Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment


Abstract
:1. Introduction
2. Carbapenemases-Producing Enterobacteriaceae from Humans
3. Companion Animals
4. Carbapenemases-Producing Enterobacteriaceae in Other Animals
5. Food Producing Animals
6. Carbapenemases Producing Enterobacteriaceae in Food
7. Carbapenemases-Producing Enterobacteriaceae from the Environment
8. Molecular Epidemiology of Carbapenem Resistance (CR) Genes
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. World Health Organization. Gloval Priority List of Antiviotic-Resistand Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Naas, T.; Poirel, L. Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M. Colistin and tigecycline resistance in Carbapenem-resistant Enterobacteriaceae: Checkmate to our last line of defense. Infect. Control. Hosp. Epidemiol. 2016, 37, 624–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu, C.-C.; Chang, Y.-T.; Lin, S.-Y.; Chen, Y.-H.; Hsueh, P.-R. Infections caused by Carbapenem-resistant Enterobacteriaceae: An update on therapeutic options. Front. Microbiol. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P. Pharmacology of specific drug groups. In Pharmacology and Therapeutics for Dentistry; Dowd, F.J., Johnson, B.S., Mariotti, A.J., Eds.; Elsevier: Milton, ON, Canada, 2017; pp. 457–487. [Google Scholar]
- Djahmi, N.; Dunyach-Rémy, C.; Pantel, A.; Dekhil, M.; Sotto, A.; Lavigne, J.-P. Epidemiology of Carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.E.V.; Boerlin, P. Carbapenemase-producing Enterobacteriaceae in animals and methodologies for their detection. Can. J. Vet. Res. 2020, 84, 3–17. [Google Scholar]
- Cheng, Z.; Thomas, P.W.; Ju, L.; Bergstrom, A.; Mason, K.; Clayton, D.; Miller, C.; Bethel, C.R.; Vanpelt, J.; Tierney, D.L.; et al. Evolution of New Delhi metallo-β-lactamase (NDM) in the clinic: Effects of NDM mutations on stability, zinc affinity, and mono-zinc activity. J. Biol. Chem. 2018, 293, 12606–12618. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Nordmann, P. Analysis of a Carbapenem-hydrolyzing class A beta-lactamase from Enterobacter cloacae and of its LysR-type regulatory protein. Proc. Natl. Acad. Sci. USA 1994, 91, 7693–7697. [Google Scholar] [CrossRef] [Green Version]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Jacoby, G.A. Updated functional classification of β-Lactamases. Antimicrob. Agents Chemother. 2009, 54, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, S.E.; Livermore, D.M.; Hooper, D.C.; Hope, W.W. Metallo-β-lactamases: Structure, function, epidemiology, treatment options, and the development pipeline. Antimicrob. Agents Chemother. 2020, 10, e00397-20. [Google Scholar] [CrossRef] [PubMed]
- Mathers, A.J.; Stoesser, N.; Chai, W.; Carroll, J.; Barry, K.; Cherunvanky, A.; Sebra, R.; Kasarskis, A.; Peto, T.E.; Walker, A.S.; et al. Chromosomal integration of the Klebsiella pneumoniae Carbapenemase gene, blaKPC, in Klebsiella species is elusive but not rare. Antimicrob. Agents Chemother. 2016, 61, e01823-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botelho, J.; Roberts, A.P.; León-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the move: It’s good to be on ICEs. Mob. DNA 2018, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souli, M.; Galani, I.; Giamarellou, H. Emergence of extensively drug-resistant and pandrug-resistant Gram-negative bacilli in Europe. Euro Surveill. Eur. Commun. Dis. Bull. 2008, 13, 19045. [Google Scholar]
- Evans, B.A.; Amyes, S. OXA-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem resistance: A review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Moxon, C.A.; Paulus, S. Beta-lactamases in Enterobacteriaceae infections in children. J. Infect. 2016, 72, S41–S49. [Google Scholar] [CrossRef] [Green Version]
- Yigit, H.; Queenan, A.M.; Anderson, G.J.; Domenech-Sanchez, A.; Biddle, J.W.; Steward, C.D.; Alberti, S.; Bush, K.; Tenover, F.C. Novel Carbapenem-hydrolyzing β-Lactamase, KPC-1, from a Carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Drew, R.J.; Turton, J.; Hill, R.; Livermore, D.; Woodford, N.; Paulus, S.; Cunliffe, N.A. Emergence of Carbapenem-resistant Enterobacteriaceae in a UK paediatric hospital. J. Hosp. Infect. 2013, 84, 300–304. [Google Scholar] [CrossRef]
- Livermore, D.M.; Woodford, N. The β-lactamase threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol. 2006, 14, 413–420. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Risk Assessment on the Spread of Carbapenemase-Producing Enterobacteriaceae (CPE) through Patient Transfer between Healthcare Facilities, with Special Emphasis on Cross-Border Transfer; ECDC Technical Report; ECDC: Stockholm, Sweden, 2011. [Google Scholar]
- Azap, O.; Otlu, B.; Yesilkaya, A.; Yakupoğulları, Y. Detection of OXA-48-like Carbapenemase-producing Klebsiella pneumoniae in a tertiary care center in Turkey: Molecular characterization and epidemiology. Balk. Med. J. 2013, 30, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Hooper, D.C. Hospital-acquired infections due to gram-negative bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Ahn, M.Y.; Chung, J.-W.; Lee, M.-K. In vitro antibacterial activity of Doripenem against gram-negative blood isolates in a Korean tertiary care center. Infect. Chemother. 2015, 47, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Aschbacher, R.; Pagani, L.; Doumith, M.; Pike, R.; Woodford, N.; Spoladore, G.; Larcher, C.; Livermore, D.M. Metallo-β-lactamases among Enterobacteriaceae from routine samples in an Italian tertiary care hospital and long-term care facilities during 2008. Clin. Microbiol. Infect. 2010, 17, 181–189. [Google Scholar] [CrossRef]
- Ahn, C.; Syed, A.; Hu, F.; O’Hara, J.A.; Rivera, J.I.; Doi, Y. Microbiological features of KPC-producing Enterobacter isolates identified in a U.S. hospital system. Diagn. Microbiol. Infect. Dis. 2014, 80, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Chopra, T.; Rivard, C.; Awali, R.A.; Krishna, A.; Bonomo, R.A.; Perez, F.; Kaye, K.S. Epidemiology of Carbapenem-resistant Enterobacteriaceae at a long-term acute care hospital. Open Forum Infect. Dis. 2018, 5, 224. [Google Scholar] [CrossRef]
- Tamma, P.D.; Kazmi, A.; Bergman, Y.; Goodman, K.E.; Ekunseitan, E.; Amoah, J.; Simner, P.J. The likelihood of developing a Carbapenem-resistant infection during the hospital stay. Antimicrob Agents Chemother 2019. [Google Scholar] [CrossRef] [Green Version]
- Senchyna, F.; Gaur, R.L.; Sandlund, J.; Truong, C.; Tremintin, G.; Kültz, D.; Gomez, C.A.; Tamburini, F.B.; Andermann, T.M.; Bhatt, A.; et al. Diversity of resistance mechanisms in Carbapenem-resistant Enterobacteriaceae at a health care system in Northern California, from 2013 to 2016. Diagn. Microbiol. Infect. Dis. 2019, 93, 250–257. [Google Scholar] [CrossRef]
- Kohler, P.P.; Melano, R.G.; Patel, S.N.; Shafinaz, S.; Faheem, A.; Coleman, B.L.; Green, K.; Armstrong, I.; Almohri, H.; Borgia, S.; et al. Emergence of Carbapenemase-producing Enterobacteriaceae, South-Central Ontario, Canada1. Emerg. Infect. Dis. 2018, 24, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- Souli, M.; Galani, I.; Antoniadou, A.; Papadomichelakis, E.; Poulakou, G.; Panagea, T.; Vourli, S.; Zerva, L.; Armaganidis, A.; Kanellakopoulou, K.; et al. An outbreak of infection due to β-Lactamase Klebsiella pneumoniae Carbapenemase 2–producing K. pneumoniae in a Greek University Hospital: Molecular characterization, epidemiology, and outcomes. Clin. Infect. Dis. 2010, 50, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giakkoupi, P.; Papagiannitsis, C.C.; Miriagou, V.; Pappa, O.; Polemis, M.; Tryfinopoulou, K.; Tzouvelekis, L.S.; Vatopoulos, A. An update of the evolving epidemic of blaKPC-2-carrying Klebsiella pneumoniae in Greece (2009–10). J. Antimicrob. Chemother. 2011, 66, 1510–1513. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, H.; Livermore, D.M.; Giske, C.G.; Rossolini, G.M.; Campos, J.; Vatopoulos, A.; Gniadkowski, M.; Toth, A.; Pfeifer, Y.; Jarlier, V.; et al. Carbapenem-non-susceptible Enterobacteriaceae in Europe: Conclusions from a meeting of national experts. Eurosurveillance 2010, 15, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonne, A.; Thiolet, J.M.; Fournier, S.; Fortineau, N.; Kassis-Chikhani, N.; Boytchev, I.; Aggoune, M.; Séguier, J.C.; Sénéchal, H.; Tavolacci, M.P.; et al. Control of a multi-hospital outbreak of KPC-producing Klebsiella pneumoniae type 2 in France, September to October 2009. Eurosurveillance 2010, 15, 19734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteo, J.; Saez, D.; Bautista, V.; Fernández-Romero, S.; Hernández-Molina, J.M.; Pérez-Vázquez, M.; Aracil, B.; Campos, J. Carbapenemase-producing Enterobacteriaceae in Spain in 2012. Antimicrob. Agents Chemother. 2013, 57, 6344–6347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Cuzon, G.; Naas, T. The real threat of Klebsiella pneumoniae Carbapenemase-producing bacteria. Lancet Infect. Dis. 2009, 9, 228–236. [Google Scholar] [CrossRef]
- Ducomble, T.; Faucheux, S.; Helbig, U.; Kaisers, U.; Konig, B.; Knaust, A.; Lübbert, C.; Möller, I.; Rodloff, A.; Schweickert, B.; et al. Large hospital outbreak of KPC-2-producing Klebsiella pneumoniae: Investigating mortality and the impact of screening for KPC-2 with polymerase chain reaction. J. Hosp. Infect. 2015, 89, 179–185. [Google Scholar] [CrossRef]
- Raro, O.H.F.; Da Silva, R.M.C.; Filho, E.M.R.; Sukiennik, T.C.T.; Stadnik, C.; Dias, C.A.G.; Iglesias, J.O.; Pérez-Vázquez, M. Carbapenemase-producing Klebsiella pneumoniae from transplanted patients in Brazil: Phylogeny, resistome, virulome and mobile genetic elements harboring blaKPC-2 or blaNDM-1. Front. Microbiol. 2020, 11, 1563. [Google Scholar] [CrossRef]
- Manenzhe, R.I.; Zar, H.J.; Nicol, M.P.; Kaba, M. The spread of Carbapenemase-producing bacteria in Africa: A systematic review. J. Antimicrob. Chemother. 2014, 70, 23–40. [Google Scholar] [CrossRef] [Green Version]
- Singh-Moodley, A.; Perovic, O. Antimicrobial susceptibility testing in predicting the presence of carbapenemase genes in Enterobacteriaceae in South Africa. BMC Infect. Dis. 2016, 16, 536. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Pitout, J.D.; Nordmann, P. Carbapenemases: Molecular diversity and clinical consequences. Futur. Microbiol. 2007, 2, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Genetic features of the widespread plasmid coding for the Carbapenemase OXA-48. Antimicrob. Agents Chemother. 2011, 56, 559–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, H.S. Global dissemination of Carbapenemase-producing Klebsiella pneumoniae: Epidemiology, genetic context, treatment options, and detection methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, C.; Coope, C.; Conway, L.; Higgins, J.P.; McCulloch, J.; Okoli, G.; Patel, B.; Oliver, I. Control of Carbapenemase-producing Enterobacteriaceae outbreaks in acute settings: An evidence review. J. Hosp. Infect. 2017, 95, 3–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senchyna, F.; Gaur, R.; Sandlund, J.; Truong, C.; Tremintin, G.; Küeltz, D.; Gomez, C.A.; Tamburini, F.B.; Andermann, T.M.; Bhatt, A.S.; et al. Diverse mechanisms of resistance in Carbapenem-resistant Enterobacteriaceae at a health care system in Silicon Valley, California. bioRxiv 2018. [Google Scholar] [CrossRef]
- Guo, X.; Rao, Y.; Guo, L.; Xu, H.; Lv, T.; Yu, X.; Chen, Y.; Liu, N.; Han, H.; Zheng, B. Detection and genomic characterization of a Morganella morganii isolate from China that produces NDM-5. Front. Microbiol. 2019, 10, 1156. [Google Scholar] [CrossRef] [PubMed]
- Cuzon, G.; Demachy, M.C.; Nordmann, P.; Naas, T. Plasmid-mediated Carbapenem-hydrolyzing β-Lactamase KPC-2 in Klebsiella pneumoniae isolate from Greece. Antimicrob. Agents Chemother. 2007, 52, 796–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, T.; Vedel, G.; Nordmann, P.; Poyart, C. Plasmid-mediated Carbapenem-hydrolyzing β-Lactamase KPC in a Klebsiella pneumoniae isolate from France. Antimicrob. Agents Chemother. 2005, 49, 4423–4424. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Nahid, F.; Khan, A.A.; Rehman, S.; Zahra, R. Prevalence of metallo-β-lactamase NDM-1-producing multi-drug resistant bacteria at two Pakistani hospitals and implications for public health. J. Infect. Public Heal. 2013, 6, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Van Der Bij, A.K.; Pitout, J.D.D. The role of international travel in the worldwide spread of multiresistant Enterobacteriaceae. J. Antimicrob. Chemother. 2012, 67, 2090–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tijet, N.; Alexander, D.C.; Richardson, D.; Lastovetska, O.; Low, N.E.; Patel, S.N.; Melano, R.G. New Delhi Metallo-β-Lactamase, Ontario, Canada. Emerg. Infect. Dis. 2011, 17, 306–307. [Google Scholar] [CrossRef] [PubMed]
- Bernick, J.; Beliavsky, A.; Bogoch, I.I. Endometritis and bacteremia with a New Delhi Metallo-Beta-Lactamase 1 (NDM-1)-containing organism in a remote traveler. J. Obstet. Gynaecol. Can. 2019, 41, 753–754. [Google Scholar] [CrossRef] [PubMed]
- Mataseje, L.F.; Boyd, D.A.; Mulvey, M.R.; Longtin, Y. Report on two hypervirulent Klebsiella pneumoniae producing a blaKPC-2 Carbapenemase from a Canadian patient. Antimicrob Agents Chemother 2019. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Vuillemin, X.; Juhas, M.; Masseron, A.; Bechtel-Grosch, U.; Tiziani, S.; Mancini, S.; Nordmann, P. KPC-50 confers resistance to Ceftazidime-Avibactam associated with reduced Carbapenemase activity. Antimicrob. Agents Chemother. 2020, 64, e00321-20. [Google Scholar] [CrossRef]
- López, J.; Correa, A.; Navon-Venezia, S.; Torres, J.; Briceno, D.; Montealegre, M.C.; Quinn, J.; Carmeli, Y.; Villegasz, M. Intercontinental spread from Israel to Colombia of a KPC-3-producing klebsiella pneumoniae strain. Clin. Microbiol. Infect. 2011, 17, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Adler, A.; Shklyar, M.; Schwaber, M.J.; Navon-Venezia, S.; Dhaher, Y.; Edgar, R.; Solter, E.; Benenson, S.; Masarwa, S.; Carmeli, Y. Introduction of OXA-48-producing Enterobacteriaceae to Israeli hospitals by medical tourism. J. Antimicrob. Chemother. 2011, 66, 2763–2766. [Google Scholar] [CrossRef] [Green Version]
- ECDC. European Centre for Disease Prevention and Control. Outbreak of VIM-Producing Carbapenem Resistant Pseudomonas aeruginosa Linked to medical Tourism to Mexico; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Cheng, V.C.; Wong, S.-C.; Wong, S.C.; Ho, P.-L.; Yuen, K.-Y. Control of Carbapenemase-producing Enterobacteriaceae: Beyond the hospital. EClinicalMedicine 2019, 6, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Gentilini, F.; Turba, M.E.; Pasquali, F.; Mion, D.; Romagnoli, N.; Zambon, E.; Terni, D.; Peirano, G.; Pitout, J.D.D.; Parisi, A.; et al. Hospitalized pets as a source of Carbapenem-resistance. Front. Microbiol. 2018, 9, 2872. [Google Scholar] [CrossRef]
- Hong, J.S.; Song, W.; Park, H.-M.; Oh, J.-Y.; Chae, J.-C.; Han, J.-I.; Jeong, S.H. First detection of New Delhi Metallo-β-Lactamase-5-producing Escherichia coli from companion animals in Korea. Microb. Drug Resist. 2019, 25, 344–349. [Google Scholar] [CrossRef]
- Adams, R.J.; Kim, S.S.; Mollenkopf, D.F.; Mathys, D.A.; Schuenemann, G.M.; Daniels, J.B.; Wittum, T. Antimicrobial-resistant Enterobacteriaceae recovered from companion animal and livestock environments. Zoonoses Public Health 2018, 65, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Stolle, I.; Prenger-Berninghoff, E.; Stamm, I.; Scheufen, S.; Hassdenteufel, E.; Guenther, S.; Bethe, A.; Pfeifer, Y.; Ewers, C. Emergence of OXA-48 Carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J. Antimicrob. Chemother. 2013, 68, 2802–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönthal, T.; Österblad, M.; Eklund, M.; Jalava, J.; Nykäsenoja, S.; Pekkanen, K.; Rantala, M. Sharing more than friendship-transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between dogs and humans in a family, Finland, 2015. Eurosurveillance 2018, 23, 1700497. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Brouwers, H.J.M.; Cobbold, R.N.; Platell, J.L.; Chapman, T.A.; Barrs, V.R.; Johnson, J.R.; Trott, D.J. Fluoroquinolone-resistant extraintestinal pathogenic Escherichia coli, including O25b-ST131, isolated from faeces of hospitalized dogs in an Australian veterinary referral centre. J. Antimicrob. Chemother. 2013, 68, 1025–1031. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Clabots, C.; Kuskowski, M.A. Multiple-host sharing, long-term persistence, and virulence of Escherichia coli clones from human and animal household members. J. Clin. Microbiol. 2008, 46, 4078–4082. [Google Scholar] [CrossRef] [Green Version]
- Nicolas-Chanoine, M.-H.; Bertrand, X.; Madec, J.-Y. Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.; O’Dea, M.; Trott, D.J.; Abraham, R.J.; Hughes, D.; Pang, S.; McKew, G.; Cheong, E.Y.L.; Merlino, J.; Saputra, S.; et al. Isolation and plasmid characterization of Carbapenemase (IMP-4) producing Salmonella enterica Typhimurium from cats. Sci. Rep. 2016, 6, 35527. [Google Scholar] [CrossRef]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: A systematic review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, X.; Schwarz, S.; Zhang, R.; Lei, L.; Liu, X.; Lin, D.; Shen, J. IMP-45-producing multidrug-resistant Pseudomonas aeruginosa of canine origin. J. Antimicrob. Chemother. 2014, 69, 2579–2581. [Google Scholar] [CrossRef] [Green Version]
- Rubin, J.; Pitout, J.D. Extended-spectrum β-lactamase, Carbapenemase and AmpC producing Enterobacteriaceae in companion animals. Veter Microbiol. 2014, 170, 10–18. [Google Scholar] [CrossRef]
- Reynolds, M.E.; Phan, H.T.T.; George, S.; Hubbard, A.; Stoesser, N.; Maciuca, E.I.; Crook, D.W.; Timofte, D. Occurrence and characterization of Escherichia coli ST410 co-harbouring blaNDM-5, blaCMY-42 and blaTEM-190 in a dog from the UK. J. Antimicrob. Chemother. 2019, 74, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.S.; Kuster, S.P.; Nigg, A.; Dazio, V.; Brilhante, M.; Rohrbach, H.; Bernasconi, O.J.; Büdel, T.; Campos-Madueno, E.I.; Brawand, S.G.; et al. Poor infection prevention and control standards are associated with environmental contamination with Carbapenemase-producing Enterobacterales and other multidrug-resistant bacteria in Swiss companion animal clinics. Antimicrob. Resist. Infect. Control. 2020, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.; Morton, J.; Cobbold, R.; Sidjabat, H.; Filippich, L.; Trott, D. Multidrug-resistant E. coli and Enterobacter extraintestinal infection in 37 dogs. J. Veter. Intern. Med. 2008, 22, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Rodríguez, I.; Schmoger, S.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Escherichia coli producing VIM-1 Carbapenemase isolated on a pig farm. J. Antimicrob. Chemother. 2012, 67, 1793–1795. [Google Scholar] [CrossRef]
- Poirel, L.; Berçot, B.; Millemann, Y.; Bonnin, R.A.; Pannaux, G.; Nordmann, P. Carbapenemase-producing Acinetobacter spp. in Cattle, France. Emerg. Infect. Dis. 2012, 18, 523–525. [Google Scholar] [CrossRef]
- Fischer, J.; Rodríguez, I.; Schmoger, S.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Salmonella enterica subsp. enterica producing VIM-1 Carbapenemase isolated from livestock farms. J. Antimicrob. Chemother. 2012, 68, 478–480. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; José, M.S.; Roschanski, N.; Schmoger, S.; Baumann, B.; Irrgang, A.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Spread and persistence of VIM-1 Carbapenemase-producing Enterobacteriaceae in three German swine farms in 2011 and 2012. Veter. Microbiol. 2017, 200, 118–123. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Zhang, Q.; Qi, J.; Liu, H.; Wang, Y.; He, T.; Ma, L.; Lai, J.; Shen, Z.; et al. Identification of New Delhi Metallo-β-lactamase 1 in Acinetobacter lwoffii of food animal origin. PLoS ONE 2012, 7, e37152. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Wei, R.; Zhang, L.; Sun, L.; Pang, M.; Wang, R.; Wang, Y. Characterization of NDM-5-positive extensively resistant Escherichia coli isolates from dairy cows. Veter. Microbiol. 2017, 207, 153–158. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Wang, X.; Liu, D.; Ke, Y.; Wang, Y.; Shen, J. Novel variant of New Delhi Metallo-β-lactamase, NDM-20, in Escherichia coli. Front. Microbiol. 2018, 9, 248. [Google Scholar] [CrossRef] [Green Version]
- Morrison, B.J.; Rubin, J. Carbapenemase producing bacteria in the food supply escaping detection. PLoS ONE 2015, 10, e0126717. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, H.; Reuland, E.A.; Wintermans, B.B.; Al Naiemi, N.; Koek, A.; Abdelwahab, A.M.; Ammar, A.M.; Mohamed, A.A.; Vandenbroucke-Grauls, C.M.J.E. Extended-spectrum β-Lactamases and/or Carbapenemases-producing Enterobacteriaceae isolated from retail chicken meat in Zagazig, Egypt. PLoS ONE 2015, 10, e0136052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, K.; Khattak, F.; Ali, A.; Rahat, S.; Noor, S.; Mahsood, N.; Somayya, R. Carbapenemases and extended-spectrum β-Lactamase-producing multidrug-resistant Escherichia coli isolated from retail chicken in Peshawar: First report from Pakistan. J. Food Prot. 2018, 81, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.-H. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef] [Green Version]
- VIM-1-producing Salmonella infantis isolated from swine and minced pork meat in Germany. J. Antimicrob. Chemother. 2017, 72, 2131–2133. [CrossRef] [Green Version]
- Wang, W.; Baloch, Z.; Peng, Z.; Hu, Y.; Xu, J.; Fanning, S.; Li, F. Genomic characterization of a large plasmid containing a bla NDM-1 gene carried on Salmonella enterica serovar Indiana C629 isolate from China. BMC Infect. Dis. 2017, 17, 479. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lv, L.; Huang, X.; Huang, Y.; Zhuang, Z.; Lu, J.; Liu, E.; Wan, M.; Xun, H.; Zhang, Z.; et al. Rapid increase in Carbapenemase-producing Enterobacteriaceae in retail meat driven by the spread of the blaNDM-5-carrying IncX3 plasmid in China from 2016 to 2018. Antimicrob. Agents Chemother. 2019, 63, e00573-19. [Google Scholar] [CrossRef] [Green Version]
- Roschanski, N.; Guenther, S.; Vu, T.T.T.; Fischer, J.; Semmler, T.; Huehn, S.; Alter, T.; Roesler, U. VIM-1 Carbapenemase-producing Escherichia coli isolated from retail seafood, Germany 2016. Eurosurveillance 2017, 22, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Janecko, N.; Martz, S.-L.; Avery, B.P.; Daignault, D.; Desruisseau, A.; Boyd, D.; Irwin, R.J.; Mulvey, M.R.; Reid-Smith, R.J. Carbapenem-resistant Enterobacter spp. in retail seafood imported from Southeast Asia to Canada. Emerg. Infect. Dis. 2016, 22, 1675–1677. [Google Scholar] [CrossRef]
- Brouwer, M.S.; Tehrani, K.H.M.E.; Rapallini, M.; Geurts, Y.; Kant, A.; Harders, F.; Mashayekhi, V.; Martin, N.I.; Bossers, A.; Mevius, D.J.; et al. Novel Carbapenemases FLC-1 and IMI-2 encoded by an Enterobacter cloacae complex isolated from food products. Antimicrob. Agents Chemother. 2019, 63, e02338-18. [Google Scholar] [CrossRef] [Green Version]
- Diab, M.; Hamze, M.; Bonnet, R.; Saras, E.; Madec, J.-Y.; Haenni, M. OXA-48 and CTX-M-15 extended-spectrum beta-lactamases in raw milk in Lebanon: Epidemic spread of dominant Klebsiella pneumoniae clones. J. Med. Microbiol. 2017, 66, 1688–1691. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yao, X.; Luo, J.; Lv, L.; Zeng, Z.; Liu, J.-H. Emergence of Escherichia coli co-producing NDM-1 and KPC-2 Carbapenemases from a retail vegetable, China. J. Antimicrob. Chemother. 2017, 73, 252–254. [Google Scholar] [CrossRef]
- Touati, A.; Mairi, A.; Baloul, Y.; Lalaoui, R.; Bakour, S.; Thighilt, L.; Gharout, A.; Rolain, J.-M. First detection of Klebsiella pneumoniae producing OXA-48 in fresh vegetables from Béjaïa city, Algeria. J. Glob. Antimicrob. Resist. 2017, 9, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Poirel, L.; Nordmann, P.; Klumpp, J.; Stephan, R. First detection of Klebsiella variicola producing OXA-181 Carbapenemase in fresh vegetable imported from Asia to Switzerland. Antimicrob. Resist. Infect. Control. 2015, 4, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, L.Y.; Apisarnthanarak, A.; Khan, E.; Suwantarat, N.; Ghafur, A.; Tambyah, P.A. Carbapenem-resistant Acinetobacter baumannii and Enterobacteriaceae in South and Southeast Asia. Clin. Microbiol. Rev. 2016, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Lerner, A.; Adler, A.; Abu-Hanna, J.; Meitus, I.; Navon-Venezia, S.; Carmeli, Y. Environmental contamination by Carbapenem-resistant Enterobacteriaceae. J. Clin. Microbiol. 2012, 51, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.K.; Yang, Y.; Tan, B.H. Increased mortality among Carbapenemase-producing Carbapenem-resistant Enterobacteriaceae carriers who developed clinical isolates of another genotype. Open Forum Infect. Dis. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Weingarten, R.A.; Johnson, R.C.; Conlan, S.; Ramsburg, A.M.; Dekker, J.P.; Lau, A.F.; Khil, P.; Odom, R.T.; Deming, C.; Park, M.; et al. Genomic analysis of hospital plumbing reveals diverse reservoir of bacterial plasmids conferring Carbapenem resistance. mBio 2018, 9, 02011–02017. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.U.; Parvez, S. Detection of bla NDM-4 in Escherichia coli from hospital sewage. J. Med. Microbiol. 2014, 63, 1404–1406. [Google Scholar] [CrossRef]
- Zurfluh, K.; Hächler, H.; Nüesch-Inderbinen, M.; Stephan, R. Characteristics of extended-spectrum β-Lactamase-and Carbapenemase-producing Enterobacteriaceae isolates from rivers and lakes in Switzerland. Appl. Environ. Microbiol. 2013, 79, 3021–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.A.; Hellmark, B.; Ehricht, R.; Söderquist, B.; Jass, J. Related Carbapenemase-producing Klebsiella isolates detected in both a hospital and associated aquatic environment in Sweden. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 2241–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piedra-Carrasco, N.; Fàbrega, A.; Calero-Cáceres, W.; Cornejo-Sánchez, T.; Brown-Jaque, M.; Mir-Cros, A.; Muniesa, M.; González-López, J.J. Carbapenemase-producing Enterobacteriaceae recovered from a Spanish river ecosystem. PLoS ONE 2017, 12, e0175246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelle, J.; Johnson, J.R.; Johnston, B.D.; Kinkle, B.; Boczek, L.; Ryu, H.; Hayes, S. Survey of US wastewater for Carbapenem-resistant Enterobacteriaceae. J. Water Health 2019, 17, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Mollenkopf, D.F.; Stull, J.W.; Mathys, D.A.; Bowman, A.S.; Feicht, S.M.; Grooters, S.V.; Daniels, J.B.; Wittum, T. Carbapenemase-Producing Enterobacteriaceae Recovered from the Environment of a Swine Farrow-to-finish operation in the United States. Antimicrob. Agents Chemother. 2016, 61, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vikram, A.; Schmidt, J.W. Functional blaKPC-2 sequences are present in U.S. beef cattle feces regardless of antibiotic use. Foodborne Pathog. Dis. 2018, 15, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Hrenovic, J.; Music, M.S.; Durn, G.; Dekic, S.; Hunjak, B.; Kisic, I. Carbapenem-resistant Acinetobacter baumannii recovered from swine manure. Microb. Drug Resist. 2019, 25, 725–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Chang, J.; Cao, L.; Luo, Q.-X.; Xu, H.; Lyu, W.; Qian, M.; Ji, X.; Zhang, Q.; Xia, X.; et al. Characterization of an NDM-5 Carbapenemase-producing Escherichia coli ST156 isolate from a poultry farm in Zhejiang, China. BMC Microbiol. 2019, 19, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adegoke, A.A.; Fatunla, O.K.; Okoh, A.I. Critical threat associated with carbapenem-resistant gram-negative bacteria: Prioritizing water matrices in addressing total antibiotic resistance. Ann. Microbiol. 2020, 70, 1–13. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Schill, S.; Stoeger, A.; Pekard-Amenitsch, S.; Huhulescu, S.; Inreiter, N.; Hartl, R.; Kerschner, H.; Sorschag, S.; Springer, B.; et al. Whole genome sequencing reveals resemblance between ESBL-producing and carbapenem resistant Klebsiella pneumoniae isolates from Austrian rivers and clinical isolates from hospitals. Sci. Total. Environ. 2019, 662, 227–235. [Google Scholar] [CrossRef]
- Tanner, W.D.; Vanderslice, J.A.; Goel, R.K.; Leecaster, M.K.; Fisher, M.A.; Olstadt, J.; Gurley, C.M.; Morris, A.G.; Seely, K.A.; Chapman, L.; et al. Multi-state study of Enterobacteriaceae harboring extended-spectrum beta-lactamase and Carbapenemase genes in U.S. drinking water. Sci. Rep. 2019, 9, 3938. [Google Scholar] [CrossRef] [Green Version]
- Hamza, D.; Dorgham, S.; Ismael, E.; El-Moez, S.I.A.; ElHariri, M.; Elhelw, R.; Hamza, E. Emergence of β-lactamase-and Carbapenemase-producing Enterobacteriaceae at integrated fish farms. Antimicrob. Resist. Infect. Control. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Bonomo, R.A.; Burd, E.M.; Conly, J.; Limbago, B.M.; Poirel, L.; Segre, J.A.; Westblade, L.F. Carbapenemase-producing organisms: A global scourge. Clin. Infect. Dis. 2017, 66, 1290–1297. [Google Scholar] [CrossRef]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new Metallo-β-Lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33, 33. [Google Scholar] [CrossRef] [PubMed]
- Pournaras, S.; Tsakris, A.; Maniati, M.; Tzouvelekis, L.S.; Maniatis, A.N. Novel variant (blaVIM-4) of the Metallo-β-Lactamase gene blaVIM-1 in a clinical strain of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2002, 46, 4026–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Peirano, G.; Motyl, M.R.; Adams, M.D.; Chen, L.; Kreiswirth, B.; DeVinney, R.; Pitout, J.D.D. Global molecular epidemiology of IMP-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e02729-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Hu, Y.; Sun, Q.; Hu, F.; Zhou, H.; Shu, L.; Ma, T.; Shen, Y.; Wang, Y.; Li, J.; et al. Emerging carriage of NDM-5 and MCR-1 in Escherichia coli from healthy people in multiple regions in China: A cross sectional observational study. EClinicalMedicine 2018, 6, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Bi, Z.; Ma, S.; Chen, B.; Cai, C.; He, J.; Schwarz, S.; Sun, C.; Zhou, Y.; Yin, J.; et al. Inter-host transmission of Carbapenemase-producing Escherichia coli among humans and backyard animals. Environ. Health Perspect. 2019, 127, 107009. [Google Scholar] [CrossRef] [Green Version]
- Zhai, R.; Fu, B.; Shi, X.; Sun, C.; Liu, Z.; Wang, S.; Shen, Z.; Walsh, T.R.; Cai, C.; Wang, Y.; et al. Contaminated in-house environment contributes to the persistence and transmission of NDM-producing bacteria in a Chinese poultry farm. Environ. Int. 2020, 139, 105715. [Google Scholar] [CrossRef] [PubMed]
- ECDC. European Centre for Disease Prevention and Control. Carbapenem-Resistant Enterobacteriaceae, second Update; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Schrijver, R.; Stijntjes, M.; Rodríguez-Baño, J.; Tacconelli, E.; Rajendran, N.B.; Voss, A. Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 2018, 24, 577–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonardi, S.; Pitino, R. Carbapenemase-producing bacteria in food-producing animals, wildlife and environment: A challenge for human health. Ital. J. Food Saf. 2019, 8, 7956. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef]
- Elshafiee, E.A.; Nader, S.M.; Dorgham, S.M.; Hamza, D. Carbapenem-resistant Pseudomonas aeruginosa originating from farm animals and people in Egypt. J. Veter.-Res. 2019, 63, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Zhang, H.; Du, H. Carbapenemases in Enterobacteriaceae: Detection and antimicrobial therapy. Front. Microbiol. 2019, 10, 1823. [Google Scholar] [CrossRef]
- Balloux, F.; Brynildsrud, O.B.; Van Dorp, L.; Shaw, L.P.; Chen, H.; Harris, K.A.; Wang, H.; Eldholm, V. From theory to practice: Translating whole-genome sequencing (WGS) into the clinic. Trends Microbiol. 2018, 26, 1035–1048. [Google Scholar] [CrossRef] [Green Version]
- Lynch, T.; Petkau, A.; Knox, N.; Graham, M.; Van Domselaar, G. A Primer on infectious disease bacterial genomics. Clin. Microbiol. Rev. 2016, 29, 881–913. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R. Analysis of antibiotic resistance regions in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 820–855. [Google Scholar] [CrossRef] [Green Version]
- Stokes, H.W.; Gillings, M.R. Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbiol. Rev. 2011, 35, 790–819. [Google Scholar] [CrossRef]
- Toleman, M.A.; Walsh, T.R. Combinatorial events of insertion sequences and ICE in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 912–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardiman, C.A.; Weingarten, R.A.; Conlan, S.; Khil, P.; Dekker, J.P.; Mathers, A.J.; Sheppard, A.E.; Segre, J.A.; Frank, K.M. Horizontal transfer of Carbapenemase-encoding plasmids and comparison with hospital epidemiology data. Antimicrob. Agents Chemother. 2016, 60, 4910–4919. [Google Scholar] [CrossRef] [Green Version]
- Liapis, E.; Bour, M.; Triponney, P.; Jové, T.; Zahar, J.-R.; Valot, B.; Jeannot, K.; Plésiat, P. Identification of diverse integron and plasmid structures carrying a novel Carbapenemase among Pseudomonas species. Front. Microbiol. 2019, 10, 404. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, Y.; Akeda, Y.; Sakamoto, N.; Takeuchi, D.; Motooka, D.; Nakamura, S.; Hagiya, H.; Yamamoto, N.; Nishi, I.; Yoshida, H.; et al. Genetic characterization of blaNDM-harboring plasmids in carbapenem-resistant Escherichia coli from Myanmar. PLoS ONE 2017, 12, e0184720. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Miriagou, V.; Bertini, A.; Loli, A.; Colinon, C.; Villa, L.; Whichard, J.M.; Rossolini, G.M. Replicon typing of plasmids encoding resistance to newer β-Lactams. Emerg. Infect. Dis. 2006, 12, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Cuzon, G.; Naas, T.; Truong, H.; Villegas, M.-V.; Wisell, K.T.; Carmeli, Y.; Gales, A.C.; Navon-Venezia, S.; Quinn, J.P.; Nordmann, P. Worldwide diversity of Klebsiella pneumoniae that produce β-Lactamase blaKPC-2gene1. Emerg. Infect. Dis. 2010, 16, 1349–1356. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Yu, H.; Wang, M.; Zhang, H.; Lv, J.; Tang, Y.-W.; Kreiswirth, B.N.; Du, H.; Chen, L.; et al. New Delhi Metallo-β-Lactamase 5-producing Klebsiella pneumoniae sequence type 258, Southwest China, 2017. Emerg. Infect. Dis. 2019, 25, 1209–1213. [Google Scholar] [CrossRef]
- Lopes, B.S.; Amyes, S.G.B. Role of ISAba1 and ISAba125 in governing the expression of bla ADC in clinically relevant Acinetobacter baumannii strains resistant to cephalosporins. J. Med. Microbiol. 2012, 61, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, S.; Poirel, L.; Croize, J.; Recule, C.; Nordmann, P. In vivo selection of reduced susceptibility to Carbapenems in Acinetobacter baumannii related to ISAba1-mediated overexpression of the natural blaOXA-66 Oxacillinase gene. Antimicrob. Agents Chemother. 2009, 53, 2657–2659. [Google Scholar] [CrossRef] [Green Version]
- Bontron, S.; Nordmann, P.; Poirel, L. Transposition of Tn125encoding the NDM-1 Carbapenemase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, AAC.01755-16. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bhattacharjee, A.; Naha, S.; Majumdar, T.; Debbarma, S.K.; Kaur, H.; Dutta, S.; Basu, S. Molecular characterization of NDM-1-producing Klebsiella pneumoniae ST29, ST347, ST1224, and ST2558 causing sepsis in neonates in a tertiary care hospital of North-East India. Infect. Genet. Evol. 2019, 69, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Antunes, N.T.; Lamoureaux, T.L.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Class D β-Lactamases: Are they all Carbapenemases? Antimicrob. Agents Chemother. 2014, 58, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheruvanky, A.; Stoesser, N.; Sheppard, A.E.; Crook, D.W.; Hoffman, P.S.; Weddle, E.; Carroll, J.; Sifri, C.D.; Chai, W.; Barry, K.; et al. Enhanced Klebsiella pneumoniae Carbapenemase expression from a novel Tn4401 deletion. Antimicrob. Agents Chemother. 2017, 61, e00025-17. [Google Scholar] [CrossRef] [Green Version]
- Moellering, R.C. NDM-1-A cause for worldwide concern. New Engl. J. Med. 2010, 363, 2377–2379. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Toleman, M.A.; Walsh, T.R. Does broad-spectrum beta-lactam resistance due to NDM-1 herald the end of the antibiotic era for treatment of infections caused by Gram-negative bacteria? J. Antimicrob. Chemother. 2011, 66, 689–692. [Google Scholar] [CrossRef]
- Li, X.; Mu, X.; Zhang, P.; Zhao, D.-D.; Ji, J.; Quan, J.; Zhu, Y.; Yu, Y. Detection and characterization of a clinical Escherichia coli ST3204 strain coproducing NDM-16 and MCR-1. Infect. Drug Resist. 2018, 11, 1189–1195. [Google Scholar] [CrossRef]
- Yang, Q.E.; Agouri, S.R.; Tyrrell, J.M.; Walsh, T.R. Heavy metal resistance genes are associated with blaNDM-1- and blaCTX-M-15-carrying Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e02642-17. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.M.; Stegger, M.; Aziz, M.; Johnson, T.J.; Waits, K.; Nordstrom, L.; Gauld, L.; Weaver, B.; Rolland, D.; Statham, S.; et al. Escherichia coli ST131-H22 as a foodborne uropathogen. mBio 2018, 9, e00470-18. [Google Scholar] [CrossRef] [Green Version]
- Pecora, N.D.; Zhao, X.; Nudel, K.; Hoffmann, M.; Li, N.; Onderdonk, A.B.; Yokoe, D.; Brown, E.; Allard, M.; Bry, L. Diverse vectors and mechanisms spread New Delhi Metallo-β-Lactamases among Carbapenem-resistant Enterobacteriaceae in the Greater Boston area. Antimicrob. Agents Chemother. 2018, 63, e02040-18. [Google Scholar] [CrossRef] [Green Version]
- Muir, A.; Weinbren, M. New Delhi metallo-β-lactamase: A cautionary tale. J. Hosp. Infect. 2010, 75, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and their bacterial producers in health care settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrër, A.; Poirel, L.; Yilmaz, M.; Akan, O.A.; Feriha, C.; Cuzon, G.; Matar, G.; Honderlick, P.; Nordmann, P. Spread of OXA-48-encoding plasmid in Turkey and beyond. Antimicrob. Agents Chemother. 2010, 54, 1369–1373. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Dou, Y.; Zhou, K.; Chen, Y.; Han, L.; Guo, X.; Sun, J. Coexistence of blaOXA-48 and truncated blaNDM-1 on different plasmids in a Klebsiella pneumoniae isolate in China. Front. Microbiol. 2017, 8, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, T.; Cuzon, G.; Villegas, M.-V.; Lartigue, M.-F.; Quinn, J.P.; Nordmann, P. Genetic structures at the origin of acquisition of the β-Lactamase blaKPC gene. Antimicrob. Agents Chemother. 2008, 52, 1257–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galetti, R.; Andrade, L.N.; Varani, A.M.; Darini, A.L.C. A phage-like plasmid carrying blaKPC-2 gene in Carbapenem-resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 572. [Google Scholar] [CrossRef]
- Rasheed, J.K.; Biddle, J.W.; Anderson, K.F.; Washer, L.; Chenoweth, C.; Perrin, J.; Newton, D.W.; Patel, J.B. Detection of the Klebsiella pneumoniae Carbapenemase type 2 Carbapenem-hydrolyzing enzyme in clinical isolates of Citrobacter freundii and K. oxytoca carrying a common plasmid. J. Clin. Microbiol. 2008, 46, 2066–2069. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, C.; Bischoff, P.; Bender, J.; Kola, A.; Gastmeier, P.; Hummel, M.; Klefisch, F.-R.; Schoenrath, F.; Frühauf, A.; Pfeifer, Y. Plasmid-mediated transmission of KPC-2 Carbapenemase in Enterobacteriaceae in critically ill patients. Front. Microbiol. 2019, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.C.; Zhou, H.W.; Zhang, R.; Chen, G.-X. Emergence of Serratia marcescens, Klebsiella pneumoniae, and Escherichia coli isolates possessing the plasmid-mediated Carbapenem-hydrolyzing β-Lactamase KPC-2 in intensive care units of a Chinese hospital. Antimicrob. Agents Chemother. 2008, 52, 2014–2018. [Google Scholar] [CrossRef] [Green Version]




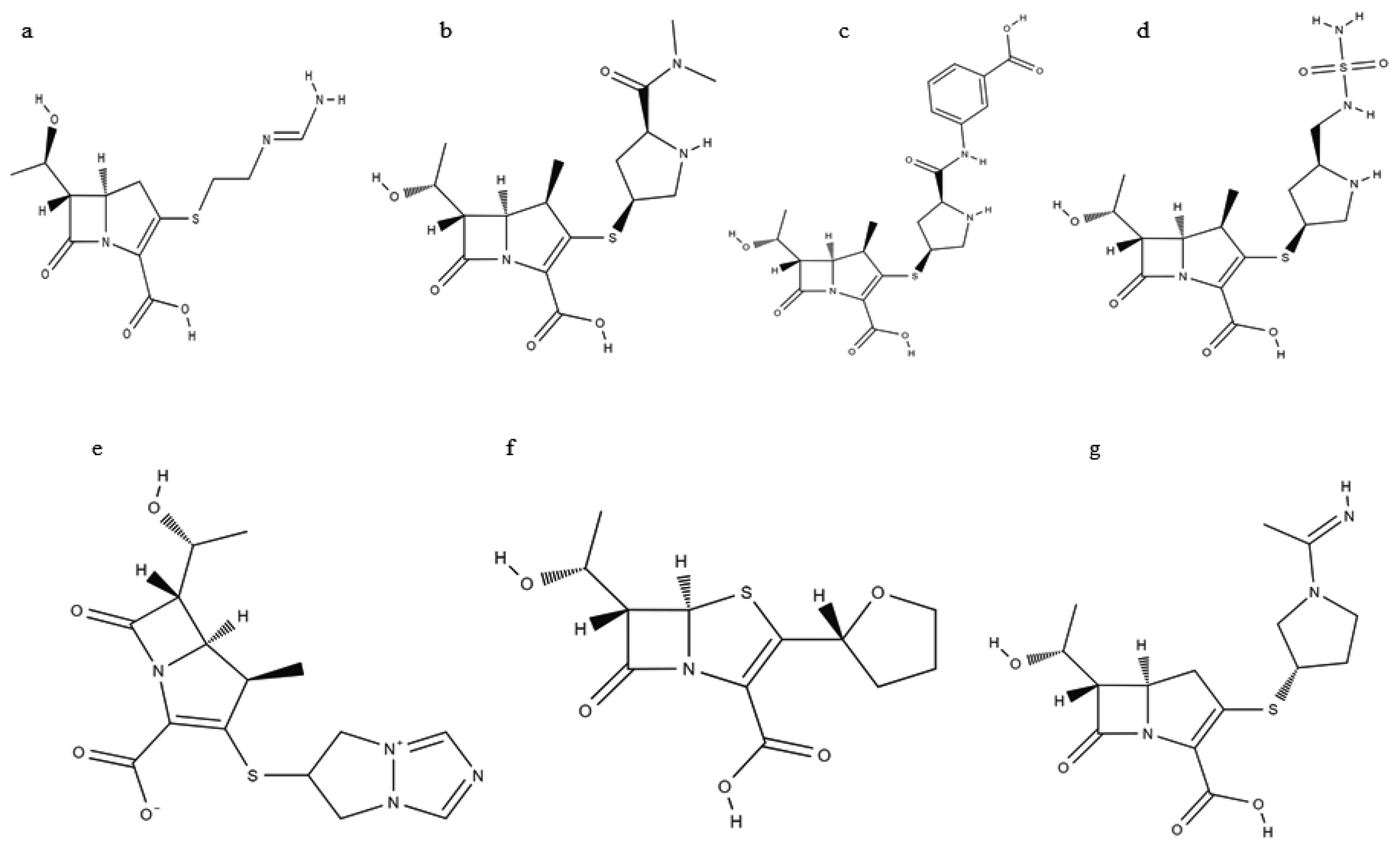{kind=link}
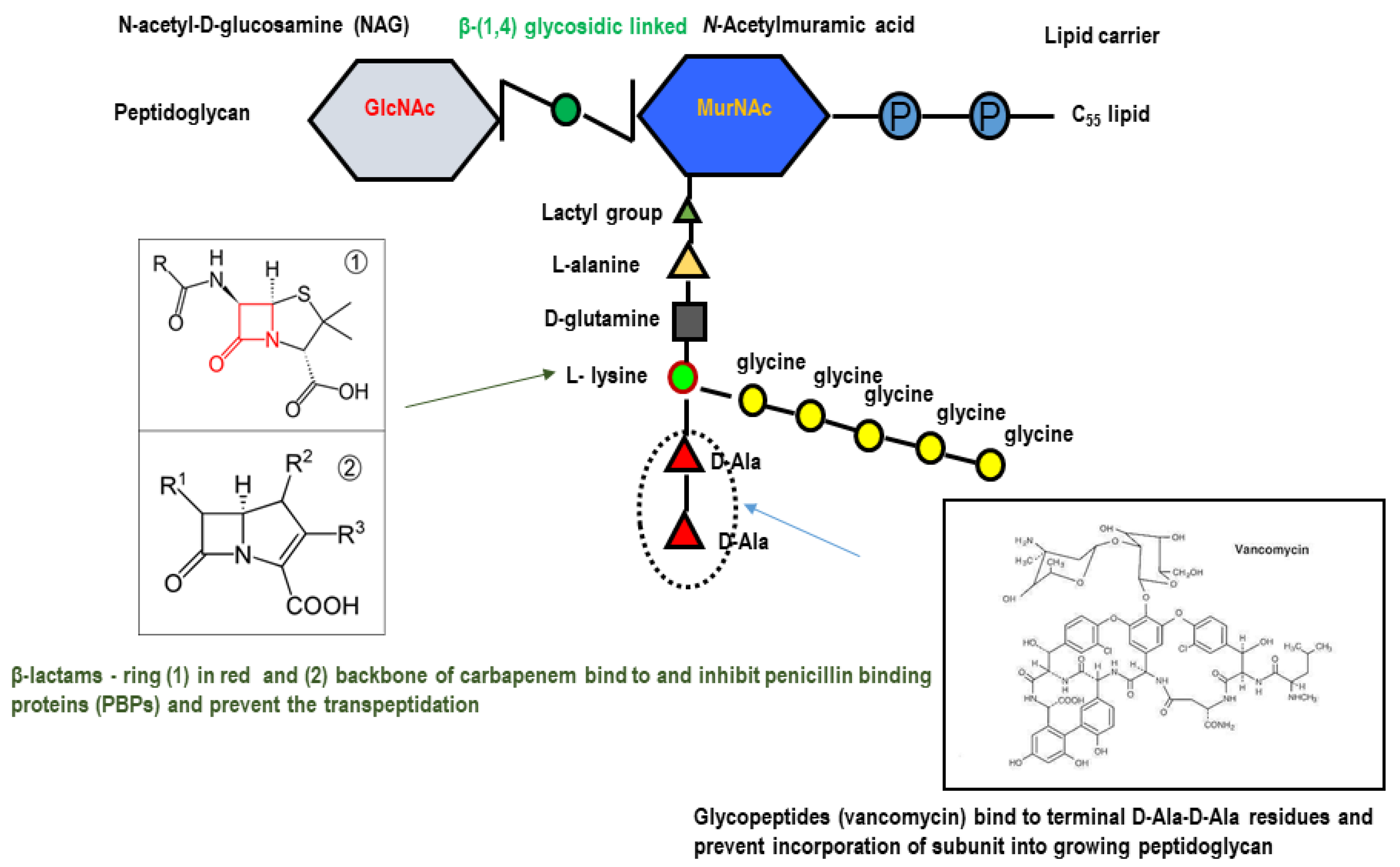{kind=link}
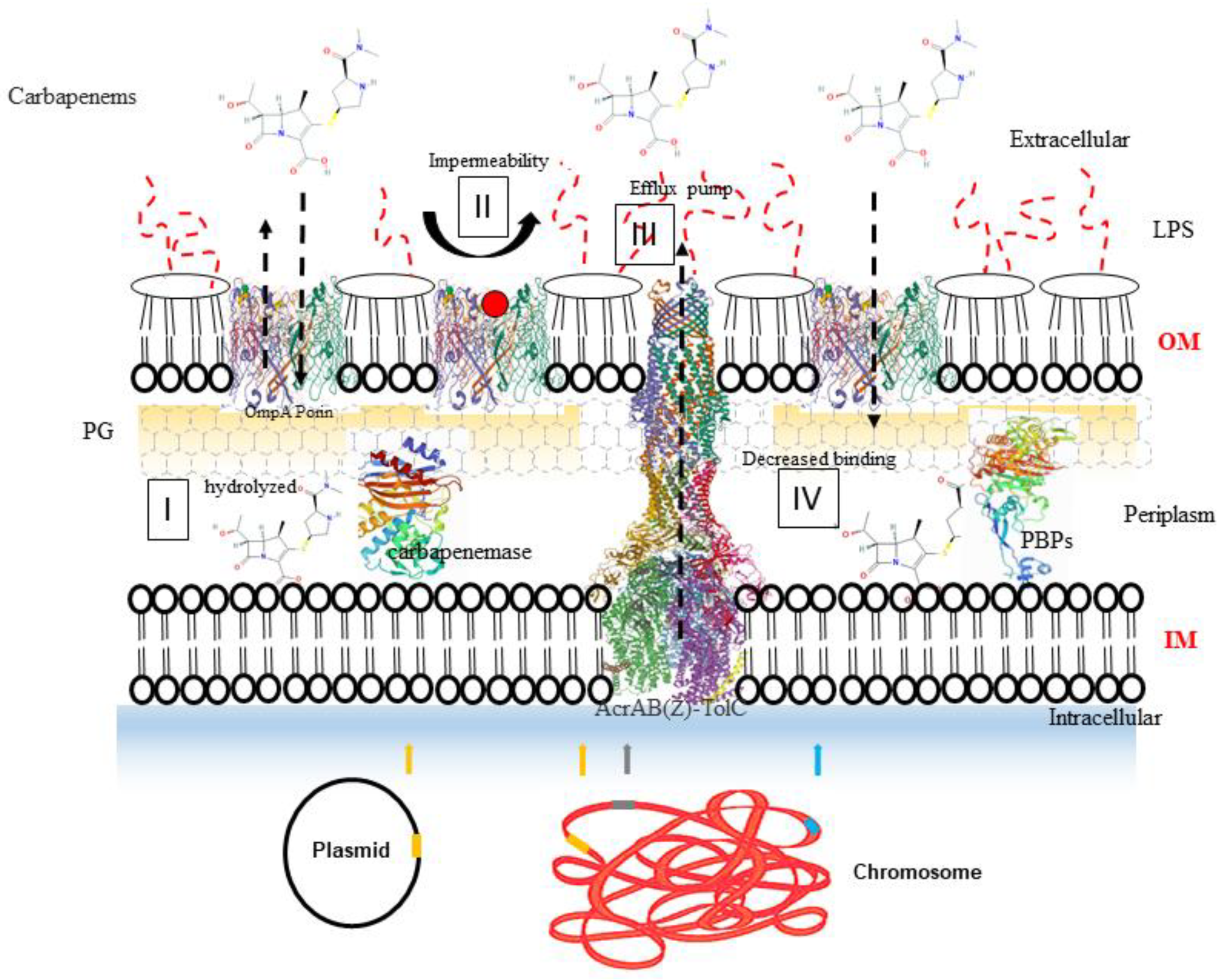{kind=link}
| Ambler Class. | Functional Class a | Representative Gene b | No. of Variants c | Active Site d | Substrate | Inhibitor(s) | Genetic Location | Species of Origin |
|---|---|---|---|---|---|---|---|---|
| A | 2F | KPC | 22 | Serine | carbapenems, cephalosporins, Penicillins | Clavulanic acid | Chromosomally encoded; IncFIIK2, IncF1A, IncI2, multiple types; Tn4401 | Klebsiella pneumoniae |
| IMI | 9 | Chromosomally encoded, IncF | Enterobacter cloacae | |||||
| SME | 5 | Chromosomally encoded, SmarGI1 novel genomic island | Serratia marcescens | |||||
| NMC-A | 1 | Chromosomally encoded | Enterobacter cloacae | |||||
| GES | 27 | Class I integrons | Pseudomonas aeruginosa | |||||
| B | 3 | NDM | 16 | Zinc | Most β-lactams including carbapenems | EDTA | IncA/C, multiple; ISAba125; Tn125 | Klebsiella pneumoniae |
| IMP | 56 | IncL/M, IncA/C, multiple types; class I integrons | Serratia marcescens | |||||
| VIM | 48 | IncN, IncI1, multiple types; class I integrons | Pseudomonas aeruginosa | |||||
| GIM | 2 | class I integrons | Pseudomonas aeruginosa | |||||
| SPM | 1 | Plasmid-mediated | Pseudomonas aeruginosa | |||||
| KHM | 1 | Most β-lactams except monobactams | Plasmid-mediated | Citrobacter freundii | ||||
| CcrA | 1 | Most β-lactams including carbapenems | Chromosomally encoded | Bacteroides fragilis | ||||
| BcII | 1 | Plasmid-mediated | Bacillus cereus | |||||
| CphA | 8 | Plasmid-mediated | Aeromonas hydrophilia | |||||
| L1 | not determined | Stenotrophomonas maltophilia | ||||||
| D | 2 | OXA | 489 | Serine | Most β-lactams including carbapenems | Clavulanic acid | IncL/M, Tn1999, IS1999, ColE plasmids, Tn2013, ISEcp1, ISAba125 | Klebsiella pneumoniae |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taggar, G.; Attiq Rehman, M.; Boerlin, P.; Diarra, M.S. Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment. Antibiotics 2020, 9, 693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100693
Taggar G, Attiq Rehman M, Boerlin P, Diarra MS. Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment. Antibiotics. 2020; 9(10):693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100693
Chicago/Turabian StyleTaggar, Gurleen, Muhammad Attiq Rehman, Patrick Boerlin, and Moussa Sory Diarra. 2020. "Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment" Antibiotics 9, no. 10: 693. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100693