, G. In vitro evaluation of mitochondrial function and estrogen signaling in cell lines exposed to the antiseptic cetylpyridinium chloride. Environ. Health Perspect. 2017, 125, 087015. [Google Scholar] [CrossRef]
- Khajavi, R.; Sattari, M.; Ashjaran, A. The antimicrobial effect of benzalkonium chloride on some pathogenic microbes observed on fibers of acrylic carpet. Pakistan J. Biol. Sci. 2007, 10, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, P.R.; Brogden, R.N.; Pinder, R.M.; Speight, T.M.; Avery, G.S. Clotrimazole: A review of its antifungal activity and therapeutic efficacy. Drugs 1975, 9, 424–447. [Google Scholar] [CrossRef] [PubMed]
- Cutsem, J.V. The antifungal activity of ketoconazole. Am. J. Med. 1983, 74, 9–15. [Google Scholar] [CrossRef]
- Sawyer, P.R.; Brogden, R.N.; Pinder, R.M.; Speight, T.M.; Avery, G.S. Miconazole: A review of its antifungal activity and therapeutic efficacy. Drugs 1975, 9, 406–423. [Google Scholar] [CrossRef]
- Cui, B.; Zheng, B.L.; He, K.; Zheng, Q.Y. Imidazole alkaloids from lepidium meyenii. J. Nat. Prod. 2003, 66, 1101–1103. [Google Scholar] [CrossRef]
- Elshaarawy, R.F.; Janiak, C. Toward new classes of potent antibiotics: Synthesis and antimicrobial activity of novel metallosaldach–imidazolium salts. Eur. J. Med. Chem. 2014, 75, 31–42. [Google Scholar] [CrossRef]
- Riduan, S.N.; Zhang, Y. Imidazolium salts and their polymeric materials for biological applications. Chem. Soc. Rev. 2013, 42, 9055–9070. [Google Scholar] [CrossRef]
- Wang, D.; Richter, C.; Rühling, A.; Hüwel, S.; Glorius, F.; Galla, H.-J. Anti-tumor activity and cytotoxicity in vitro of novel 4, 5-dialkylimidazolium surfactants. Biochem. Biophys. Res. Commun. 2015, 467, 1033–1038. [Google Scholar] [CrossRef]
- Laali, K.K.; Insuasty, D.; Abonia, R.; Insuasty, B.; Bunge, S.D. Novel quinoline–imidazolium adducts via the reaction of 2-oxoquinoline-3-carbaldehyde and quinoline-3-carbaldehydes with 1-butyl-3-methylimidazolium chloride [BMIM][Cl]. Tetrahedron Lett. 2014, 55, 4395–4399. [Google Scholar] [CrossRef]
- Meth-Cohn, O. The synthesis of pyridines, quinolines and other related systems by the Vilsmeier and the reverse Vilsmeier method. Heterocycles 1993, 35, 539–557. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Document M27A3. In Reference Method For Broth Dilution Antifungal Susceptibility Testing of Yeasts; CLSI: Wayne, PA, USA, 2008; pp. 1–25. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Document M38A2. In Reference Method For Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; CLSI: Wayne, PA, USA, 2008; pp. 1–35. [Google Scholar]
- Desai, N.C.; Rajpara, K.M.; Joshi, V.V.; Vaghani, H.V.; Satodiya, H.M. Synthesis, characterization and antimicrobial screening of hybrid molecules containing quinoline, pyrimidine and morpholine analogues. J. Chem. Sci. 2013, 125, 321–333. [Google Scholar] [CrossRef]
- Desai, N.C.; Rajpara, K.M.; Joshi, V.V. Synthesis and characterization of some new quinoline based derivatives endowed with broad spectrum antimicrobial potency. Bioorg. Med. Chem. Lett. 2012, 22, 6871–6875. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Batchu, H.; Srivastava, K.; Singh, P.; Shukla, P.K.; Batra, S. Synthesis and evaluation of new diaryl ether and quinoline hybrids as potential antiplasmodial and antimicrobial agents. Bioorg. Med. Chem. Lett. 2014, 24, 1719–1723. [Google Scholar] [CrossRef] [PubMed]
- Nayak, N.; Ramprasad, J.; Dalimba, U. Synthesis and antitubercular and antibacterial activity of some active fluorine containing quinoline–pyrazole hybrid derivatives. J. Fluor. Chem. 2016, 183, 59–68. [Google Scholar] [CrossRef]
- Keri, R.S.; Patil, S.A. Quinoline: A promising antitubercular target. Biomed. Pharmacother. 2014, 68, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Kaur, H.; Smith, P.; de Kock, C.; Chibale, K.; Balzarini, J. Quinoline–Pyrimidine Hybrids: Synthesis, Antiplasmodial Activity, SAR, and Mode of Action Studies. J. Med. Chem. 2014, 57, 435–448. [Google Scholar] [CrossRef]
- Guantai, E.M.; Ncokazi, K.; Egan, T.J.; Gut, J.; Rosenthal, P.J.; Bhampidipati, R.; Kopinathan, A.; Smith, P.J.; Chibale, K. Enone– and Chalcone–Chloroquinoline Hybrid Analogues: In Silico Guided Design, Synthesis, Antiplasmodial Activity, in Vitro Metabolism, and Mechanistic Studies. J. Med. Chem. 2011, 54, 3637–3649. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Avula, S.R.; Mishra, V.; Palnati, G.R.; Singh, L.R.; Singh, N.; Chhonker, Y.S.; Swami, P.; Bhatta, R.S.; Palit, G. Identification of quinoline-chalcone hybrids as potential antiulcer agents. Eur. J. Med. Chem. 2015, 89, 638–653. [Google Scholar] [CrossRef]
- Mantoani, S.; Chierrito, T.; Vilela, A.; Cardoso, C.; Martínez, A.; Carvalho, I. Novel Triazole-Quinoline Derivatives as Selective Dual Binding Site Acetylcholinesterase Inhibitors. Molecules 2016, 21, 193. [Google Scholar] [CrossRef]
- Camps, P.; Formosa, X.; Galdeano, C.; Muñoz-Torrero, D.; Ramírez, L.; Gómez, E.; Isambert, N.; Lavilla, R.; Badia, A.; Clos, M.V. Pyrano[3,2-c]quinoline−6-Chlorotacrine Hybrids as a Novel Family of Acetylcholinesterase- and β-Amyloid-Directed Anti-Alzheimer Compounds. J. Med. Chem. 2009, 52, 5365–5379. [Google Scholar] [CrossRef]
- Guzman, J.D.; Gupta, A.; Evangelopoulos, D.; Basavannacharya, C.; Pabon, L.C.; Plazas, E.A.; Muñoz, D.R.; Delgado, W.A.; Cuca, L.E.; Ribon, W. Anti-tubercular screening of natural products from Colombian plants: 3-methoxynordomesticine, an inhibitor of MurE ligase of Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2010, 65, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Evangelopoulos, D.; Bhakta, S. Rapid Methods for Testing Inhibitors of Mycobacterial Growth. In Antibiotic Resistance Protocols, 2nd ed.; Gillespie, S.H., McHugh, T.D., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; pp. 193–201. ISBN 978-1-60327-279-7. [Google Scholar]
- Van Meerloo, J.; Kaspers, G.J.L.; Cloos, J. Cell Sensitivity Assays: The MTT Assay. In Cancer Cell Culture: Methods and Protocols; Cree, I.A., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; pp. 237–245. ISBN 978-1-61779-080-5. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Hybrid | R1 | R2 | R3 | R4 | Ca | Cn | Afu | Afl | Ani |
|---|---|---|---|---|---|---|---|---|---|---|
 | 7a | OMe | H | H | - | 125 | 62.5 | i | i | i |
| 7b | Me | H | H | - | 62.5 | 62.5 | i | i | i | |
| 7c | H | H | Me | - | 62.5 | 15.6 | 62.5 | 62.5 | 62.5 | |
| 7d | H | Cl | H | - | 62.5 | 15.6 | 62.5 | 62.5 | 62.5 | |
 | 7e | OMe | H | H | Bu | i | 250 | i | i | i |
| 7f | Me | H | H | Bu | 250 | 125 | i | i | i | |
| 7g | H | Cl | H | Bu | 250 | 62.5 | i | i | i | |
| 7h | H | H | H | Bn | i | 250 | 250 | 250 | 250 | |
| AmpB | 0.78 | 0.25 | 0.50 | 0.50 | 0.50 | |||||
| Comp. | Concentration in µg/mL | MIC in µg/mL | ||||||
|---|---|---|---|---|---|---|---|---|
| 250 | 125 | 62.50 | 31.25 | 15.62 | 7.81 | 3.90 | ||
| 7a | 100 | 100 | 100 | 43.6 ± 3.86 | 18.60 ± 8.59 | 4.72 ± 1.39 | 0 | 62.5 |
| 7b | 100 | 100 | 100 | 92.58 ± 4.81 | 67.84 ± 5.66 | 57.14 ± 4.93 | 12.54 ± 0.36 | 31.25 |
| 7c | 100 | 100 | 100 | 100 | 100 | 53.45 ± 4.42 | 48.41 ± 4.63 | 15.62 |
| 7d | 100 | 100 | 100 | 100 | 100 | 67.59 ± 9.19 | 0 | 15.62 |
| 7e | 100 | 62.46 ± 0.30 | 41.46 ± 0.58 | 38.79 ± 3.79 | 29.92 ± 6.13 | 17.19 ± 0.91 | 8.23 ± 3.18 | 250 |
| 7f | 100 | 100 | 49.28 ± 3.20 | 24.91 ± 0.15 | 19.01 ± 3.40 | 9.34 ± 2.90 | 0 | 62.5 |
| 7g | 100 | 94.15 ± 4.15 | 90.90 ± 7.60 | 50.44 ± 2.09 | 30.94 ± 5.61 | 25.50 ± 5.35 | 0 | 250 |
| 7h | 100 | 76.77 ± 1.91 | 48.99 ± 3.00 | 46.84 ±5.32 | 35.32 ± 2.02 | 22.76 ± 2.87 | 2.52 ± 0.56 | 250 |
| AmpB | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 0.25 |
| Hybrid | MICs in µg/mL (µM) | ||||
|---|---|---|---|---|---|
| E. coli | K. pneumoniae | S. aureus | M. bovis BCG | M. tuberculosis H37Rv | |
| 7a | 100 (231) | 100 (231) | 100 (231) | 50 (115) | 20 (46) |
| 7b | 200 (480) | 50 (120) | 2 (5) | 50 (120) | 10 (24) |
| 7c | 200 (480) | 200 (480) | 100 (240) | 200 (480) | 200 (480) |
| 7d | 100 (229) | 200 (457) | 100 (229) | 50 (114) | 50 (114) |
| 7e | 200 (462) | 100 (231) | 100 (231) | 50 (115) | 50 (115) |
| 7f | 200 (478) | 200 (478) | 200 (478) | 200 (478) | 100 (239) |
| 7g | 200 (228) | 100 (114) | 100 (114) | 100 (228) | 100 (228) |
| 7h | 100 (228) | 100 (228) | 20 (47) | 50 (114) | 50 (114) |
| chloramphenicol | 20 (62) | 20 (62) | 20 (62) | nd | nd |
| isoniazid | nd | nd | nd | 0.05 (0.36) | 0.05 (0.36) |
| Hybrid | Selectivity Index (SI) (SI = LC50/MIC) | |||||
|---|---|---|---|---|---|---|
| Cytotoxicity LC50-Vero cells | E. coli | K. pneumoniae | S. aureus | M. bovis BCG | M. tuberculosis H37Rv | |
| 7a | 277 ± 14.6 | 2.77 | 2.77 | 2.77 | 5.54 | 13.9 |
| 7b | 234 ± 4 | 1.17 | 4.68 | 117 | 4.68 | 23.4 |
| 7c | 268 ± 5.69 | 1.34 | 1.34 | 2.68 | 1.34 | 1.34 |
| 7d | 189 ± 14.5 | 1.89 | 0.94 | 1.89 | 3.78 | 3.78 |
| 7e | 125 ± 22 | 0.62 | 1.25 | 1.25 | 2.5 | 2.50 |
| 7f | 150 ± 12.2 | 0.75 | 0.75 | 0.75 | 0.75 | 1.50 |
| 7g | 144 ± 8.9 | 1.44 | 1.44 | 1.44 | 1.44 | 1.44 |
| 7h | 235 ± 6.5 | 2.35 | 2.35 | 11.8 | 4.70 | 4.70 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Insuasty, D.; Vidal, O.; Bernal, A.; Marquez, E.; Guzman, J.; Insuasty, B.; Quiroga, J.; Svetaz, L.; Zacchino, S.; Puerto, G.; et al. Antimicrobial Activity of Quinoline-Based Hydroxyimidazolium Hybrids. Antibiotics 2019, 8, 239. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040239
Insuasty D, Vidal O, Bernal A, Marquez E, Guzman J, Insuasty B, Quiroga J, Svetaz L, Zacchino S, Puerto G, et al. Antimicrobial Activity of Quinoline-Based Hydroxyimidazolium Hybrids. Antibiotics. 2019; 8(4):239. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040239
Chicago/Turabian StyleInsuasty, Daniel, Oscar Vidal, Anthony Bernal, Edgar Marquez, Juan Guzman, Braulio Insuasty, Jairo Quiroga, Laura Svetaz, Susana Zacchino, Gloria Puerto, and et al. 2019. "Antimicrobial Activity of Quinoline-Based Hydroxyimidazolium Hybrids" Antibiotics 8, no. 4: 239. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040239