Dalbavancin, Vancomycin and Daptomycin Alone and in Combination with Cefazolin against Resistant Phenotypes of Staphylococcus aureus in a Pharmacokinetic/Pharmacodynamic Model

 ,
, 
Abstract
:1. Introduction
2. Results
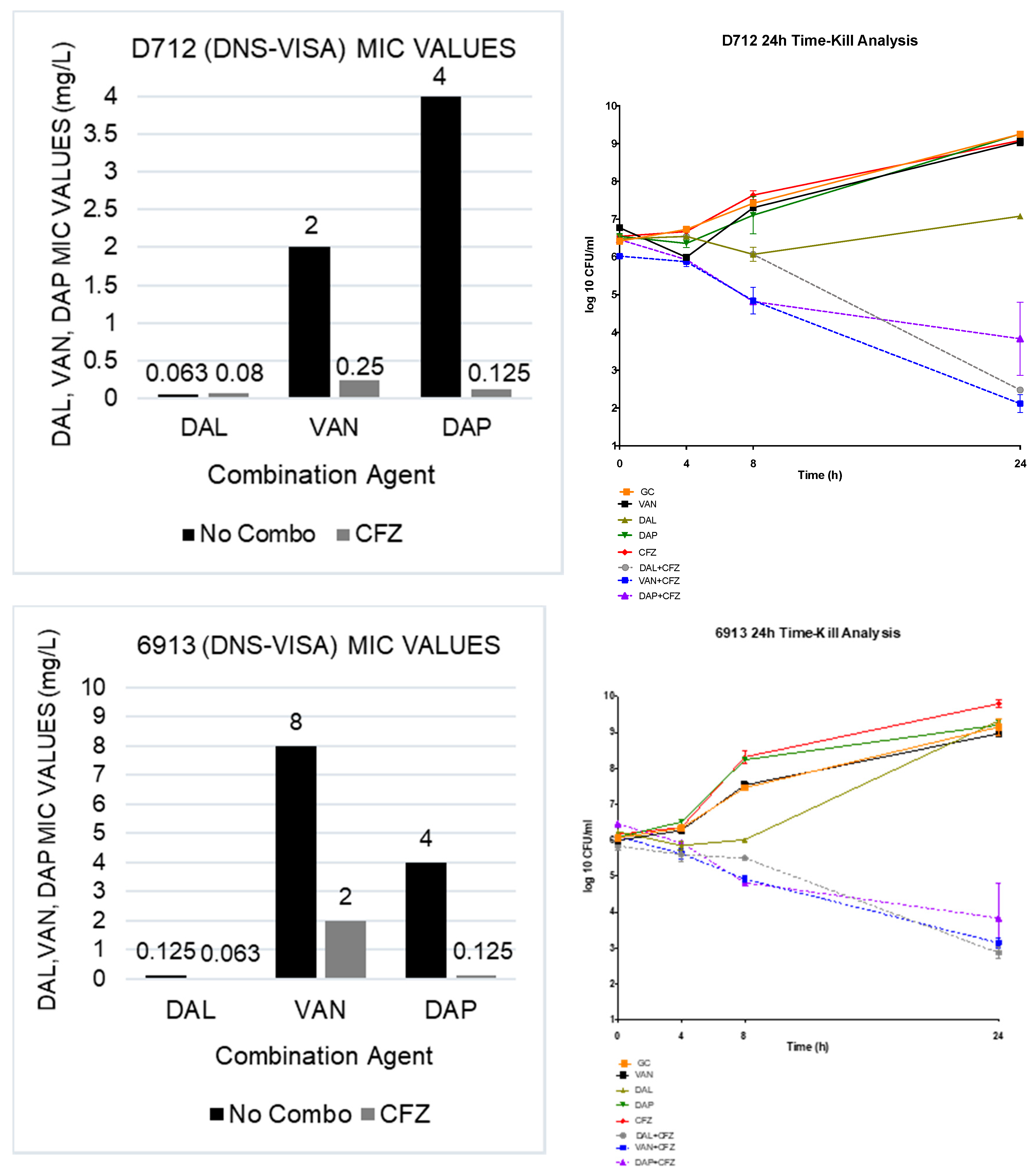
2.1. Susceptibility Testing
2.2. In Vitro Time-Kill Analysis
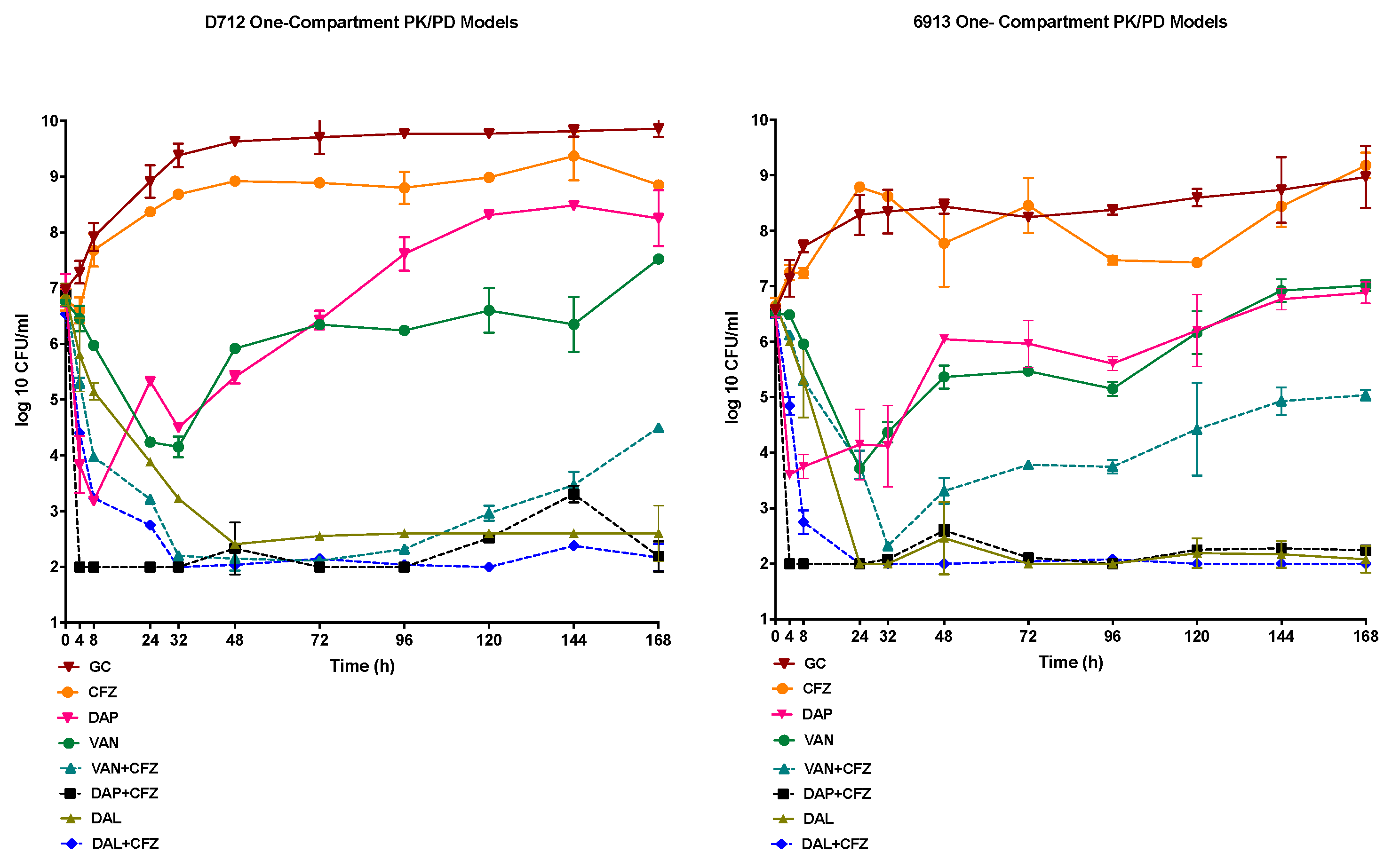
2.3. In Vitro One-Compartment PK/PD Model
2.3.1. Pharmacokinetics
2.3.2. Pharmacodynamics
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antimicrobials and Media
4.3. Susceptibility Testing
4.4. In Vitro Time-Kill Analysis
4.5. In Vitro PK/PD Model
4.6. Pharmacodynamic Analysis
4.7. Pharmacokinetic Analysis
4.8. Resistance
4.9. Population Analysis Profile (PAP) Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Gardete, S.; Tomasz, A. Mechanisms of vancomycin resistance in Staphylococcus aureus. J. Clin. Investig. 2014, 124, 2836–2840. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Dick, J.D.; Perl, T.M. Vancomycin resistance in staphylococci. Clin. Microbiol. Rev. 2002, 15, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.N.; McKinnell, J.; Yeaman, M.R.; Rubio, A.; Nast, C.C.; Chen, L.; Kreiswirth, B.N.; Bayer, A.S. In vitro cross-resistance to daptomycin and host defense cationic antimicrobial peptides in clinical methicillin-resistant Staphylococcus aureus isolates. Antimicrob. Agents Chemother. 2011, 55, 4012–4018. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Tominaga, E.; Neoh, H.M.; Hiramatsu, K. Correlation between Reduced Daptomycin Susceptibility and Vancomycin Resistance in Vancomycin-Intermediate Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 1079–1082. [Google Scholar] [CrossRef] [Green Version]
- Barber, K.E.; Ireland, C.E.; Bukavyn, N.; Rybak, M.J. Observation of “seesaw effect” with vancomycin, teicoplanin, daptomycin and ceftaroline in 150 unique MRSA strains. Infect. Dis. Ther. 2014, 3, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Stefani, S.; Goglio, A. Methicillin-resistant Staphylococcus aureus: Related infections and antibiotic resistance. Int. J. Infect. Dis. 2010, 14 (Suppl. 4), S19–S22. [Google Scholar] [CrossRef] [Green Version]
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S.; National Healthcare Safety Network (NHSN) Team; et al. Antimicrobial-resistant pathogens associated with healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control Hosp. Epidemiol. 2013, 34, 1–14. [Google Scholar] [CrossRef]
- Dalbavancin. Drugs and Lactation Database (LactMed); Bethesda: Rockville, MD, USA, 2006. [Google Scholar]
- Smith, J.R.; Roberts, K.D.; Rybak, M.J. Dalbavancin: A Novel Lipoglycopeptide Antibiotic with Extended Activity Against Gram-Positive Infections. Infect. Dis. Ther. 2015, 4, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Azanza, J.R.; Sadaba, B.; Reis, J. Dalbavancin: Pharmacokinetic and pharmacodynamic parameters. Enferm. Infecc. Microbiol. Clin. 2017, 35 (Suppl. 1), 22–27. [Google Scholar] [CrossRef]
- Maple, P.A.; Hamilton-Miller, J.M.; Brumfitt, W. World-wide antibiotic resistance in methicillin-resistant Staphylococcus aureus. Lancet 1989, 1, 537–540. [Google Scholar] [CrossRef]
- Barber, K.E.; Werth, B.J.; Rybak, M.J. The combination of ceftaroline plus daptomycin allows for therapeutic de-escalation and daptomycin sparing against MRSA. J. Antimicrob. Chemother. 2015, 70, 505–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werth, B.J.; Sakoulas, G.; Rose, W.E.; Pogliano, J.; Tewhey, R.; Rybak, M.J. Ceftaroline increases membrane binding and enhances the activity of daptomycin against daptomycin-nonsusceptible vancomycin-intermediate Staphylococcus aureus in a pharmacokinetic/pharmacodynamic model. Antimicrob. Agents Chemother. 2013, 57, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagihara, M.; Wiskirchen, D.E.; Kuti, J.L.; Nicolau, D.P. In vitro pharmacodynamics of vancomycin and cefazolin alone and in combination against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 202–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.B.; Yim, J.; Jahanbakhsh, S.; Sakoulas, G.; Rybak, M.J. Impact of cefazolin co-administration with vancomycin to reduce development of vancomycin-intermediate Staphylococcus aureus. Diagn Microbiol. Infect. Dis. 2018, 91, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhand, A.; Bayer, A.S.; Pogliano, J.; Yang, S.J.; Bolaris, M.; Nizet, V.; Wang, G.; Sakoulas, G. Use of antistaphylococcal beta-lactams to increase daptomycin activity in eradicating persistent bacteremia due to methicillin-resistant Staphylococcus aureus: Role of enhanced daptomycin binding. Clin. Infect. Dis. 2011, 53, 158–163. [Google Scholar] [CrossRef]
- Mehta, S.; Singh, C.; Plata, K.B.; Chanda, P.K.; Paul, A.; Riosa, S.; Rosato, R.R.; Rosato, A.E. beta-Lactams increase the antibacterial activity of daptomycin against clinical methicillin-resistant Staphylococcus aureus strains and prevent selection of daptomycin-resistant derivatives. Antimicrob. Agents Chemother. 2012, 56, 6192–6200. [Google Scholar] [CrossRef] [Green Version]
- Berti, A.D.; Sakoulas, G.; Nizet, V.; Tewhey, R.; Rose, W.E. beta-Lactam antibiotics targeting PBP1 selectively enhance daptomycin activity against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.; Stinear, T.P.; Grayson, M.L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: Resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Iwamoto, A.; Lian, J.Q.; Neoh, H.M.; Maruyama, T.; Horikawa, Y.; Hiramatsu, K. Novel mechanism of antibiotic resistance originating in vancomycin-intermediate Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Sieradzki, K.; Tomasz, A. Inhibition of cell wall turnover and autolysis by vancomycin in a highly vancomycin-resistant mutant of Staphylococcus aureus. J. Bacteriol. 1997, 179, 2557–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieradzki, K.; Tomasz, A. Suppression of beta-lactam antibiotic resistance in a methicillin-resistant Staphylococcus aureus through synergic action of early cell wall inhibitors and some other antibiotics. J. Antimicrob. Chemother. 1997, 39 (Suppl. A), 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, F.W.; Atoui, R.; Ben Ali, A.; Nguyen, J.C.; Ly, A.; Kitzis, M.D. False synergy between vancomycin and beta-lactams against glycopeptide-intermediate Staphylococcus aureus (GISA) caused by inappropriate testing methods. Clin. Microbiol. Infect. 2004, 10, 342–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, K.N.; Rybak, M.J. beta-Lactam Combinations with Vancomycin Show Synergistic Activity against Vancomycin-Susceptible Staphylococcus aureus, Vancomycin-Intermediate S. aureus (VISA), and Heterogeneous VISA. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chambers, H.F. Staphylococcus aureus with heterogeneous resistance to vancomycin: Epidemiology, clinical significance, and critical assessment of diagnostic methods. Antimicrob. Agents Chemother. 2003, 47, 3040–3045. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.Y.C.; Lye, D.C.; Yahav, D.; Sud, A.; Robinson, J.O.; Nelson, J.; Archuleta, S.; Roberts, M.A.; Cass, A.; Paterson, D.L.; et al. Effect of Vancomycin or Daptomycin With vs Without an Antistaphylococcal beta-Lactam on Mortality, Bacteremia, Relapse, or Treatment Failure in Patients With MRSA Bacteremia: A Randomized Clinical Trial. JAMA 2020, 323, 527–537. [Google Scholar] [CrossRef]
- Werth, B.J.; Jain, R.; Hahn, A.; Cummings, L.; Weaver, T.; Waalkes, A.; Sengupta, D.; Salipante, S.J.; Rakita, R.M.; Butler-Wu, S.M. Emergence of dalbavancin non-susceptible, vancomycin-intermediate Staphylococcus aureus (VISA) after treatment of MRSA central line-associated bloodstream infection with a dalbavancin- and vancomycin-containing regimen. Clin. Microbiol. Infect. 2018, 24, 429.e1–429.e5. [Google Scholar] [CrossRef] [Green Version]
- Steele, J.M.; Seabury, R.W.; Hale, C.M.; Mogle, B.T. Unsuccessful treatment of methicillin-resistant Staphylococcus aureus endocarditis with dalbavancin. J. Clin. Pharm. Ther. 2018, 43, 101–103. [Google Scholar] [CrossRef]
- Steed, M.E.; Werth, B.J.; Ireland, C.E.; Rybak, M.J. Evaluation of the novel combination of high-dose daptomycin plus trimethoprim-sulfamethoxazole against daptomycin-nonsusceptible methicillin-resistant Staphylococcus aureus using an in vitro pharmacokinetic/pharmacodynamic model of simulated endocardial vegetations. Antimicrob. Agents Chemother. 2012, 56, 5709–5714. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. M100-S25 Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Eigth Informational Supplement. 2018. Available online: https://clsi.org/media/1930/m100ed28 (accessed on 9 September 2019).
- Kebriaei, R.; Rice, S.A.; Stamper, K.C.; Rybak, M.J. Dalbavancin Alone and in Combination with Ceftaroline against Four Different Phenotypes of Staphylococcus aureus in a Simulated Pharmacodynamic/Pharmacokinetic Model. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Interscience. easySpiral: Technical Specifications; Interscience: Woburn, MA, USA, 2018. [Google Scholar]
- Food and Drug Administration. Dalbavancin: Package Insert Administration; Food and Drug Administration: Silver Spring, MD, USA, 2014.
- Leonard, S.N.; Rybak, M.J. Evaluation of vancomycin and daptomycin against methicillin-resistant Staphylococcus aureus and heterogeneously vancomycin-intermediate S. aureus in an in vitro pharmacokinetic/pharmacodynamic model with simulated endocardial vegetations. J. Antimicrob. Chemother. 2009, 63, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benvenuto, M.; Benziger, D.P.; Yankelev, S.; Vigliani, G. Pharmacokinetics and tolerability of daptomycin at doses up to 12 milligrams per kilogram of body weight once daily in healthy volunteers. Antimicrob. Agents Chemother. 2006, 50, 3245–3249. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.; Udy, A.A.; Wallis, S.C.; Jarrett, P.; Stuart, J.; Lassig-Smith, M.; Deans, R.; Roberts, M.S.; Taraporewalla, K.; Jenkins, J.; et al. Plasma and tissue pharmacokinetics of cefazolin in patients undergoing elective and semielective abdominal aortic aneurysm open repair surgery. Antimicrob. Agents Chemother. 2011, 55, 5238–5242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens-Lobenhoffer, J.; Kielstein, J.T.; Oye, C.; Bode-Boger, S.M. Validated high performance liquid chromatography-UV detection method for the determination of daptomycin in human plasma. J. Chromatogr. B Analyt Technol. Biomed Life Sci. 2008, 875, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Wootton, M.; Howe, R.A.; Hillman, R.; Walsh, T.R.; Bennett, P.M.; MacGowan, A.P. A modified population analysis profile (PAP) method to detect hetero-resistance to vancomycin in Staphylococcus aureus in a UK hospital. J. Antimicrob. Chemother. 2001, 47, 399–403. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |




{kind=link}
{kind=link}
{kind=link}
| N = 20 | DAL | DAL + CFZ | VAN | VAN + CFZ | DAP | DAP + CFZ | CFZ |
|---|---|---|---|---|---|---|---|
| MRSA | 0.016–0.063 | 0.002–0.008 | 0.5–2 | 0.063–0.25 | 0.063–1 | 0.0313–0.125 | 64–≥256 |
| hVISA | 0.016–0.125 | 0.004–0.008 | 1–2 | 0.25–0.5 | 0.063–1 | 0.063–0.25 | 64–>256 |
| VISA | 0.063–0.125 | 0.008 | 4 | 0.25–1 | 0.125–1 | 0.125–0.25 | >256 |
| DNS | 0.125–0.25 | 0.008 | 1–2 | 0.25–1 | 2–4 | 0.125–0.25 | >256 |
| DNS-VISA | 0.125–0.25 | 0.008 | 4 | 0.25–1 | 2–4 | 0.125 | >256 |
| Parameter | DAL | VAN | DAP | CFZ |
|---|---|---|---|---|
| fCmax (mg/L) | 30.1 ± 0.018 (30.1) | 34.9 ± 0.76 (36.0) | 13.95 ± 0.04 (14.1) | 27.55 ± 0.91 (26.0) |
| T1/2 (h) | 184 ± 0.01 (187.40) | 6.6 ± 0.03 (6.00) | 7.6 ± 0.07 (8.00) | 2.4 ± 0.04 (2.3) |
| AUC (mg × h/mL) or T > MIC (h) | 7255± 0.03 | 324. 66 ± 1.3 | 192.6 ± 0.09 | 0.00 |
| fAUC/MIC | 58,040 (D712) 50,040 (6913) | 81 (D712) 40 (6913) | 96 (D712) 49 (6913) | 234,082 (DAL+CFZ D712); 646 (VAN+CFZ; D712); 1541( DAP+CFZ; D712); 234,082 (DAL+CFZ; 6913) 323 (VAN+CFZ; 6913); 385 (DAP+CFZ; 6913) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul-Mutakabbir, J.C.; Kebriaei, R.; Stamper, K.C.; Sheikh, Z.; Maassen, P.T.; Lev, K.L.; Rybak, M.J. Dalbavancin, Vancomycin and Daptomycin Alone and in Combination with Cefazolin against Resistant Phenotypes of Staphylococcus aureus in a Pharmacokinetic/Pharmacodynamic Model. Antibiotics 2020, 9, 696. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100696
Abdul-Mutakabbir JC, Kebriaei R, Stamper KC, Sheikh Z, Maassen PT, Lev KL, Rybak MJ. Dalbavancin, Vancomycin and Daptomycin Alone and in Combination with Cefazolin against Resistant Phenotypes of Staphylococcus aureus in a Pharmacokinetic/Pharmacodynamic Model. Antibiotics. 2020; 9(10):696. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100696
Chicago/Turabian StyleAbdul-Mutakabbir, Jacinda C., Razieh Kebriaei, Kyle C. Stamper, Zain Sheikh, Philip T. Maassen, Katherine L. Lev, and Michael J. Rybak. 2020. "Dalbavancin, Vancomycin and Daptomycin Alone and in Combination with Cefazolin against Resistant Phenotypes of Staphylococcus aureus in a Pharmacokinetic/Pharmacodynamic Model" Antibiotics 9, no. 10: 696. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9100696