Effects of Early Intervention with Antibiotics and Maternal Fecal Microbiota on Transcriptomic Profiling Ileal Mucusa in Neonatal Pigs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Donor Material Preparation
2.3. Animal Experiment and Sampling
2.4. Morphometric Analysis
2.5. Serum Cytokines Analysis
2.6. RNA Isolation and Sequencing
2.7. Quantitative Real-Time PCR
2.8. Data Analysis
3. Results
3.1. Ileal Morphological Structure
3.2. Serum Inflammatory Cytokines Concentrations
3.3. Expression of Genes Related to Intestinal Epithelial Tight Junction Proteins, Toll-Like Receptors, and Inflammatory Cytokines in the Ileum
3.4. Expression of Genes Related to Intestinal Development in the Ileum
3.5. Transcriptomic Profiling of the Ileal Mucosa
3.6. Validation of RNA-Sequencing by Quantitative Real-Time PCR
4. Discussion
4.1. Effects of Early Maternal Fecal Microbiota and Antibiotics Intervention on Intestinal Morphology and Barrier Function in the Ileum
4.2. Effects of Early Maternal Fecal Microbiota and Antibiotics Intervention on Transcriptomic Profiles in the Ileum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kamada, N.; Núñez, G. Regulation of the immune system by the resident intestinal bacteria. Gastroenterology 2014, 146, 1477–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.H.F.; Nielsen, D.S.; Kverka, M.; Zakostelska, Z.; Klimesova, K.; Hudcovic, T.; Hansen, A.K. Patterns of early gut colonization shape future immune responses of the host. PLoS ONE 2012, 7, e34043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandtzaeg, P. Development and basic mechanisms of human gut immunity. Nutr. Rev. 1998, 56, S5–S18. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schokker, D.; Zhang, J.; Zhang, L.L.; Vastenhouw, S.A.; Heilig, H.G.; Smidt, H.; Rebel, J.M.; Smits, M.A. Early-life environmental variation affects intestinal microbiota and immune development in new-born piglets. PLoS ONE 2014, 9, e100040. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Cernada, M.; Baüerl, C.; Vento, M.; Pérez-Martínez, G. Microbial ecology and host-microbiota interactions during early life stages. Gut Microbes 2012, 3, 352–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, S.K.; Xu, B.; Nyachoti, C.M.; Giesting, D.W.; Krause, D.O. Evaluation of alternatives to antibiotics using an Escherichia coli K88+ model of piglet diarrhea: Effects on gut microbial ecology. J. Anim. Sci. 2008, 86, 836–847. [Google Scholar] [CrossRef]
- Nathan, C. Antibiotics at the crossroads. Nature 2004, 431, 899–902. [Google Scholar] [CrossRef]
- Cho, I.; Yamanishi, S.; Cox, L.; Methé, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef]
- Morel, F.B.; Oozeer, R.; Piloquet, H.; Moyon, T.; Pagniez, A.; Knol, J.; Darmaun, D.; Michel, C. Preweaning modulation of intestinal microbiota by oligosaccharides or amoxicillin can contribute to programming of adult microbiota in rats. Nutrition 2015, 31, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, D.A.; Huse, S.M.; Morrison, H.G.; Schmidt, T.M.; Sogin, M.L.; Young, V.B. Reproducible community dynamics of the gastrointestinal microbiota following antibiotic perturbation. Infect. Immun. 2009, 77, 2367–2375. [Google Scholar] [CrossRef] [Green Version]
- Connelly, S.; Subramanian, P.; Hasan, N.A.; Colwell, R.R.; Kaleko, M. Distinct consequences of amoxicillin and ertapenem exposure in the porcine gut microbiome. Anaerobe 2018, 53, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Schumann, A. Neonatal antibiotic treatment alters gastrointestinal tract developmental gene expression and intestinal barrier transcriptome. Physiol. Genom. 2005, 23, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Khoruts, A.; Sadowsky, M.J. Understanding the mechanisms of fecal microbiota transplantation. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quraishi, M.N.; Widlak, M.; Bhala, N.; Moore, D.; Price, M.; Sharma, N.; Iqbal, T.H. Systematic review with meta-analysis: The efficacy of fecal microbiota transplantation for the treatment of recurrent and refractory Clostridium difficile infection. Aliment. Pharm. Ther. 2017, 46, 479–493. [Google Scholar] [CrossRef] [Green Version]
- McCormack, U.M.; Curião, T.; Wilkinson, T.; Metzler-Zebeli, B.U.; Reyer, H.; Ryan, T.; Calderon-Diaz, J.A.; Crispie, F.; Cotter, P.D.; Creevey, C.J.; et al. Fecal microbiota transplantation in gestating sows and neonatal offspring alters lifetime intestinal microbiota and growth in offspring. MSystems 2018, 3, e00134-17. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, J.; Ren, E.; Su, Y.; Zhu, W. Co-occurrence of early gut colonization in neonatal piglets with microbiota in the maternal and surrounding delivery environments. Anaerobe 2018, 49, 30–40. [Google Scholar] [CrossRef]
- Lin, C.; Wan, J.; Su, Y.; Zhu, W. Effects of early intervention with maternal fecal microbiota and antibiotics on the gut microbiota and metabolite profiles of piglets. Metabolites 2018, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.J.; Weingarden, A.R.; Sadowsky, M.J.; Khoruts, A. Standardized frozen preparation for transplantation of fecal microbiota for recurrent clostridium difficile infection. Am. J. Gastroenterol. 2012, 107, 761–767. [Google Scholar] [CrossRef]
- Ait-Ali, T.; Díaz, I.; Soldevila, F.; Cano, E.; Li, Y.; Wilson, A.D.; Giotti, B.; Archibald, A.L.; Mateu, L.; Darwich, L. Distinct functional enrichment of transcriptional signatures in pigs with high and low IFN-gamma responses after vaccination with a porcine reproductive and respiratory syndrome virus (PRRSV). Vet. Res. 2017, 47, 104. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jin, H.; Chen, P.; Li, Z.; Meng, X.; Liu, M.; Li, S.; Shi, D.; Xiao, Y.; Wang, X.; et al. Haemophilus parasuis infection activates the NF-κB pathway in PK-15 cells through IκB degradation. Vet. Microbiol. 2012, 160, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, M.D.; Ross, J.W.; Ritchey, J.W.; Desilva, U.; Stein, D.R.; Geisert, R.D.; White, F.J. Effects of aberrant estrogen on the endometrial transcriptional profile in pigs. Reprod. Toxicol. 2012, 34, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.R.; Hansen, T.V.A.; Krych, L.; Ahmad, H.F.B.; Nielsen, D.S.; Skovgaard, K.; Thamsborg, S.M. Dietary cinnamaldehyde enhances acquisition of specific antibodies following helminth infection in pigs. Vet. Immunol. Immunopathol. 2017, 189, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Okutani, M.; Tsukahara, T.; Kato, Y.; Fukuta, K.; Inoue, R. Gene expression profiles of CD4/CD8 double-positive T cells in porcine peripheral blood. Anim. Sci. J. 2018, 89, 979–987. [Google Scholar] [CrossRef]
- Shinkai, H.; Morozumi, T.; Toki, D.; Eguchi-Ogawa, T.; Muneta, Y.; Awata, T.; Uenishi, H. Genomic structure of eight porcine chemokine receptors and intergene sharing of an exon between CCR1 and XCR1. Gene 2005, 349, 55–66. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Biogeosciences 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, A.; Bar-Shira, E.; Sklan, D. Ontogeny of gut associated immune competence in the chick. World Poult. Sci. J. 2003, 59, 209–219. [Google Scholar] [CrossRef]
- Kroismayr, A.; Sehm, J.; Pfaffl, M.W.; Schedle, K.; Plitzner, C.; Windisch, W. Effects of avilamycin and essential oils on mRNA expression of apoptotic and inflammatory markers, and gut morphology of piglets. Czech J. Anim. Sci. 2008, 53, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Diao, H.; Yan, H.L.; Xiao, Y.; Yu, B.; Zheng, P.; He, J.; Yu, J.; Mao, X.B.; Chen, D.W. Modulation of intestine development by fecal microbiota transplantation in suckling pigs. RSC Adv. 2018, 8, 8709–8720. [Google Scholar] [CrossRef] [Green Version]
- Förster, C. Tight junctions and the modulation of barrier function in disease. Histochem. Cell Biol. 2008, 130, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, E.C.; Murphy, C.; O’Neill, L.A.J.; Creagh, E.M. The role of TLRs, NLRs, and RLRs in mucosal innate immunity and homeostasis. Mucosal Immunol. 2009, 3, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Yu, K.; Mu, C.; Yang, Y.; Su, Y.; Zhu, W. Segment-specific responses of intestinal epithelium transcriptome to in-feed antibiotics in pigs. Physiol. Genom. 2017, 49, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.J. Role of chemokines in the biology of natural killer cells. J. Leukoc. Biol. 2002, 71, 173–183. [Google Scholar] [PubMed]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef] [PubMed]
- San Luis, B.; Sondgeroth, B.; Nassar, N.; Carpino, N. Sts-2 is a phosphatase that negatively regulates zeta-associated protein (ZAP)-70 and T cell receptor signaling pathways. J. Biol. Chem. 2011, 286, 15943–15954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mege, D.; Di Bartolo, V.; Germain, V.; Tuosto, L.; Michel, F.; Acuto, O. Mutation of tyrosines 492/493 in the kinase domain of ZAP-70 affects multiple T-cell receptor signaling pathways. J. Biol. Chem. 1996, 271, 32644–32652. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.C.; Irving, B.A.; Fraser, J.D.; Weiss, A. The zeta chain is associated with a tyrosine kinase and upon T-cell antigen receptor stimulation associates with ZAP-70, a 70-kDa tyrosine phosphoprotein. Proc. Natl. Acad. Sci. USA 1991, 88, 9166–9170. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
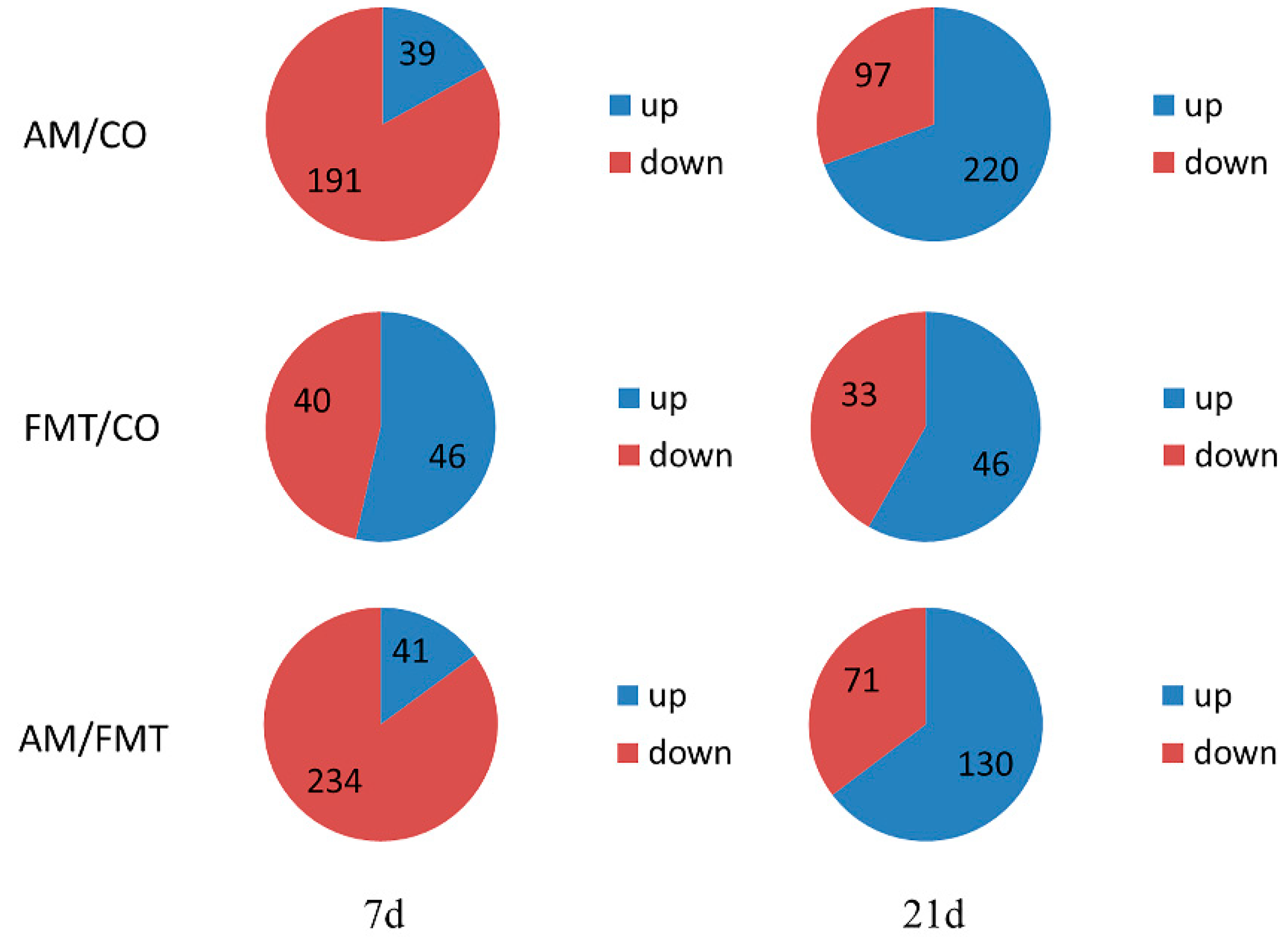{kind=link}
{kind=link}
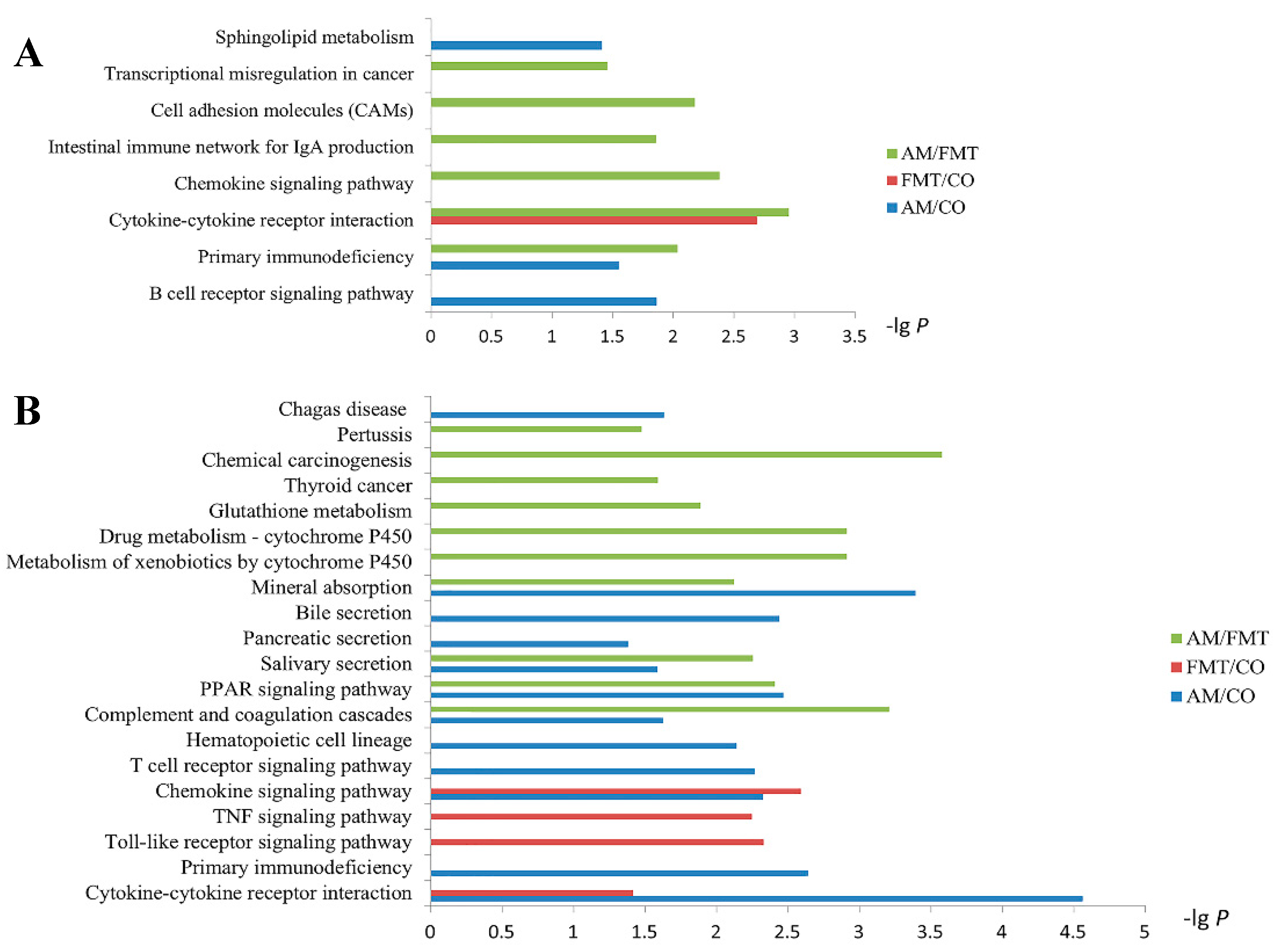{kind=link}
| Item | 7 d | 21 d | ||||
|---|---|---|---|---|---|---|
| CO | AM | FMT | CO | AM | FMT | |
| Villus height, μm | 694.65 (66.66) | 679.60 (26.00) | 607.97 (48.76) | 264.69 (3.57) | 271.55 (8.48) | 222.02 (44.56) |
| Crypt depth, μm | 73.45 (1.67) a | 63.14 (4.38) b | 56.92 (0.91) b | 73.34 (1.18) | 67.39 (1.01) | 72.62 (3.81) |
| Villus height/Crypt depth | 9.47 (0.95) | 10.90 (0.57) | 10.67 (0.77) | 3.61 (0.06) | 4.03 (0.13) | 3.85 (0.53) |
| Items, ng/mL | 7 d | 21 d | ||||
|---|---|---|---|---|---|---|
| AM | FMT | CO | AM | FMT | CO | |
| IFN-γ | 16.12 (1.17) | 17.65 (0.77) | 17.61 (1.35) | 13.26 (0.89) b | 19.35 (1.11) a | 17.66 (1.55) a |
| IL-2 | 1.27 (0.25) | 1.05 (0.12) | 1.35 (0.19) | 0.86 (0.14) | 0.80 (0.07) | 0.92 (0.1) |
| IL-6 | 0.820 (0.14) | 0.67 (0.12) | 0.91 (0.1) | 0.25 (0.051) | 0.19 (0.04) | 0.26 (0.05) |
| IL-8 | 0.26 (0.03) | 0.19 (0.03) | 0.18 (0.03) | 0.41 (0.05) ab | 0.47 (0.04) a | 0.34 (0.03) b |
| IL-10 | 2.39 (0.27) | 2.00 (0.28) | 2.78 (0.32) | 0.82 (0.12) | 0.64 (0.12) | 0.95 (0.15) |
| IL-12 | 0.46 (0.09) | 0.44 (0.06) | 0.55 (0.10) | 0.77 (0.04) | 0.74 (0.07) | 0.76 (0.07) |
| Gene | 7 d | 21 d | ||||
|---|---|---|---|---|---|---|
| AM/CO | FMT/CO | AM/FMT | AM/CO | FMT/CO | AM/FMT | |
| CCL4 | NO | NO | DOWN | UP | NO | NO |
| CCL5 | NO | NO | NO | UP | UP | NO |
| CXCL9 | NO | UP | NO | UP | UP | NO |
| CXCL11 | NO | NO | NO | UP | UP | NO |
| ZAP70 | NO | NO | NO | UP | NO | NO |
| CD8B | NO | NO | NO | UP | NO | NO |
| CD3G | NO | NO | NO | UP | UP | NO |
| CD3D | NO | NO | NO | UP | NO | NO |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Wan, J.; Lin, C.; Su, Y. Effects of Early Intervention with Antibiotics and Maternal Fecal Microbiota on Transcriptomic Profiling Ileal Mucusa in Neonatal Pigs. Antibiotics 2020, 9, 35. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010035
Xu R, Wan J, Lin C, Su Y. Effects of Early Intervention with Antibiotics and Maternal Fecal Microbiota on Transcriptomic Profiling Ileal Mucusa in Neonatal Pigs. Antibiotics. 2020; 9(1):35. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010035
Chicago/Turabian StyleXu, Rongying, Jiajia Wan, Chunhui Lin, and Yong Su. 2020. "Effects of Early Intervention with Antibiotics and Maternal Fecal Microbiota on Transcriptomic Profiling Ileal Mucusa in Neonatal Pigs" Antibiotics 9, no. 1: 35. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010035