Impetigo Animal Models: A Review of Their Feasibility and Clinical Utility for Therapeutic Appraisal of Investigational Drug Candidates


Abstract
:1. Introduction
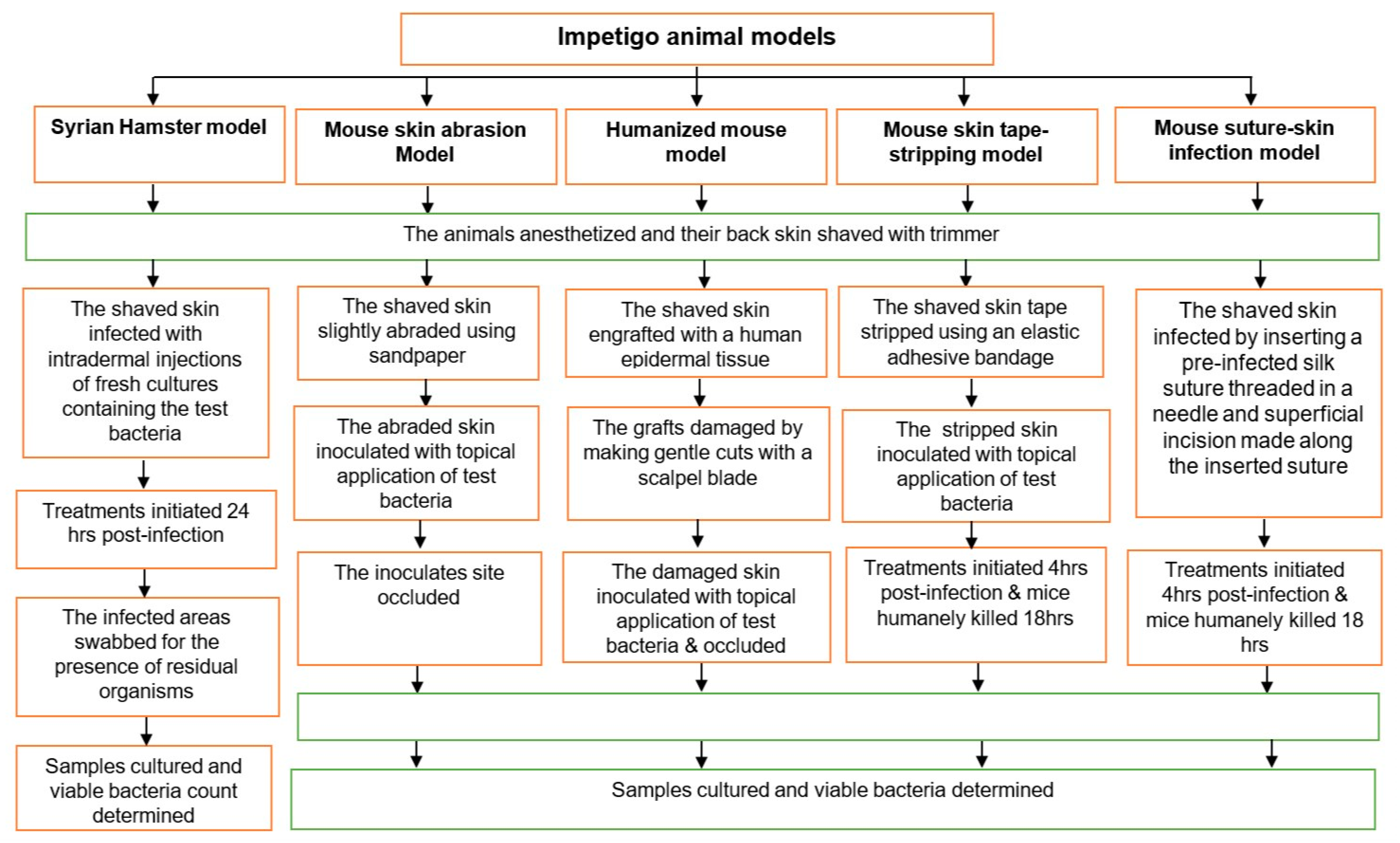
1.1. Syrian Hamster Impetigo Model
1.2. Mouse Skin Abrasion Impetigo Model
1.3. Humanized Mouse Model
1.4. Mouse Skin Tape-Stripping Model
1.5. Mouse Suture-Superficial Skin Infection Model
2. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Hay, R.J.; Johns, N.E.; Williams, H.C.; Bolliger, I.W.; Dellavalle, R.P.; Margolis, D.J.; Marks, R.; Naldi, L.; Weinstock, M.A.; Wulf, S.K.; et al. The Global Burden of Skin Disease in 2010: An Analysis of the Prevalence and Impact of Skin Conditions. J. Investig. Dermatol. 2014, 134, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, A.C.; Carapetis, J.R.; Currie, B.J.; Fowler, V.; Chambers, H.F.; Tong, S.Y.C. Sulfamethoxazole-Trimethoprim (Cotrimoxazole) for Skin and Soft Tissue Infections Including Impetigo, Cellulitis, and Abscess. Open Forum Infect. Dis. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, A.C.; Mahe, A.; Hay, R.J.; Andrews, R.M.; Steer, A.C.; Tong, S.Y.C.; Carapetis, J.R. The Global Epidemiology of Impetigo: A Systematic Review of the Population Prevalence of Impetigo and Pyoderma. PLoS ONE 2015, 10, e0136789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.B. Impetigo—Review. An. Bras. Dermatol. 2014, 89, 293–299. [Google Scholar] [CrossRef]
- May, P.J.; Tong, S.Y.C.; Steer, A.C.; Currie, B.J.; Andrews, R.M.; Carapetis, J.R.; Bowen, A.C. Treatment, prevention and public health management of impetigo, scabies, crusted scabies and fungal skin infections in endemic populations: A systematic review. Trop. Med. Int. Heal. 2019, 24, 280–293. [Google Scholar] [CrossRef]
- Koning, S.; Van Der Sande, R.; Verhagen, A.P.; A Van Suijlekom-Smit, L.W.; Morris, A.D.; Butler, C.C.; Berger, M.; Van Der Wouden, J.C. Interventions for impetigo. Cochrane Database Syst. Rev. 2012, 1, CD003261. [Google Scholar] [CrossRef] [Green Version]
- D’Cunha, N.M.; Peterson, G.M.; Baby, K.E.; Thomas, J. Impetigo: A need for new therapies in a world of increasing antimicrobial resistance. J. Clin. Pharm. Ther. 2017, 43, 150–153. [Google Scholar] [CrossRef] [Green Version]
- Steer, A.C.; Danchin, M.H.; Carapetis, J.R. Group A streptococcal infections in children. J. Paediatr. Child Health 2007, 43, 203–213. [Google Scholar] [CrossRef]
- Dajani, A.S.; Wannamaker, L.W. Experimental Infection of the Skin in the Hamster Simulating Human Impetigo. I. Natural History of the Infection. J. Infect. Dis. 1970, 122, 196–204. [Google Scholar] [CrossRef]
- Vila, J.; Hebert, A.A.; Torrelo, A.; López, Y.; Tato, M.; García-Castillo, M.; Cantón, R. Ozenoxacin: A review of preclinical and clinical efficacy. Expert Rev. Anti Infect. Ther. 2019, 17, 159–168. [Google Scholar] [CrossRef]
- Cole, C.; Gazewood, J. Diagnosis and treatment of impetigo. Am. Fam. Physician 2007, 75, 859–864. [Google Scholar] [PubMed]
- WHO. Antimicrobial Resistance: Global Report on Surveillance Geneva; WHO Press, World Health Organization: Geneva, Switzerland, 2014. Available online: https://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 5 July 2019).
- Bowen, A.C.; Tong, S.Y.C.; Chatfield, M.; Carapetis, J.R. The microbiology of impetigo in indigenous children: Associations between Streptococcus pyogenes, Staphylococcus aureus, scabies, and nasal carriage. BMC Infect. Dis. 2014, 14, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeoh, D.K.; Anderson, A.; Cleland, G.; Bowen, A.C. Are scabies and impetigo “normalised”? A cross-sectional comparative study of hospitalised children in northern Australia assessing clinical recognition and treatment of skin infections. PLoS Negl. Trop. Dis. 2017, 11, e0005726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangilinan, R.; Tice, A.; Tillotson, G. Topical antibiotic treatment for uncomplicated skin and skin structure infections: Review of the literature. Expert Rev. Anti Infect. Ther. 2009, 7, 957–965. [Google Scholar] [CrossRef]
- Bohaty, B.R.; Choi, S.; Cai, C.; Hebert, A.A. Clinical and bacteriological efficacy of twice daily topical retapamulin ointment 1% in the management of impetigo and other uncomplicated superficial skin infections. Int. J. Women’s Dermatol. 2015, 1, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonev, B.B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Publishes List of Bacteria for Which New Antibiotics are Urgently Needed Geneva. 2017. Available online: https://www.who.int/en/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 1 October 2019).
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- The Pew Charitable Trusts. Antibiotics Currently in Clinical Development 2017 (cited 2019 October 17). Available online: http://www.pewtrusts.org/~/media/assets/2017/05/antibiotics-currently-in-clinical-development-03-2017.pdf?la=en (accessed on 17 October 2019).
- FDA. Guideline for Clinical Evaluation of Anti-Nfective Drugs for Systemic Administration USA: U.S Department of Health and Human Service, Food and Drug Adminstration 1977. Available online: https://catalog.hathitrust.org/Record/000090753 (accessed on 10 July 2020).
- Zak, O.; O’Reilly, T. Animal models in the evaluation of antimicrobial agents. Antimicrob. Agents Chemother. 1991, 35, 1527–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, T.K. In vivo Pharmacological Disease Models for Psoriasis and Atopic Dermatitis in Drug Discovery. Basic Clin. Pharmacol. Toxicol. 2006, 99, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Avci, P.; Sadasivam, M.; Gupta, A.; De Melo, W.C.; Huang, Y.-Y.; Yin, R.; Chandran, R.; Kumar, R.; Otufowora, A.; Nyame, T.; et al. Animal models of skin disease for drug discovery. Expert Opin. Drug Discov. 2013, 8, 331–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, D. The use of animal models in studying genetic disease: Transgenesis and induced mutation. Nat. Educ. 2008, 1, 70. [Google Scholar]
- Barré-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Futur. Sci. OA 2015, 1, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Craig, W. Relevance of animal models for clinical treatment. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12 (Suppl. 1), S55–S57. [Google Scholar] [CrossRef]
- Beam, J.T.R.; Gilbert, D.N.; Kunin, C.M. General Guidelines for the Clinical Evaluation of Anti-Infective Drug Products. Clin. Infect. Dis. 1992, 15 (Suppl. 1), S5–S32. [Google Scholar] [CrossRef] [PubMed]
- Soriano, F.; Aguilar, L.; Ponte, C. In vitro antibiotic sensitivity testing breakpoints and therapeutic activity in induced infections in animal models. J. Chemother. 1997, 9 (Suppl. 1), 36–46. [Google Scholar]
- Watson, M.E.J.; Neely, M.N.; Caparon, M.G. Animal models of Streptococcus pyogenes infection. In Streptococcus pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Zhao, M.; Lepak, A.J.; Andes, D. Animal models in the pharmacokinetic/pharmacodynamic evaluation of antimicrobial agents. Bioorg. Med. Chem. 2016, 24, 6390–6400. [Google Scholar] [CrossRef] [PubMed]
- Ghiselli, R.; Cirioni, O.; Giacometti, A.; Scalise, A.; Simonetti, O.; Mocchegiani, F.; Orlando, F.; Goteri, G.; Della Vittoria, A.; Filosa, A.; et al. Comparative Efficacy of Topical Versus Systemic Teicoplanin in Experimental Model of Wound Infections. J. Surg. Res. 2008, 144, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Van Der Staay, F.J.; Arndt, S.S.; Nordquist, R.E. Evaluation of animal models of neurobehavioral disorders. Behav. Brain Funct. 2009, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRipley, R.J.; Whitney, R.R. Characterization and Quantitation of Experimental Surgical-Wound Infections Used to Evaluate Topical Antibacterial Agents. Antimicrob. Agents Chemother. 1976, 10, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRipley, R.J.; Whitney, R.R. Responsiveness of Experimental Surgical-Wound Infections to Topical Chemotherapy. Antimicrob. Agents Chemother. 1976, 10, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Missiakas, D.; Schneewind, O. Mouse models for infectious diseases caused by Staphylococcus aureus. J. Immunol. Methods 2014, 410, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; Kobayashi, S.D.; Braughton, K.R.; DeLeo, F.R. Mouse Model of Staphylococcus aureus Skin Infection. Recent Results Cancer Res. 2013, 1031, 109–116. [Google Scholar] [CrossRef]
- Yamakawa, T.; Mitsuyama, J.; Hayashi, K. In vitro and in vivo antibacterial activity of T-3912, a novel non-fluorinated topical quinolone. J. Antimicrob. Chemother. 2002, 49, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Kharkwal, G.B.; Tanaka, M.; Huang, Y.-Y.; De Arce, V.J.B.; Hamblin, M.R. Animal models of external traumatic wound infections. Virulence 2011, 2, 296–315. [Google Scholar] [CrossRef] [Green Version]
- Cushing, A.H.; Mortimer, E.A. A Hamster Model for Streptococcal Impetigo. J. Infect. Dis. 1970, 122, 224–226. [Google Scholar] [CrossRef]
- Dajani, A.S.; Hill, P.L.; Wannamaker, L.W. Experimental infection of the skin in the hamster simulating human impetigo. II. Assessment of various therapeutic regimens. Pediatrics 1971, 48, 83–90. [Google Scholar]
- Dajani, A.S.; Wannamaker, L.W. Experimental infection of the skin in the hamster simulating human impetigo. III. Interaction between Staphylococci and group A Streptococci. J. Exp. Med. 1971, 134, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Dajani, A.S.; Wannamaker, L.W. Experimental infection of the skin in the hamster simulating human impetigo. IV. Cellular responses after Streptococcal and Staphylococcal infections. Infect. Immun. 1972, 5, 942–946. [Google Scholar] [CrossRef] [Green Version]
- Gisby, J.; Bryant, J. Efficacy of a New Cream Formulation of Mupirocin: Comparison with Oral and Topical Agents in Experimental Skin Infections. Antimicrob. Agents Chemother. 2000, 44, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Abe, Y.; Akiyama, H.; Arata, J. Production of experimental Staphylococcal impetigo in mice. J. Dermatol. Sci. 1992, 4, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Scaramuzzino, D.A.; McNiff, J.M.; Bessen, D.E. Humanized In Vivo Model for Streptococcal Impetigo. Infect. Immun. 2000, 68, 2880–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, M.D.; Scaramuzzino, D.A.; Sjobring, U.; Olsen, A.; Frank, C.; Bessen, D.E. Role for a secreted cysteine proteinase in the establishment of host tissue tropism by group A streptococci. Mol. Microbiol. 2000, 38, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.D.; Sjöbring, U.; Luo, F.; Bessen, D.E. Roles of the plasminogen activator streptokinase and the plasminogen-associated M protein in an experimental model for streptococcal impetigo. Microbiology 2002, 148, 3933–3945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugelberg, E.; Norström, T.; Petersen, T.K.; Duvold, T.; Andersson, D.I.; Hughes, D. Establishment of a Superficial Skin Infection Model in Mice by Using Staphylococcus aureus and Streptococcus pyogenes. Antimicrob. Agents Chemother. 2005, 49, 3435–3441. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, I.; Hattori, S.; Hisatsune, J.; Ide, K.; Sugai, M.; Nishifuji, K. Staphylococcus aureus penetrate the interkeratinocyte spaces created by skin-infiltrating neutrophils in a mouse model of impetigo. Vet. Dermatol. 2016, 28, 126-e27. [Google Scholar] [CrossRef] [Green Version]
- Hahn, B.L.; Onunkwo, C.C.; Watts, C.J.; Sohnle, P.G. Systemic dissemination and cutaneous damage in a mouse model of Staphylococcal skin infections. Microb. Pathog. 2009, 47, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Tarragó, C.; Esquirol, L.P.; Arañó, A.; Lachamp, L.; D’Aniello, F.; Zsolt, I. Therapeutic efficacy of ozenoxacin in animal models of dermal infection with Staphylococcus aureus. Futur. Microbiol. 2018, 13, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Håkansson, J.; Björn, C.; Lindgren, K.; Sjöström, E.; Sjöstrand, V.; Mahlapuu, M. Efficacy of the Novel Topical Antimicrobial Agent PXL150 in a Mouse Model of Surgical Site Infections. Antimicrob. Agents Chemother. 2014, 58, 2982–2984. [Google Scholar] [CrossRef] [Green Version]
- Rittenhouse, S.; Singley, C.; Hoover, J.; Page, R.; Payne, D. Use of the Surgical Wound Infection Model to Determine the Efficacious Dosing Regimen of Retapamulin, a Novel Topical Antibiotic. Antimicrob. Agents Chemother. 2006, 50, 3886–3888. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.J.; Beale, A.S. Response of Streptococcus pyogenes to therapy with amoxicillin or amoxicillin-clavulanic acid in a mouse model of mixed infection caused by Staphylococcus aureus and Streptococcus pyogenes. Antimicrob. Agents Chemother. 1987, 31, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.J.; Beale, A.S.; Sutherland, R. Efficacy of topical mupirocin against an experimental Staphylococcus aureus surgical wound infection. J. Antimicrob. Chemother. 1985, 16, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Berry, V.; Page, R.; Satterfield, J.; Singley, C.; Straub, R.; Woodnutt, G. Comparative efficacy of gemifloxacin in experimental models of pyelonephritis and wound infection. J. Antimicrob. Chemother. 2000, 45, 87. [Google Scholar] [CrossRef] [Green Version]
- Håkansson, J.; Ringstad, L.; Umerska, A.; Johansson, J.; Andersson, T.; Boge, L.; Rozenbaum, R.T.; Sharma, P.K.; Tollbäck, P.; Björn, C.; et al. Characterization of the in vitro, ex vivo, and in vivo Efficacy of the Antimicrobial Peptide DPK-060 Used for Topical Treatment. Front. Cell. Infect. Microbiol. 2019, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, E.; Lengeling, A. Genetic regulation of host responses to Group A Streptococcus in mice. Briefings Funct. Genom. Proteom. 2005, 4, 248–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sladden, M.J.; Johnston, A.G. Common skin infections in children. BMJ 2004, 329, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangert, S.; Levy, M.; Hebert, A.A. Bacterial Resistance and Impetigo Treatment Trends: A Review. Pediatr. Dermatol. 2012, 29, 243–248. [Google Scholar] [CrossRef]
- Leyden, J.J.; Stewart, R.; Kligman, A.M. Experimental Infections with Group A Streptococci in Humans. J. Investig. Dermatol. 1980, 75, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Duncan, W.C.; McBride, M.E.; Knox, J.M. Experimental Production of Infections in Humans. J. Investig. Dermatol. 1970, 54, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Marples, R.R.; Kligman, A.M. Experimental Staphylococcus aureus Infections in Humans. J. Investig. Dermatol. 1971, 57, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Arata, J.; Kashiwa, N. Antibiotic effects on bacterial counts in skin lesions of experimental Staphylococcal skin infections in the hamster. J. Dermatol. 1984, 11, 67–72. [Google Scholar] [CrossRef]
- Baxter, V.K.; Griffin, D.E. Chapter 10—Animal models: No model is perfect, but many are useful. In Viral Pathogenesis, 3rd ed.; Katze, M.G., Korth, M.J., Law, G.L., Nathanson, N., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 125–138. [Google Scholar]
- Justice, M.J.; Dhillon, P. Using the mouse to model human disease: Increasing validity and reproducibility. Dis. Model. Mech. 2016, 9, 101–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ericsson, A.C.; Crim, M.J.; Franklin, C.L. A Brief History of Animal Modeling. Mo. Med. 2013, 110, 201–205. [Google Scholar] [PubMed]
- Bunce, C.; Wheeler, L.; Reed, G.; Musser, J.; Barg, N. Murine model of cutaneous infection with gram-positive cocci. Infect. Immun. 1992, 60, 2636–2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, Y.; Akiyama, H.; Arata, J. Production of Staphylococcal impetigo-like lesion on human skin explants in culture. J. Dermatol. Sci. 1993, 5, 150–164. [Google Scholar] [CrossRef]
- Marples, R.R.; Kligman, A.M. Methods for Evaluating Topical Antibacterial Agents on Human Skin. Antimicrob. Agents Chemother. 1974, 5, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Sumitomo, T.; Mori, Y.; Nakamura, Y.; Honda-Ogawa, M.; Nakagawa, S.; Yamaguchi, M.; Matsue, H.; Terao, Y.; Nakata, M.; Kawabata, S. Streptococcal Cysteine Protease-Mediated Cleavage of Desmogleins Is Involved in the Pathogenesis of Cutaneous Infection. Front. Cell. Infect. Microbiol. 2018, 8, 10. [Google Scholar] [CrossRef]
- Iwatsuki, K.; Yamasaki, O.; Morizane, S.; Oono, T. Staphylococcal cutaneous infections: Invasion, evasion and aggression. J. Dermatol. Sci. 2006, 42, 203–214. [Google Scholar] [CrossRef]
- Mempel, M.; Schnopp, C.; Hojka, M.; Fesq, H.; Weidinger, S.; Schaller, M.; Korting, H.; Ring, J.; Abeck, D. Invasion of human keratinocytes by Staphylococcus aureus and intracellular bacterial persistence represent haemolysin-independent virulence mechanisms that are followed by features of necrotic and apoptotic keratinocyte cell death. Br. J. Dermatol. 2002, 146, 943–951. [Google Scholar] [CrossRef]
- Onunkwo, C.C.; Hahn, B.L.; Sohnle, P.G. Clearance of experimental cutaneous Staphylococcus aureus infections in mice. Arch. Dermatol. Res. 2010, 302, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Shamaei-Tousi, A.; Liu, Y.; Coates, A.R.M. A New Approach for the Discovery of Antibiotics by Targeting Non-Multiplying Bacteria: A Novel Topical Antibiotic for Staphylococcal Infections. PLoS ONE 2010, 5, e11818. [Google Scholar] [CrossRef]
- Hu, Y.; Coates, A.R.M. Enhancement by novel anti-methicillin-resistant Staphylococcus aureus compound HT61 of the activity of neomycin, gentamicin, mupirocin and chlorhexidine: In vitro and in vivo studies. J. Antimicrob. Chemother. 2012, 68, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, E.H.; Gutekunst, D.P.; Chamberlain, E.R. Evaluation of furazolium chloride in topical treatment of model infections in laboratory animals. Antimicrob. Agents Chemother. 1967, 7, 497–501. [Google Scholar]
- Alexander, J.W.; Kaplan, J.Z.; Altemeier, W.A. Role of Suture Materials in the Development of Wound Infection. Ann. Surg. 1967, 165, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Elek, S.D. Experimental Staphylococcal infections in the skin of man. Ann. N. Y. Acad. Sci. 1956, 65, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Hillman, E.M.; Amoozegar, C.B.; Wang, T.; McCaslin, A.F.H.; Bouchard, M.B.; Mansfield, J.; Levenson, R.M. In vivo optical imaging and dynamic contrast methods for biomedical research. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 4620–4643. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Zussman, J.; Donegan, N.P.; Ramos, R.I.; Garcia, N.C.; Uslan, D.Z.; Iwakura, Y.; Simon, S.I.; Cheung, A.L.; Modlin, R.L.; et al. Noninvasive In Vivo Imaging to Evaluate Immune Responses and Antimicrobial Therapy against Staphylococcus aureus and USA300 MRSA Skin Infections. J. Investig. Dermatol. 2011, 131, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Ramos, R.I.; Cho, J.S.; Donegan, N.P.; Cheung, A.L.; Miller, L.S. In Vivo Bioluminescence Imaging to Evaluate Systemic and Topical Antibiotics against Community-Acquired Methicillin-Resistant Staphylococcus aureus—Infected Skin Wounds in Mice. Antimicrob. Agents Chemother. 2012, 57, 855–863. [Google Scholar] [CrossRef] [Green Version]
- Albareda, N.; Zeichner, J.; Rosenberg, N. A Randomized Vehicle-Controlled Trial to Assess the Efficacy, Safety, and Tolerability of Ozenoxacin 1% Cream in 412 Patients 2 Months and Older with Impetigo. Ski. J. Cutan. Med. 2017, 1, s103. [Google Scholar] [CrossRef]
- Gropper, S.; Albareda, N.; Chelius, K.; Kruger, D.; Mitha, I.; Vahed, Y.; Gani, M.; García-Alonso, F. Ozenoxacin 1% cream in the treatment of impetigo: A multicenter, randomized, placebo- and retapamulin-controlled clinical trial. Futur. Microbiol. 2014, 9, 1013–1023. [Google Scholar] [CrossRef]
- Koning, S.; Van Der Wouden, J.; Chosidow, O.; Twynholm, M.; Singh, K.; Scangarella, N.; Oranje, A.; Van Der Wouden, J.C. Efficacy and safety of retapamulin ointment as treatment of impetigo: Randomized double-blind multicentre placebo-controlled trial. Br. J. Dermatol. 2008, 158, 1077–1082. [Google Scholar] [CrossRef]
- Goldfarb, J.; Crenshaw, D.; O’Horo, J.; Lemon, E.; Blumer, J.L. Randomized clinical trial of topical mupirocin versus oral erythromycin for impetigo. Antimicrob. Agents Chemother. 1988, 32, 1780–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rist, T.; Parish, L.C.; Capin, L.R.; Sulica, V.; Bushnell, W.D.; Cupo, M.A. A comparison of the efficacy and safety of mupirocin cream and cephalexin in the treatment of secondarily infected eczema. Clin. Exp. Dermatol. 2002, 27, 14–20. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |


{kind=link}
| Model | Bacteria Tested | Host Animal | Anaesthetic Agent | Inoculation Route | Inoculum Dose | Time for Infection Occurrence | Antimicrobial Agents Tested | Sampling Method |
|---|---|---|---|---|---|---|---|---|
| Hamster impetigo model [9,40,41,42,43,44] | S. aureus or S. pyogenes | Hamster (Golden Syrian type, 6–8 weeks, 80–120 g, n = 4–75) | Isoflurane (3%, inhalation) | Intradermal injections | 1.2 × 107 CFU | 24 h post-inoculation | Gentamicin ointment, Bacitracin ointment, Benzathine penicillin G injection, Procaine penicillin injection | Swabbing the lesion surface |
| Mouse skin abrasion impetigo model [45] | S. aureus | Mouse (ddY type, female, 5 weeks old, ~20 g, n = 5) | Not reported | Topical/epicutaneous inoculation to slightly damaged skin by sandpaper | 7 × 104 CFU | 24 h post-inoculation | No antimicrobial agent used | Biopsy of infected skin area |
| Humanized mouse impetigo Model [46,47,48] | S. pyogenes | Mouse (SCID type, female, 4–6-week-old, n = no report) | Ketamine-Xylazine (Intraperitoneal injection) | Topical/epicutaneous inoculation to slightly damaged skin by sandpaper, scalpel, or tape stripping | 50 CFU | 1 week post-inoculation | No antimicrobial agent used | Biopsy of infected skin area |
| Mouse skin tape-stripping model [49,50,51] | S. aureus or S. pyogenes | Mouse (BALB/c type, female, 6–8-weeks-old, n = no report) | 1:1:2 v/v mixture of hypnorm (fentanyl, fluanisone), dormicum (midazolam) and distilled water, Intraperitoneal injection) | Topical/epicutaneous inoculation to slightly damaged skin by tape stripping | 107 cells | 4 h post-inoculation | Fusidic acid ointment | Biopsy of infected skin area |
| Mouse suture-superficial skin infection model [34,35,44,52,53,54,55,56,57,58] | S. aureus and/or S. pyogenes | Mouse (CF-1, CD1, and MF1 type, female and male, 18–20 g, n = 10–50) | Sodium pentobarbital (30 mg/kg, Intraperitoneal injection) Or Diazepam plus fentanyl Fluanisone (1.25 mg/kg plus 0.5 mL/kg Intramuscular injection) | Topical/epicutaneous inoculation by insertion of an infected suture | 103–105 cells | 6 h post-inoculation | Gentamicin cream, Polymyxin B-bacitracin-neomycin ointment, Retapamulin ointment, Fusidic acid cream, Muprocin ointment and cream, Ozenoxacin cream, amoxicillin or amoxicillin-clavulanic acid oral, Gemifloxacin oral | Swabbing the lesion surface Or Biopsy of infected skin area |
| Models | General Strengths | General Limitations |
|---|---|---|
| Hamster impetigo model [9,40,41,42,43,44] |
|
|
| Mouse skin abrasion impetigo model [45] |
|
|
| Humanized mouse impetigo Model [46,47,48] |
|
|
| Mouse skin tape-stripping model [49,50,51] |
|
|
| Mouse suture-superficial skin infection model [34,35,44,52,53,54,55,56,57,58] |
|
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrha, S.; Bartholomaeus, A.; Tesfaye, W.; Thomas, J. Impetigo Animal Models: A Review of Their Feasibility and Clinical Utility for Therapeutic Appraisal of Investigational Drug Candidates. Antibiotics 2020, 9, 694. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100694
Abrha S, Bartholomaeus A, Tesfaye W, Thomas J. Impetigo Animal Models: A Review of Their Feasibility and Clinical Utility for Therapeutic Appraisal of Investigational Drug Candidates. Antibiotics. 2020; 9(10):694. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100694
Chicago/Turabian StyleAbrha, Solomon, Andrew Bartholomaeus, Wubshet Tesfaye, and Jackson Thomas. 2020. "Impetigo Animal Models: A Review of Their Feasibility and Clinical Utility for Therapeutic Appraisal of Investigational Drug Candidates" Antibiotics 9, no. 10: 694. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100694