1. Introduction
Antimicrobial resistance is one of the greatest concerns of public health worldwide since drug-resistant bacterial infections have increased risk of worse clinical outcomes and death. The infection caused by resistant bacteria results in lengthier stays in hospital and may need more intensive care or more expensive drugs [
1]. Antibacterial resistance can come from various mechanisms, including enzymatic degradation of antibiotics, alteration of antibiotic targets, blocking of drug entry, and increasing expression of efflux pumps (EPs) [
2].
EPs play a significant role in bacteria; six different families of EP systems enable bacteria to extrude various potentially toxic substances such as antibiotics, heavy metals, organic pollutants, and plant-produced compounds. Among these families we can differentiate: ATP-binding cassette (ABC) transporters, multidrug and toxic compound extrusion (MATE) transporters, small multidrug resistance (SMR) transporters, major facilitator superfamily (MFS) transporters, resistance nodulation cell division (RND) transporters, and proteobacterial antimicrobial compound efflux (PACE) transporters [
3]. MFS and MATE transporters are expressed mainly by Gram-positive bacteria, while RND transporters are found almost exclusively in Gram-negative bacteria [
4]. The most investigated RND efflux system is AcrAB-TolC, which constitutes the AcrB RND EP, TolC outer membrane protein, and AcrA serves as periplasmic adaptor protein [
5].
The formation of biofilms gives rise to many clinical problems and plays a role in antibacterial resistance. Biofilm is a dynamic structure built by multicellular bacterial community and its extracellular polymeric matrix. This environment not only provides bacteria with protection against drugs, host immune attacks, and environmental stresses, but also forms a platform of metabolic exchange and cellular communication [
6].
Quorum sensing (QS) is a cell–cell communication system enabling bacteria to regulate gene expression based on cell population density [
7]. Though QS bacteria can alter their phenotypes and behaviors, they may be able to form biofilms and express various virulence factors (such as EPs) resulting in an enhanced drug resistance [
8].
Metal complexes are already widely used in clinical settings and many antibiotics containing metal complexes have better pharmaceutical characteristics [
9,
10,
11]. Benzoxazole skeleton-containing molecules are also in the scope of research due to their antimicrobial and antiproliferative properties [
12,
13,
14]. The importance of benzoxazole core and 2-trifluoroacetonyl moiety of 2-trifluoroacetonylbenzoxazole
1 in order to express their antimicrobial activities has been previously highlighted [
15,
16,
17]. In recent work on 2-trifluoroacetonylbenzoxazole ligand
1 for metal complexes, Zn(II) complex
4 of ligand
1 formed a stable six-membered ring in a bidentate mode to consist of
1 with Zn(II) ion in 2:1 ratio with good antibacterial activities against Gram-positive pathogens [
18] and potent multidrug-resistance reversing activities in cancer cells [
19]. In this article, we report the multidrug resistance-modulating activities of 2-trifluoroacetonylbenzoxazole ligands (
1–3) and their metal complexes (
4–12) in bacteria related to the inhibition of QS and biofilm formation.
3. Discussion
Metal complexes could provide a beneficial tool for pharmaceuticals and there have been several compounds with anticancer, antimalarial, and anti-neurodegenerative activities in clinical trials. However, metal complexes as potential antibacterial agents gained less attention until now, despite the fact that they possess remarkable antimicrobial effects [
20]. Based on our results, the antibacterial effect of 2-fluoroacetonylbenzoxazole ligands could be increased by metal-based coordination complexes of these ligands. The metal complexes could be applied to reduce the virulence of bacteria (inhibition of QS and biofilm formation) and restore the sensitivity of multidrug resistant bacteria by modulating the overexpressed EP systems, and they could be administered together with antibiotics to improve the efficacy of antibacterial therapy.
As shown previously, transition metal complexes can demonstrate better antimicrobial activity than the ligands themselves. 2-substituted benzoxazoles are important scaffolds because they possess antimicrobial, antiviral, antifungal, and anticancer effects as well [
18].
Based on the results, it has been confirmed that the metal complexes exerted more pronounced antibacterial activity than the ligands: This tendency was observed in the case of the complexes of the 2-trifluoroacetonylbenzoxazole ligand 1 with Zn(II), Cu(II), Ni(II), Mg(II), and, furthermore, in the case of the complex of the ligand 3 with Zn(II) on the Gram-positive S. aureus MRSA and reference ATCC strains. However, ligand 2 and with Zn(II), furthermore, ligand 1 with Cu(II), had a more potent antibacterial effect (MIC: 3.125 µM) than the reference standard TET (MIC: 6.25 µM) or CIP (MIC: 12.5 µM) on the S. aureus MRSA strain. Ligand 1 with Zn(II), Ni(II), and Mg(II) were more effective (MIC: 6.25 µM) compared to CIP against S. aureus MRSA. The MIC of TET was 50 µM on E. coli AG100, but stronger growth inhibition was observed for ligand 1 with Zn(II) and Ag(II); MIC was 25 µM.
In three ligands 1–3, the order of antibacterial potency was 2 > 1> 3. Interestingly, ligand 3 with the lowest antibacterial activity could potentiate the activity of CIP on MRSA strain. The other ligands 1 and 2 could show the same activity in the form of metal complexes: the Zn(II) complex 5 of ligand 2 and the complexes of ligand 1 with Cu (7), Ni (8), Mg (9), and Fe (12). In addition, complexes 8 and 9 were effective in combination with CIP on E. coli AG100 as well.
In order to re-sensitize antibiotic-resistant bacteria, EP inhibitors could be used to overcome the resistant phenotype. Ligand
3 was a potent EP inhibitor on MRSA but had no activity on the sensitive
S. aureus reference strain and the Gram-negative
E. coli AG100 expressing the AcrAB-TolC efflux system. The reason for that is the different cell wall composition, because Gram-positives have a thick layer of peptidoglycan, and in the case of MRSA isolate, the distribution of essential proteins involved in lipid metabolism, and the expression of EPs and membrane proteins may be different compared to sensitive ATCC isolates [
21]. On the contrary, Gram-negatives possess a thin peptidoglycan layer, but because of the presence of the inner and outer membranes, tripartite efflux pump systems such as the AcrAB-TolC system can be overexpressed in resistant Gram-negative isolates. The Zn(II) complex
5 of ligand
2 and the Ag(II) complex
11 of ligand
1 inhibited the AcrAB-TolC efflux system in
E. coli AG100, and these complexes exhibited moderate toxicity on normal MRC-5 human embryonal fibroblast cells (IC
50 of 24.57 µM and 65.89 µM, respectively). According to previous studies [
18], the Ag(II) complex
11 was found to exhibit the strongest inhibitory activity against various Gram-negative bacteria, especially against
Pseudomonas aeruginosa, causing membrane permeabilization and degradation.
The bacterial virulence can be reduced if the bacterial communication and the virulence factors related to QS can be inhibited. The bacterial communication could be hampered by treatment with ligand
3, the Ag(II) complex
11 of ligand
1, and the Zn(II) complexes
4 and
5 of ligand
1 and
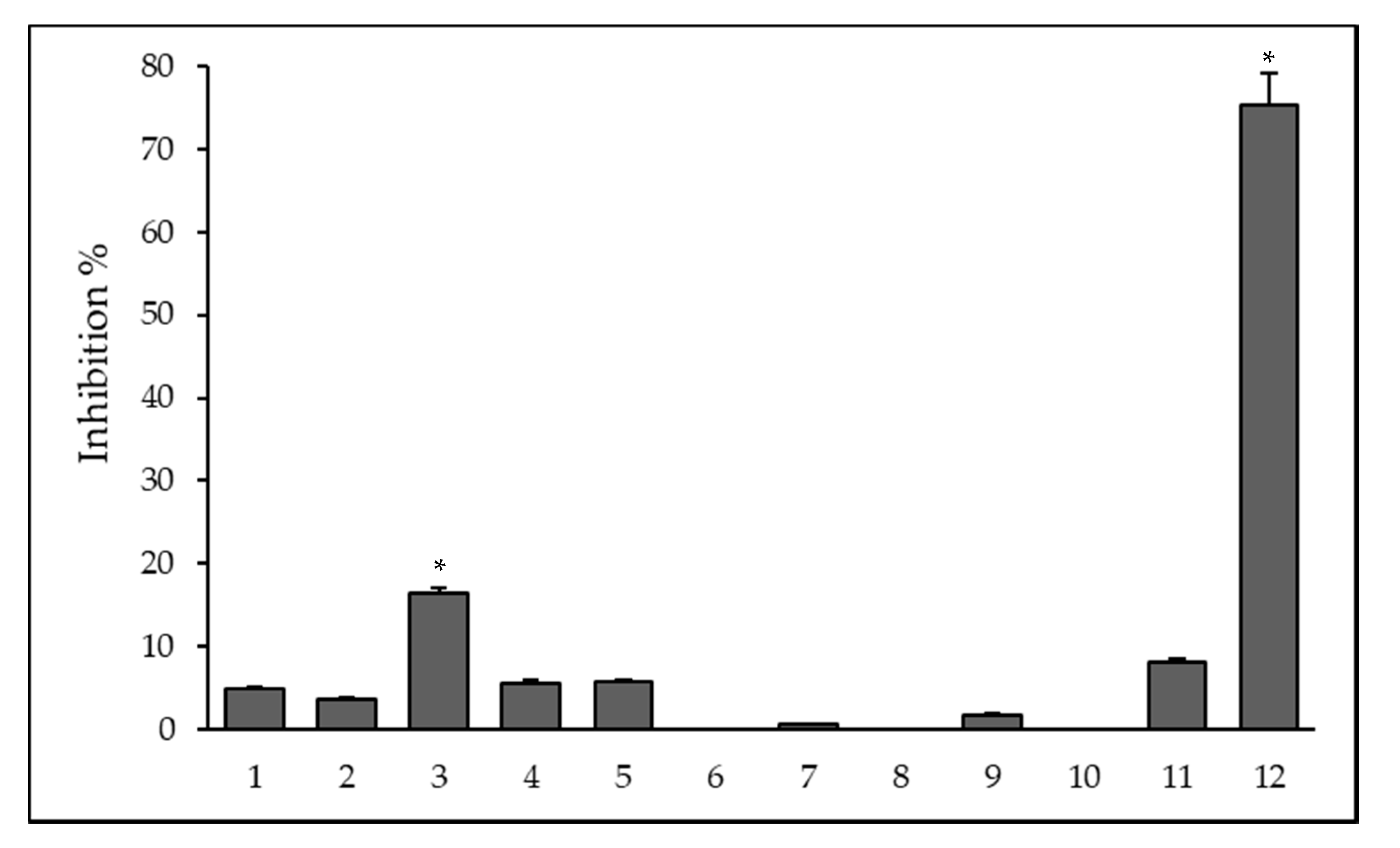
2, respectively; however, the nature of the interaction between QS molecules and benzoxazole-based compounds needs further investigation. The QS-related process is the biofilm formation whereby microorganisms irreversibly attach to and grow on a surface and produce extracellular polymers that facilitate attachment and matrix formation, resulting in increased resistance towards antibiotic treatment. The most pronounced anti-biofilm activity was found in case of the Fe(III) complex
12 of ligand
1 on reference
S. aureus and this complex had no toxicity on normal human fibroblast cells. Regarding antibacterial activity of the metal complexes, there were multiple modes of action. They could undergo ligand exchange reactions, release bioactive molecules, or be triggered by light irradiation to generate reactive oxygen species (ROS). However, complexes
4–
12, except the Ag(I) complex
11, were fairly stable against air and moisture because the ligands
1–
3 can possess a heteroaryl-substituted alkenol ligand system and act as the
N,O-chelating ligand, producing highly stable metal complexes, which could be reinforced by a positive inductive effect of the heterocyclic moiety and a negative inductive effect of the CF
3 group [
18]. For example, Zn(II) complex
6 was inert to excess ethylenediaminetetraacetic acid (EDTA) at room temperature [
18]. Therefore, these metal complexes may be stable in vivo. The increased potency on complexation of ligands with metals can be explained by the enhancement of the penetration into the lipid membranes due to the increased lipophilicity of the complexes [
22,
23].
Based on preliminary results, all compounds except Pd(II) complex
10 showed higher toxicity (IC
50: 0.5–10.99 µM) against MDR mouse T-cell lymphoma cells than chemotherapeutic drug cisplatin (IC
50: 11.20 µM) used as reference compound [
19]. The present data demonstrated that Zn(II) complexes
4,
5,
6, and Ni(II) complex
8 had a stronger inhibitory effect on MRC-5 human embryonic lung fibroblast cell line compared to the activity of cisplatin (IC
50: 33.45 µM) [
24] on the same cell line.
4. Materials and Methods
4.1. Compounds
The synthesis of three 2-trifluoroacetonylbenzoxazole ligands and their metal complexes (Zn, Cu, Ni, Mg, Pd, and Ag) was described previously [
18] (
Table 6). The Fe(III) complex
12 was obtained by the reaction of
1 with FeCl
3 using the same method described for the preparation of
4 [
18]. The compounds (
1–
12) screened for their antibacterial, MDR-modulating, anti-biofilm, and QS inhibition activities, furthermore, they were investigated in combination with antibiotics. The powdered compounds were dissolved in DMSO and stock solutions of 10 mM were prepared. The preparation was performed by vortexing at room temperature.
PMZ, CPZ, crystal violet (CV), EB, VER, TET, CIP, Luria–Bertani (LB) broth, and LB agar were obtained from Sigma–Aldrich Chemie GmbH (Steinheim, Germany). The modified LB medium (LB*) consisted of yeast extract 5 g/L, tryptone 10 g/L, NaCl 10 g/L, K2HPO4 1 g/L, MgSO4 × 7H2O 0.3 g/L, and FeNaEDTA 36 mg/L. To obtain modified LB* agar, the LB* medium was enriched with agar 20 g/L (Difco, Detroit, MI, USA). The pH was adjusted to 7.2. Tryptic soy broth (TSB), tryptic soy agar, and Mueller Hinton broth (MHB) were obtained from Scharlau Chemie S.A. (Barcelona, Spain). CV solution was prepared by adding 0.05 g of CV to 50 mL of distilled water, then the solution was filtered (0.22 μM filter).
4.2. Bacterial Strains
Wild-type E. coli K-12 AG100 strain (argE3 thi-1 rpsL xyl mtl Δ(gal-uvrB) supE44) expressing the AcrAB TolC EP at its basal level and its AcrAB-TolC deleted mutant E. coli K-12 AG100 A strain. The strains were donated by Prof. Dr. Hiroshi Nikaido (Department of Molecular and Cell Biology and Chemistry, University of California, Berkeley, CA, USA).
The compounds were further investigated against two Gram-positive strains, S. aureus ATCC 25923 (MSSA), used as the methicillin susceptible reference strain, and the methicillin- and ofloxacin-resistant S. aureus 272123 clinical isolate (MRSA), which was given by Prof. Dr. Leonard Amaral (Institute of Hygiene and Tropical Medicine, Lisbon, Portugal).
The following bacterial strains were applied in the QS tests: C. violaceum 026 as sensor strain and E. cloacae 31298 isolated from clinical wound (used as N-acyl-homoserine lactone (AHL) producer); this strain induced pigment production by C. violaceum and is a model strain to study QS interactions.
4.3. Cell Culture
The MRC-5 human embryonic lung fibroblast cell line (ATCC CCL-171; provider: LGC Promochem, Teddington, UK) was used in the experiments. The cells were cultivated in Eagle’s Minimal Essential Medium (EMEM; Sigma–Aldrich, St. Louis, MO, USA), supplemented with 1% non-essential amino acid mixture, 10% heat-inactivated fetal bovine serum, 2 mM L-glutamine, 1 mM Na-pyruvate, nystatin, and a penicillin–streptomycin mixture in concentrations of 100 U/L and 10 mg/L, respectively. The supplementary components were obtained from Sigma–Aldrich, St. Louis, MO, USA. The cell culture was kept in a humidified atmosphere (5% CO2, 95% air) at 37 °C.
4.4. Determination of Minimum Inhibitory Concentrations
The minimum inhibitory concentrations (MICs) of compounds were determined in 96-well plates based on the Clinical and Laboratory Standard Institute guidelines (CLSI guidelines) [
25]. The compounds were diluted in 100 μL of MHB. Then, 10
−4 dilution of an overnight bacterial culture in 100 μL of medium were added to each well, with the exception of the medium control wells. The plates were further incubated at 37 °C for 18 h; at the end of the incubation period, the MIC values of tested compounds were determined by visual inspection.
4.5. Combination Effect
The chemosensitizing activity of benzoxazole-type ligands and their metal complexes was determined based on the MIC values of the antibiotics in the presence of sub-inhibitory concentrations of the compounds (¼ MIC) in both Gram-negative and Gram-positive strains. The MICs were determined in strains by the two-fold broth microdilution method in 96-well plates, using serial dilutions of TET and CIP. The first four rows contained two-fold dilutions of antibiotics, and combinations of the TET and CIP and tested compounds were transferred into the last four rows. The 10−4 dilution of an overnight bacterial culture in 50 μL of MHB was then measured to each well, except for the medium control wells. The plates were kept at 37 °C for 18 h. MIC values of antibiotics and their combination with tested compounds were detected by visual inspection.
4.6. Real-Time Accumulation Assay
The accumulation of the efflux pump substrate EB was monitored by the automated EB method [
26] using a LightCycler real-time thermocycler (LightCycler 1.5, Roche, Indianapolis, IN, USA). An aliquot of an overnight culture of the
S. aureus strains in TSB medium was inoculated into fresh TSB medium, and it was incubated until it reached an optical density (OD) of 0.6 at 600 nm. In the case of
E. coli strains, the medium applied in the assay was LB broth; the preparation of the inoculum was similar to the one of
S. aureus. The cells were washed with phosphate buffered saline (PBS; pH 7.4) and centrifuged at 13,000 ×
g for 3 min, the pellets were re-suspended in PBS (pH 7.4), and the OD was calibrated to 0.6 at 600 nm. The compounds were measured at ¼ MIC concentration (in double-concentrated form) to the EB solution in PBS. The final concentration of EB was determined according to the MIC and the fluorescent signal produced by this amount of EB. In the case of all strains, the concentration of EB was 1 μg/mL. Then, 10 μL of the EB solution plus the compound were pipetted into standard glass capillary tubes of 20 µL maximum volume (Roche, Indianapolis, USA), and 10 μL of bacterial culture (OD of 0.6 at 600 nm) were transferred to the capillaries. The capillaries containing the samples were placed into the carousel, and the fluorescence was monitored at the FL-2 channel in every minute on a real-time basis.
Based on the data obtained, the relative fluorescence index (RFI) of the last time point (minute 30) was determined as follows:
where RF
treated is the relative fluorescence (RF) at the last time point of the EB retention curve in the presence of an inhibitor, and RF
untreated is the relative fluorescence at the last time point of the EB retention curve of the untreated control having the solvent control (DMSO). VER was used as a positive control on MRSA strain and CPZ was used on Gram-negative and
S. aureus ATCC strains.
4.7. Anti-Biofilm Effect
The biofilm production of the S. aureus ATCC 25923 strain was studied in 96-well plates using TSB broth in the presence of compounds. The overnight cultures were diluted to an OD of 0.1 at 600 nm and then measured to each well with the exception of the medium control wells, and compounds were pipetted individually at ¼ MIC concentration. The final volume was 200 μL in each well. The samples were incubated at 30 °C for 48 h with gentle agitation (100 rpm). Then, the medium was removed, and the plate was washed with tap water to discard unattached cells. 200 μL CV (0.1% [v/v]) was added to the wells and incubated for 15 min at room temperature. CV was removed from the wells and the plate was washed again with tap water. 200 μL 70% ethanol was transferred to each well and the biofilm formation was determined by measuring the OD at 600 nm using Multiscan EX ELISA reader (Thermo Labsystems, Cheshire, CT, USA). The anti-biofilm effect of compounds was determined as the percentage (%) of decrease in biofilm formation. The assay was repeated a minimum of three times. The results were calculated using a t-test and p-values of <0.001 were considered significant.
4.8. Assay for Quorum Sensing Inhibition
LB* was applied for these experiments. The sensor strain
C. violaceum 026 and the AHL producer strains
E. cloacae 31298 were inoculated as parallel lines and incubated at room temperature for 24–48 h. The inhibition of bacterial communication was determined by the agar diffusion method. Filter paper discs (7.0 mm in diameter) were impregnated with 10 μL of stock solutions (1 mM) of the compounds in DMSO. The discs were placed between the parallel lines of the sensor and the AHL producer strains. The plates were kept at room temperature for another 24–48 h, and the interactions between the strains and compounds were determined as the reduction in the size of the zone of purple pigment (violacein) production in millimeters [
27]. PMZ was applied as a positive control.
4.9. Assay for Cytotoxic Effect
The activity of increasing concentrations of the compounds on cell growth was determined in 96-well flat-bottomed microtiter plates by two-fold serial dilutions of the benzoxazole derivatives. The adherent human embryonic lung fibroblast cells (10
4/well) was seeded in EMEM medium in 96-well microtiter plates for 4 h prior to the assay. The serial dilutions of the compounds were made in a separate plate, and then pipetted to the plates containing the MRC-5 cell line. Culture plates were incubated at 37 °C for 24 h; at the end of the incubation period, 20 μL of MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) solution (from a 5 mg/mL stock solution) was measured to each well. After the incubation at 37 °C for 4 h, 100 μL of sodium dodecyl sulfate (SDS) solution (10% SDS in 0.01 M HCl) was added to each well to dissolve the formazan crystals and the samples were further incubated at 37 °C overnight. Cell growth was calculated by measuring the OD at 540 nm (ref. 630 nm) with a Multiscan EX ELISA reader. The cytotoxicity was expressed as IC
50 values, defined as the inhibitory dose that reduces the growth of the cells exposed to the tested compounds by 50%. IC
50 values and the standard deviation (SD) of triplicate experiments were calculated by using GraphPad Prism software version 5.00 for Windows with nonlinear regression curve fit (GraphPad Software, San Diego, CA, USA;
www.graphpad.com) where IC
50: 0–30, IC
50: 31–60, IC
50: 61–100, and IC
50 > 100 µM represent strong, moderate, weak, and no cytotoxic effects, respectively.
4.10. Statistical Analysis
The values were given as the mean ± SD determined for three replicates from three independent experiments. The analysis of data was performed using SigmaPlot for Windows Version 12.0 software (Systat Software Inc, San Jose, CA, USA), applying the two-tailed t-test.

 ,
, 


{kind=link}
