Towards the Identification of Antibiotic-Resistant Bacteria Causing Urinary Tract Infections Using Volatile Organic Compounds Analysis—A Pilot Study

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
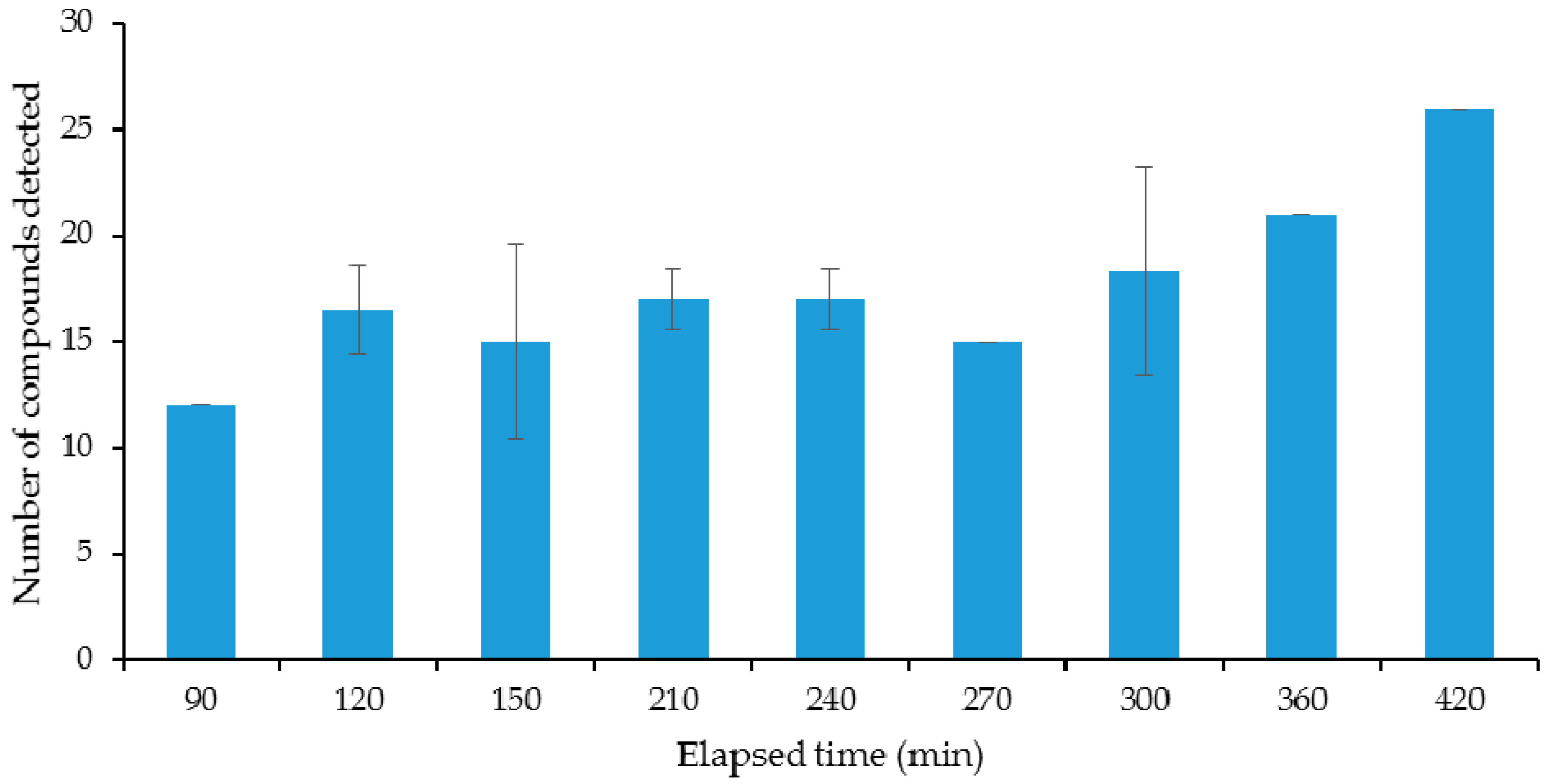
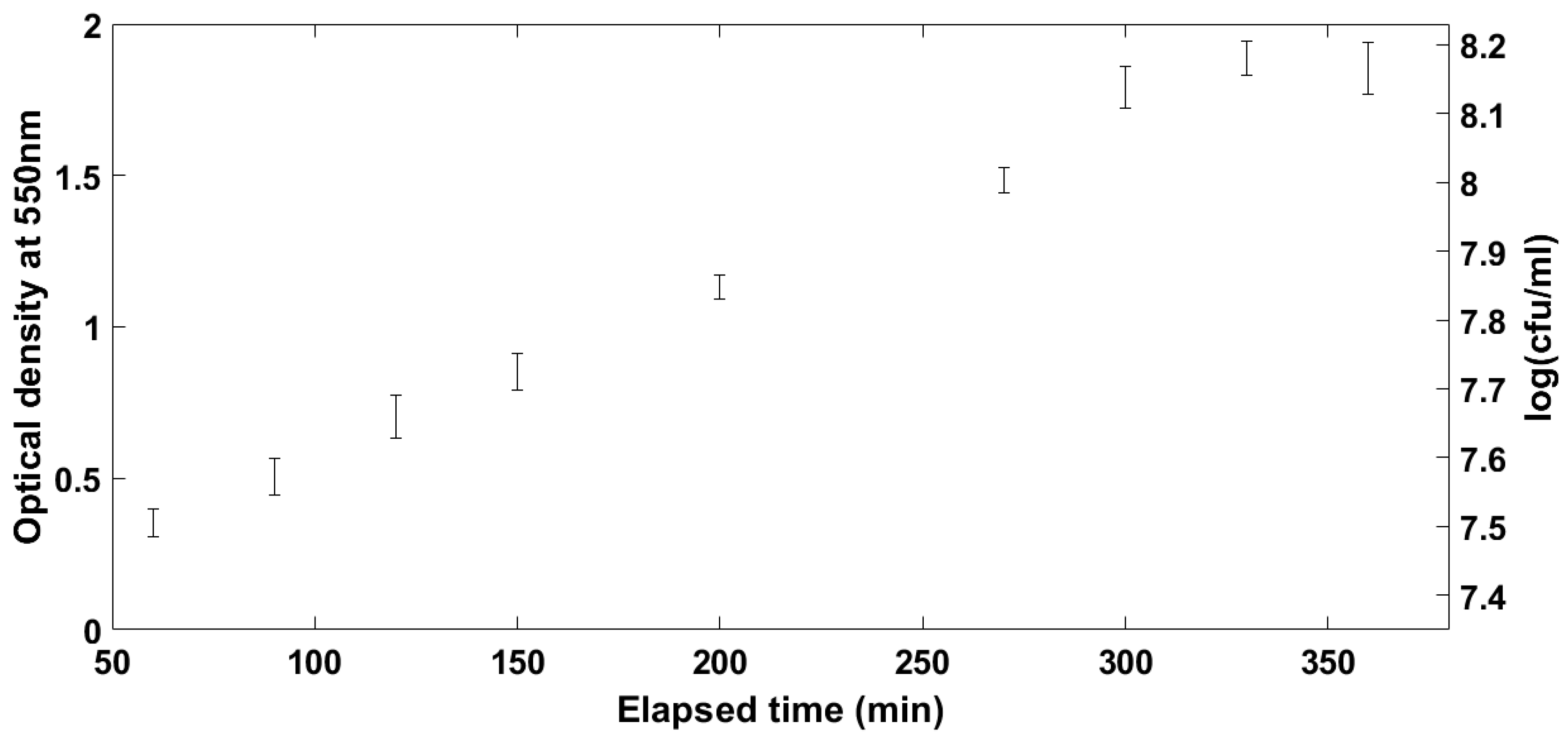
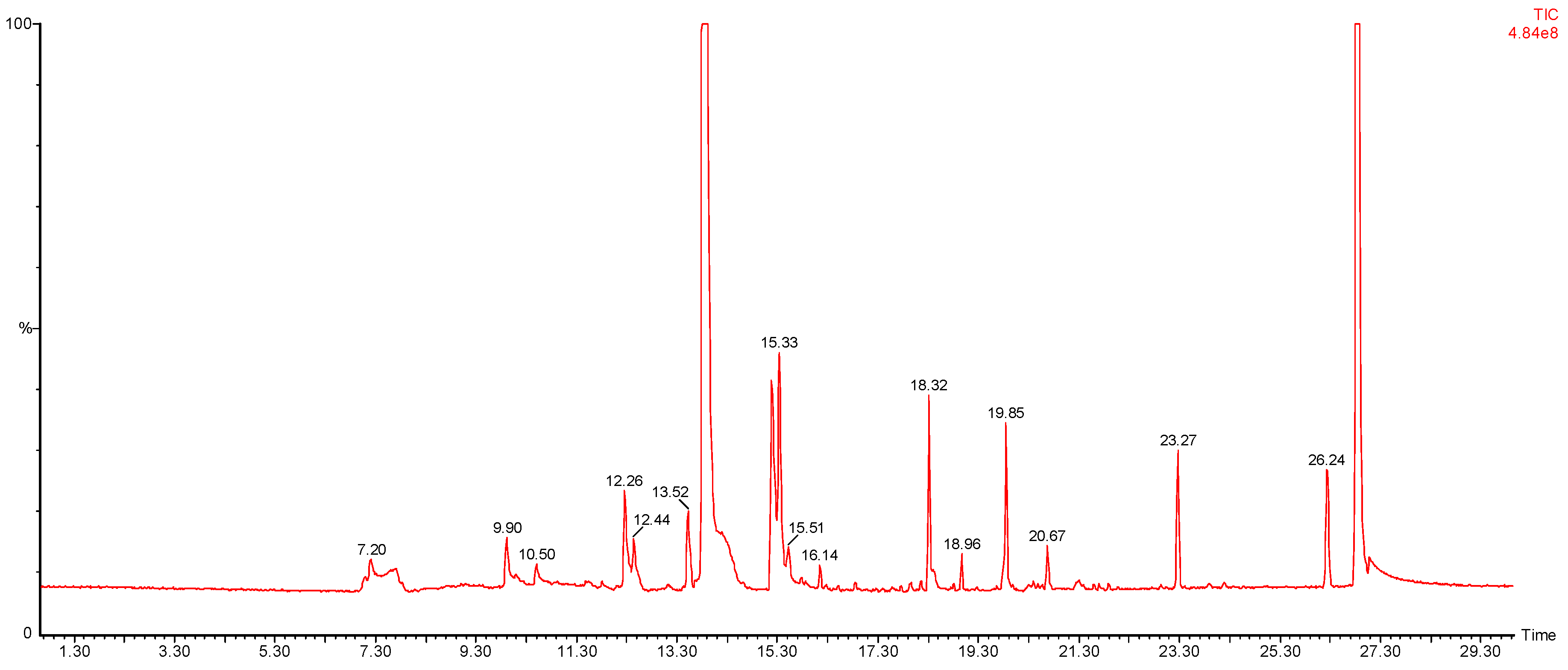
2.1. Selecting of the Optimal Microbiological Conditions
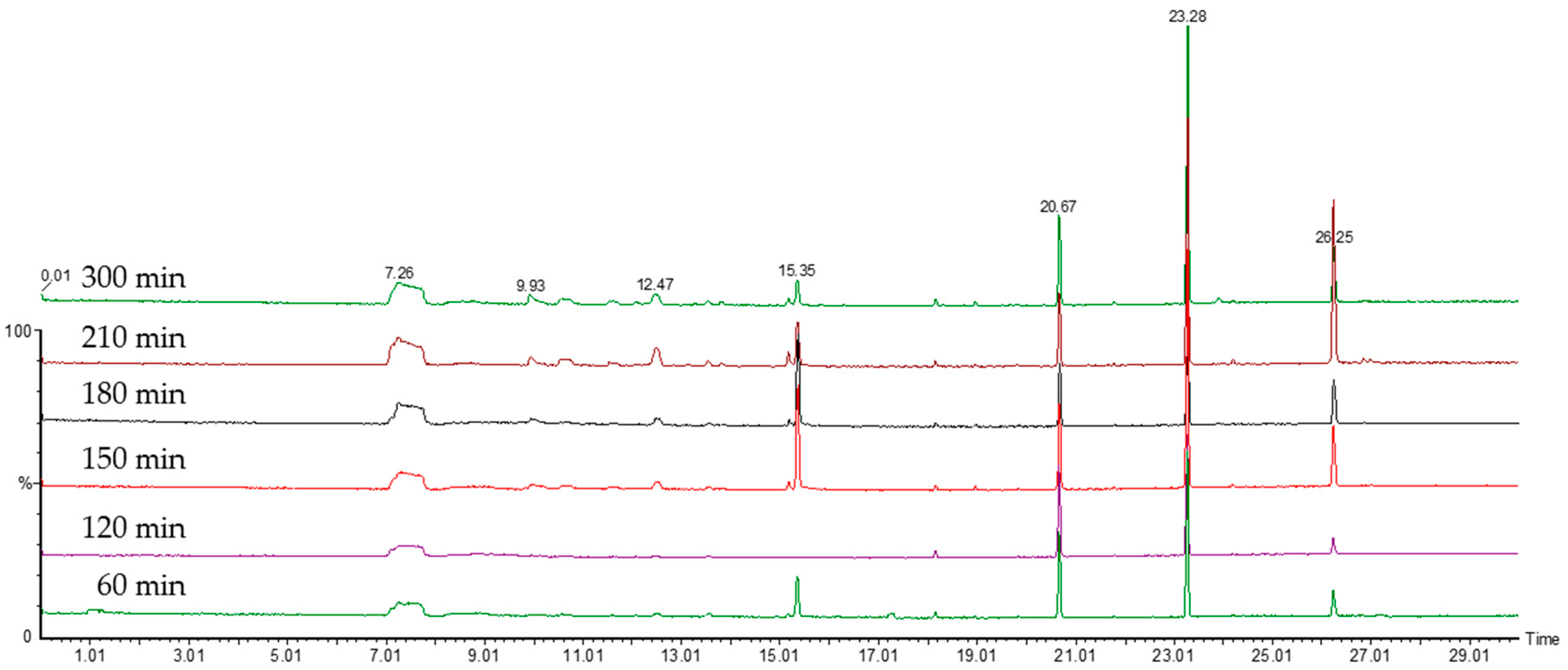
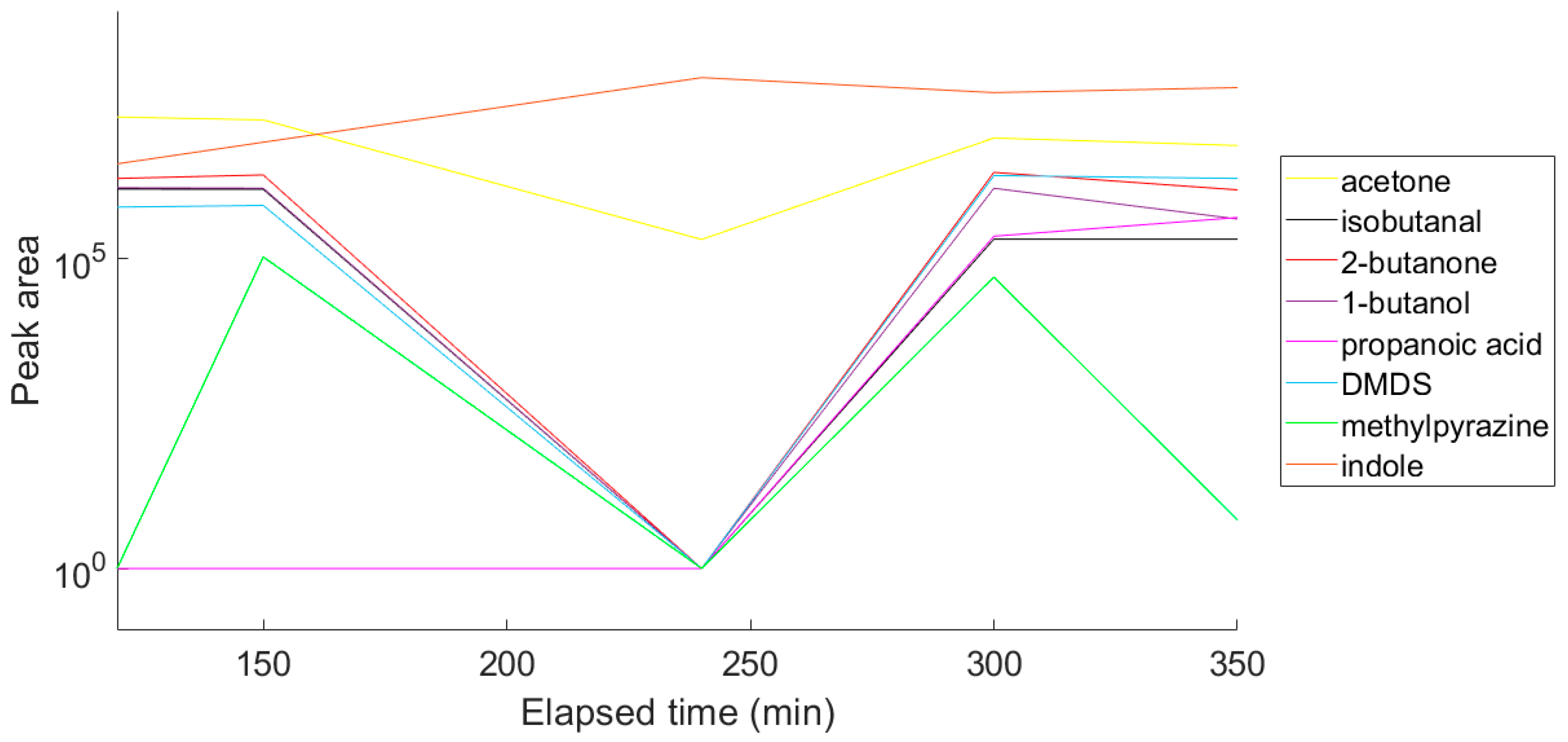
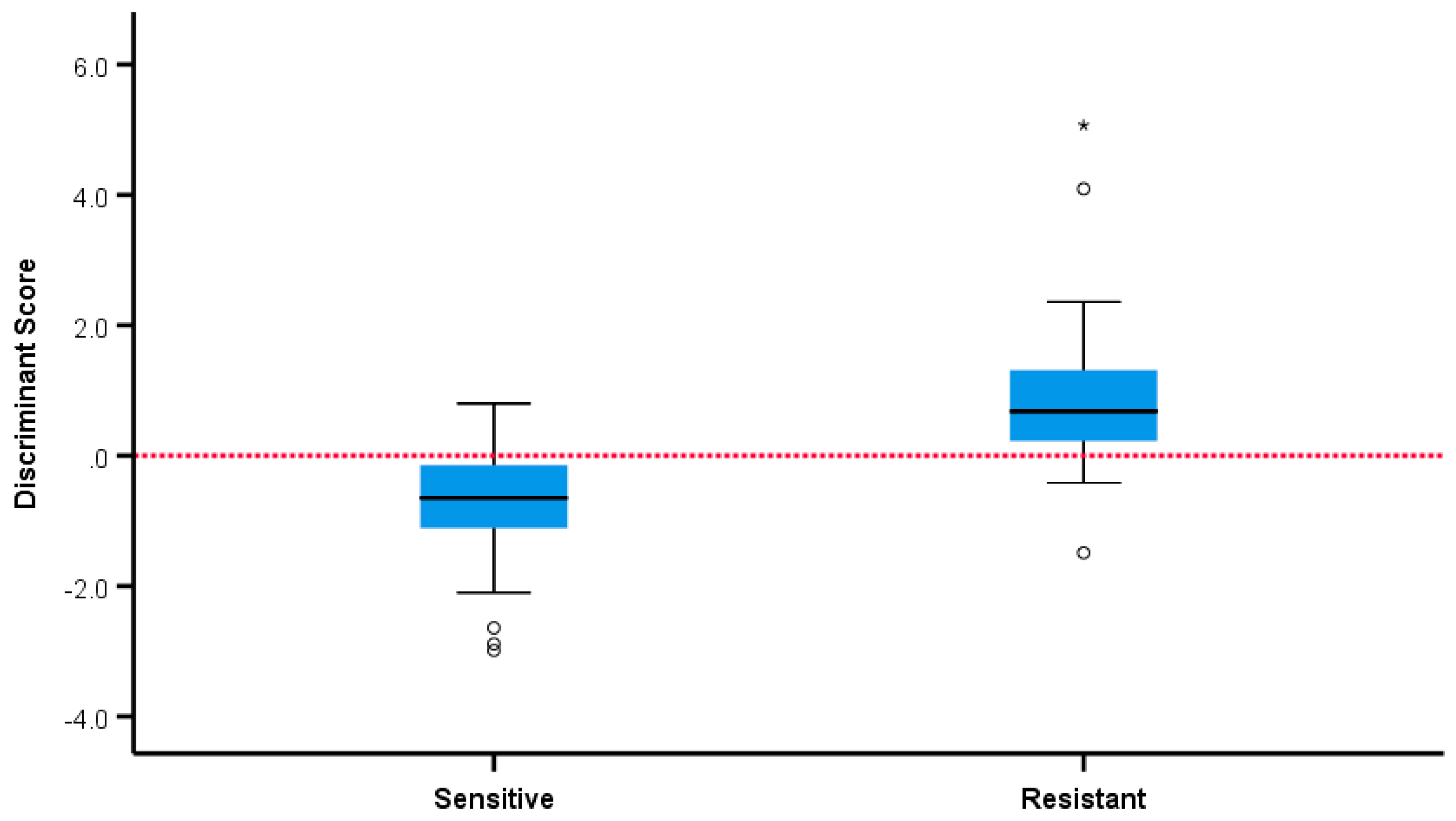
2.2. Comparison of VOC Profiles of Resistant and Susceptible Bacteria
3. Materials and Methods
3.1. Bacterial Cultures and Sampling
3.2. VOC Analysis
3.3. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tandogdu, Z.; Wagenlehner, F.M.E. Global epidemiology of urinary tract infections. Curr. Opin. Infect. Dis. 2016, 29, 73–79. [Google Scholar] [CrossRef]
- Tandogdu, Z.; Cek, M.; Wagenlehner, F.; Naber, K.; Tenke, P.; van Ostrum, E.; Johansen, T.B. Resistance patterns of nosocomial urinary tract infections in urology departments: 8-year results of the global prevalence of infections in urology study. World J. Urol. 2014, 32, 791–801. [Google Scholar] [CrossRef]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria able to Destroy Penicillin. Nature 1940, 146, 837. [Google Scholar] [CrossRef]
- Datta, N.; Kontomichalou, P. Penicillinase Synthesis Controlled By Infectious R Factors In Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Baño, J.; Navarro, M.D.; Romero, L.; Martínez-Martínez, L.; Muniain, M.A.; Perea, E.J.; Pérez-Cano, R.; Pascual, A. Epidemiology and clinical features of infections caused by extended-spectrum beta-lactamase-producing Escherichia coli in nonhospitalized patients. J. Clin. Microbiol. 2004, 42, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Canto, R. The CTX-M b -lactamase pandemic’ n and Teresa M Coque. Curr. Opin. Microbiol. 2006, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Guglielmo, B.J. Diagnosis and Treatment of Extended-Spectrum and AmpC. SAGE J. 2007, 41, 1427–1435. [Google Scholar] [CrossRef]
- Bergeron, M.G.; Ouellette, M. Preventing antibiotic resistance through rapid genotypic identification of bacteria and of their antibiotic resistance genes in the clinical microbiology laboratory. J. Clin. Microbiol. 1998, 36, 2169–2172. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Call, D.R.; Bakko, M.K.; Krug, M.J.; Roberts, M.C. Identifying Antimicrobial Resistance Genes with DNA Microarrays. Antimicrob. Agents Chemother. 2003, 47, 3290–3295. [Google Scholar] [CrossRef] [Green Version]
- Hrabák, J.; Walková, R.; Študentová, V.; Chudáčková, E.; Bergerová, T. Carbapenemase activity detection by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2011, 49, 3222–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostrzewa, M.; Sparbier, K.; Maier, T.; Schubert, S. MALDI-TOF MS: An upcoming tool for rapid detection of antibiotic resistance in microorganisms. Proteom. -Clin. Appl. 2013, 7, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Sparbier, K.; Schubert, S.; Kostrzewa, M. MBT-ASTRA: A suitable tool for fast antibiotic susceptibility testing? Methods 2016, 104, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; de Lacy Costello, B.; Hewett, K.; Smart, A.; Ratcliffe, N. From fast identification to resistance testing: Volatile compound profiling as a novel diagnostic tool for detection of antibiotic susceptibility. TrAC-Trends Anal. Chem. 2019, 115, 1–12. [Google Scholar] [CrossRef]
- Wiesner, K.; Jaremek, M.; Pohle, R.; Von Sicard, O.; Stuetz, E. Monitoring of bacterial growth and rapid evaluation of antibiotic susceptibility by headspace gas analysis. In Procedia Engineering; Elsevier: Amsterdam, The Netherlands, 2014; Volume 87, pp. 332–335. [Google Scholar]
- Smart, A.; de Lacy Costello, B.; White, P.; Avison, M.; Batty, C.; Turner, C.; Persad, R.; Ratcliffe, N. Sniffing Out Resistance – Rapid Identification of Urinary Tract Infection-Causing Bacteria and their Antibiotic Susceptibility Using Volatile Metabolite Profiles. J. Pharm. Biomed. Anal. 2019. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Panigrahi, S. Solid-Phase Microextraction (SPME) Techniques for Quality Characterization of Food Products: A Review. Food Bioprocess Technol. 2011, 4, 1–26. [Google Scholar] [CrossRef]
- Cavalli, J.F.; Fernandez, X.; Lizzani-Cuvelier, L.; Loiseau, A.M. Comparison of Static Headspace, Headspace Solid Phase Microextraction, Headspace Sorptive Extraction, and Direct Thermal Desorption Techniques on Chemical Composition of French Olive Oils. J. Agric. Food Chem. 2003, 51, 7709–7716. [Google Scholar] [CrossRef]
- Jeleń, H.H.; Wlazły, K.; Wa̧sowicz, E.; Kamiński, E. Solid-Phase Microextraction for the Analysis of Some Alcohols and Esters in Beer: Comparison with Static Headspace Method. J. Agric. Food Chem. 1998, 46, 1469–1473. [Google Scholar] [CrossRef]
- Paolini, J.; Leandri, C.; Desjobert, J.M.; Barboni, T.; Costa, J. Comparison of liquid-liquid extraction with headspace methods for the characterization of volatile fractions of commercial hydrolats from typically Mediterranean species. J. Chromatogr. A 2008, 1193, 37–49. [Google Scholar] [CrossRef]
- Nunez-montiel, O.L.; Thompson, F.S.; Dowell, V.R. Norleucine-Tyrosine Broth for Rapid Identification of Clostridium difficile by Gas-Liquid Chromatography. J Clin Microbiol. 1983, 17, 382–385. [Google Scholar] [CrossRef] [Green Version]
- Hiele, M.; Ghoos, Y.; Rutgeerts, P.; Vantrappen, G.; Schoorens, D. Influence of Nutritional Substrates on the Formation of Volatiles by the Fecal Flora. Gastroenterology 1991, 1597–1602. [Google Scholar] [CrossRef]
- Zareian, M.; Silcock, P.; Bremer, P. Effect of medium compositions on microbially mediated volatile organic compounds release profile. J. Appl. Microbiol. 2018, 125, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Scholpp, J.; Schubert, J.K.; Miekisch, W.; Geiger, K. Breath markers and soluble lipid peroxidation markers in critically ill patients. Clin. Chem. Lab. Med. 2002, 40, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, W.; Ruzsanyi, V.; Mochalski, P.; Filipiak, A.; Bajtarevic, A.; Ager, C.; Denz, H.; Hilbe, W.; Jamnig, H.; Hackl, M.; et al. Dependence of exhaled breath composition on exogenous factors, smoking habits and exposure to air pollutants. J. Breath Res. 2012, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.H.M.P. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2013, 486, 207–214. [Google Scholar] [CrossRef]
- Rolfe, M.D.; Rice, C.J.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.D.S.; Alston, M.; Stringer, M.F.; Betts, R.P.; Baranyi, J.; et al. Lag Phase Is a Distinct Growth Phase That Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. J. Bacteriol. 2012, 194, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Harder, W.; Dijkhuizen, L. Strategies of mixed substrate utilization in microorganisms. Philisophical Trans. R. Soc. London B 1982, 480, 459–480. [Google Scholar]
- Thorn, R.M.S.; Greenman, J. Microbial volatile compounds in health and disease conditions. J. Breath Res. 2012, 6. [Google Scholar] [CrossRef]
- Magasanik, B. Catabolite Repression. Cold Spring Harb. Symp. Quant. Biol. 1961, 26, 249–256. [Google Scholar] [CrossRef]
- Baev, M.V.; Baev, D.; Jancso, A.; Campbell, J.W. Growth of Escherichia coli MG1655 on LB medium: Determining metabolic strategy with transcriptional microarrays. Appl. Microbiol. Biotechnol. 2006, 71, 323–328. [Google Scholar] [CrossRef]
- Zhu, J.; Bean, H.D.; Kuo, Y.M.; Hill, J.E. Fast detection of volatile organic compounds from bacterial cultures by secondary electrospray ionization-mass spectrometry. J. Clin. Microbiol. 2010, 48, 4426–4431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Tang, J.; Shi, H.; Tang, C.; Zhang, R. Characteristics of volatile organic compounds produced from five pathogenic bacteria by headspace-solid phase micro-extraction/gas chromatography-mass spectrometry. J. Basic Microbiol. 2017, 57, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Bos, L.D.J.; Sterk, P.J.; Schultz, M.J. Volatile Metabolites of Pathogens: A Systematic Review. PLoS Pathog. 2013, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakkarach, A.; Foo, H.L.; Song, A.A.-L.; Nitisinprasert, S.; Withayagiat, U. Promising discovery of beneficial Escherichia coli in the human gut. 3 Biotech 2020, 10, 296. [Google Scholar] [CrossRef]
- Findlay, J.; Gould, V.C.; North, P.; Bowker, K.E.; Williams, M.O.; MacGowan, A.P.; Avison, M.B. Characterization of cefotaxime-resistant urinary Escherichia coli from primary care in South-West England 2017–18. J. Antimicrob. Chemother. 2020, 75, 65–71. [Google Scholar] [CrossRef]
- Zellner, B.D.; Bicchi, C.; Dugo, P.; Rubiolo, P.; Dugo, G.; Mondello, L. Linear retention indices in gas chromatographic analysis: A review. Flavour Fragr. J. 2008, 23, 297–314. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time | Tentatively Identified VOCs | RI 1 |
|---|---|---|
| 6.99 | unknown 1 | 419 |
| 7.14 | unknown 2 | 428 |
| 9.84 | Ethanol | 590 |
| 10.44 | Acetone | 626 |
| 10.86 | unknown 3 | 652 |
| 11.70 | Propanol | 702 |
| 12.01 | unknown 4 | 721 |
| 12.36 | 2-butanone | 742 |
| 13.04 | Isobutanol | 782 |
| 13.17 | unknown 5 | 790 |
| 13.34 | unknown 6 | 800 |
| 13.44 | Isopentanal | 806 |
| 13.77 | 1-butanol | 826 |
| 14.06 | unknown 7 | 844 |
| 14.15 | Dimethylfuran | 849 |
| 14.67 | unknown 8 | 880 |
| 14.90 | propanoic acid | 894 |
| 15.12 | 1-pentanol | 907 |
| 15.24 | DMDS | 914 |
| 15.47 | Toluene | 928 |
| 15.71 | unknown 9 | 943 |
| 16.19 | Dimethylheptene | 971 |
| 16.44 | Octane | 986 |
| 18.24 | Methylpyrazine | 1094 |
| 18.90 | 2,2,4,6,6-pentamethylheptane | 1134 |
| 19.79 | Benzaldehyde | 1187 |
| 26.76 | Indole | 1606 |
| Retention Time (mins) | Tentatively Identified VOCs | RI 1 | p-value |
|---|---|---|---|
| 9.91 | ethanol | 595 | 0.112 |
| 10.09 | unknown 3 | 605 | 0.740 |
| 10.5 | acetone | 630 | 0.501 |
| 10.65 | dimethyl sulfide | 639 | 0.756 |
| 11.56 | isobutanal | 694 | 0.892 |
| 11.64 | unknown 10 | 698 | 0.193 |
| 11.8 | propanol | 708 | 0.650 |
| 12.27 | butanal | 736 | 0.057 |
| 12.43 | 2-butanone | 746 | 0.851 |
| 12.86 | unknown 11 | 772 | 0.861 |
| 13.13 | isobutanol | 788 | 0.942 |
| 13.5 | isopentanal | 810 | 0.052 |
| 13.65 | methylbutanal | 819 | 0.760 |
| 13.71 | pentanone | 823 | 0.413 |
| 13.8 | butanol | 828 | 0.647 |
| 14.61 | unknown 12 | 877 | 0.063 |
| 14.76 | unknown 8 | 886 | 0.228 |
| 15.15 | pentanol | 909 | 0.124 |
| 15.33 | DMDS | 920 | 0.767 |
| 15.51 | toluene | 931 | 0.957 |
| 15.77 | unknown 9 | 946 | 0.759 |
| 16.13 | butyl acetate | 968 | 0.117 |
| 16.25 | unknown 13 | 975 | 0.317 |
| 16.43 | methylbutenal | 986 | 0.917 |
| 16.52 | unknown 14 | 991 | 0.507 |
| 16.68 | butanoic acid | 1001 | 0.040 |
| 16.83 | methylpyrazine | 1010 | 0.660 |
| 17.03 | unknown 15 | 1022 | 0.104 |
| 17.12 | dimethylheptane | 1027 | 0.314 |
| 17.19 | 1-methoxy-2-propyl acetate | 1031 | 0.507 |
| 17.27 | ethylbenzene | 1036 | 0.410 |
| 17.38 | x-xylene2 | 1043 | 0.925 |
| 17.55 | allyl butyrate | 1053 | 0.065 |
| 17.64 | unknown 16 | 1058 | 0.246 |
| 17.79 | butyl propionate | 1067 | 0.069 |
| 17.87 | unknown 17 | 1072 | 0.826 |
| 17.93 | 2-heptanone | 1076 | 0.754 |
| 18.3 | 2,5-dimethylpyrazine | 1098 | 0.713 |
| 18.38 | unknown 18 | 1103 | 0.407 |
| 18.5 | unknown 19 | 1110 | 0.011 |
| 18.77 | methyl-heptanone | 1126 | 0.069 |
| 18.8 | unknown 20 | 1128 | 0.513 |
| 18.96 | 2,3,4,6,6-pentamethylheptane | 1138 | 0.101 |
| 19.04 | unknown 21 | 1142 | 0.304 |
| 19.11 | unknown 22 | 1147 | 0.054 |
| 19.2 | unknown 23 | 1152 | 0.025 |
| 19.26 | butyl isobutyrate | 1156 | 0.067 |
| 19.47 | unknown 24 | 1168 | 0.030 |
| 19.62 | unknown 25 | 1177 | 0.478 |
| 19.82 | dimethyl trisulfide | 1189 | 0.943 |
| 19.83 | benzaldehyde | 1190 | 0.203 |
| 19.92 | D-limonene | 1195 | 0.943 |
| 19.98 | unknown 26 | 1199 | 0.224 |
| 20.24 | 2-ethyl-hexanol | 1214 | 0.155 |
| 20.4 | unknown 27 | 1224 | 0.003 |
| 20.48 | unknown 28 | 1229 | 0.914 |
| 20.52 | unknown 29 | 1231 | 0.814 |
| 20.57 | unknown 30 | 1234 | 0.781 |
| 20.79 | unknown 31 | 1247 | 0.200 |
| 20.91 | butylglycol acetate | 1255 | 0.098 |
| 20.94 | unknown 32 | 1256 | 0.199 |
| 21.00 | unknown 33 | 1260 | 0.264 |
| 21.21 | benzyl alcohol | 1273 | 0.536 |
| 21.29 | benzyl alcohol | 1277 | 0.088 |
| 21.39 | unknown 34 | 1283 | 0.399 |
| 21.59 | acetophenone | 1295 | 0.211 |
| 21.71 | unknown 35 | 1303 | 0.580 |
| 21.78 | unknown 36 | 1307 | 0.878 |
| 21.89 | unknown 37 | 1313 | 0.161 |
| 22.07 | cresol | 1324 | 0.197 |
| 22.79 | unknown 38 | 1367 | 0.202 |
| 23.01 | unknown 39 | 1381 | 0.837 |
| 23.39 | unknown 40 | 1403 | 0.259 |
| 23.46 | unknown 41 | 1408 | 0.822 |
| 23.9 | pentyllfuran | 1434 | 0.873 |
| 24.02 | unknown 42 | 1441 | 0.461 |
| 24.18 | unknown 43 | 1451 | 0.369 |
| 24.36 | 2-dodecanone | 1462 | 0.005 |
| 24.57 | unknown 44 | 1474 | 0.363 |
| 25.07 | unknown 45 | 1504 | 0.045 |
| 26.48 | unknown 46 | 1589 | 0.550 |
| 26.87 | indole | 1612 | 0.156 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hewett, K.; Drabińska, N.; White, P.; Avison, M.B.; Persad, R.; Ratcliffe, N.; Costello, B.d.L. Towards the Identification of Antibiotic-Resistant Bacteria Causing Urinary Tract Infections Using Volatile Organic Compounds Analysis—A Pilot Study. Antibiotics 2020, 9, 797. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9110797
Hewett K, Drabińska N, White P, Avison MB, Persad R, Ratcliffe N, Costello BdL. Towards the Identification of Antibiotic-Resistant Bacteria Causing Urinary Tract Infections Using Volatile Organic Compounds Analysis—A Pilot Study. Antibiotics. 2020; 9(11):797. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9110797
Chicago/Turabian StyleHewett, Keith, Natalia Drabińska, Paul White, Matthew B. Avison, Raj Persad, Norman Ratcliffe, and Ben de Lacy Costello. 2020. "Towards the Identification of Antibiotic-Resistant Bacteria Causing Urinary Tract Infections Using Volatile Organic Compounds Analysis—A Pilot Study" Antibiotics 9, no. 11: 797. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9110797