Phenotypic and Genotypic Features of Klebsiella pneumoniae Harboring Carbapenemases in Egypt: OXA-48-Like Carbapenemases as an Investigated Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
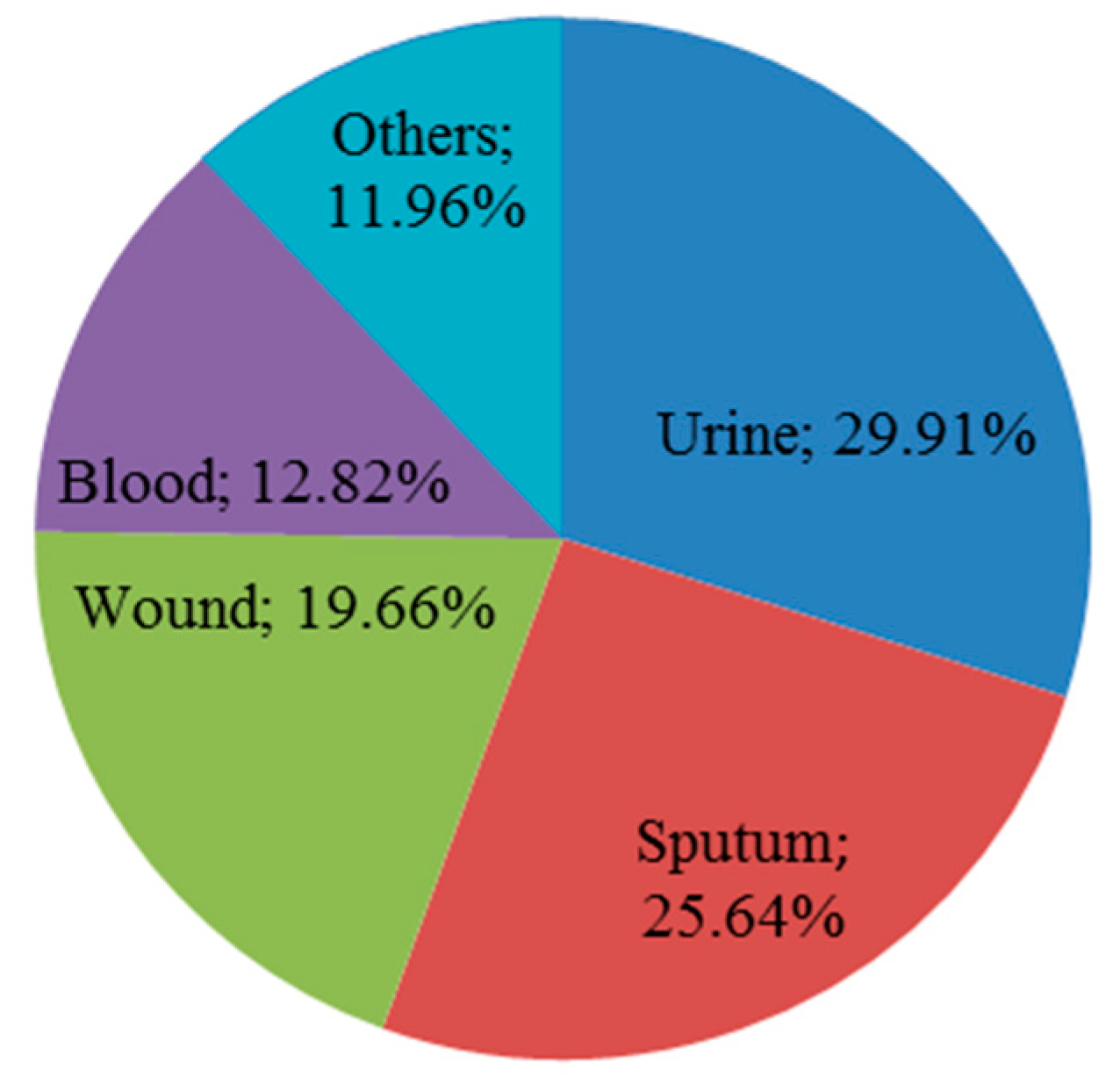
2.1. Distribution and Identification of K. pneumoniae Isolates
2.2. Isolate Antimicrobial Susceptibility Pattern
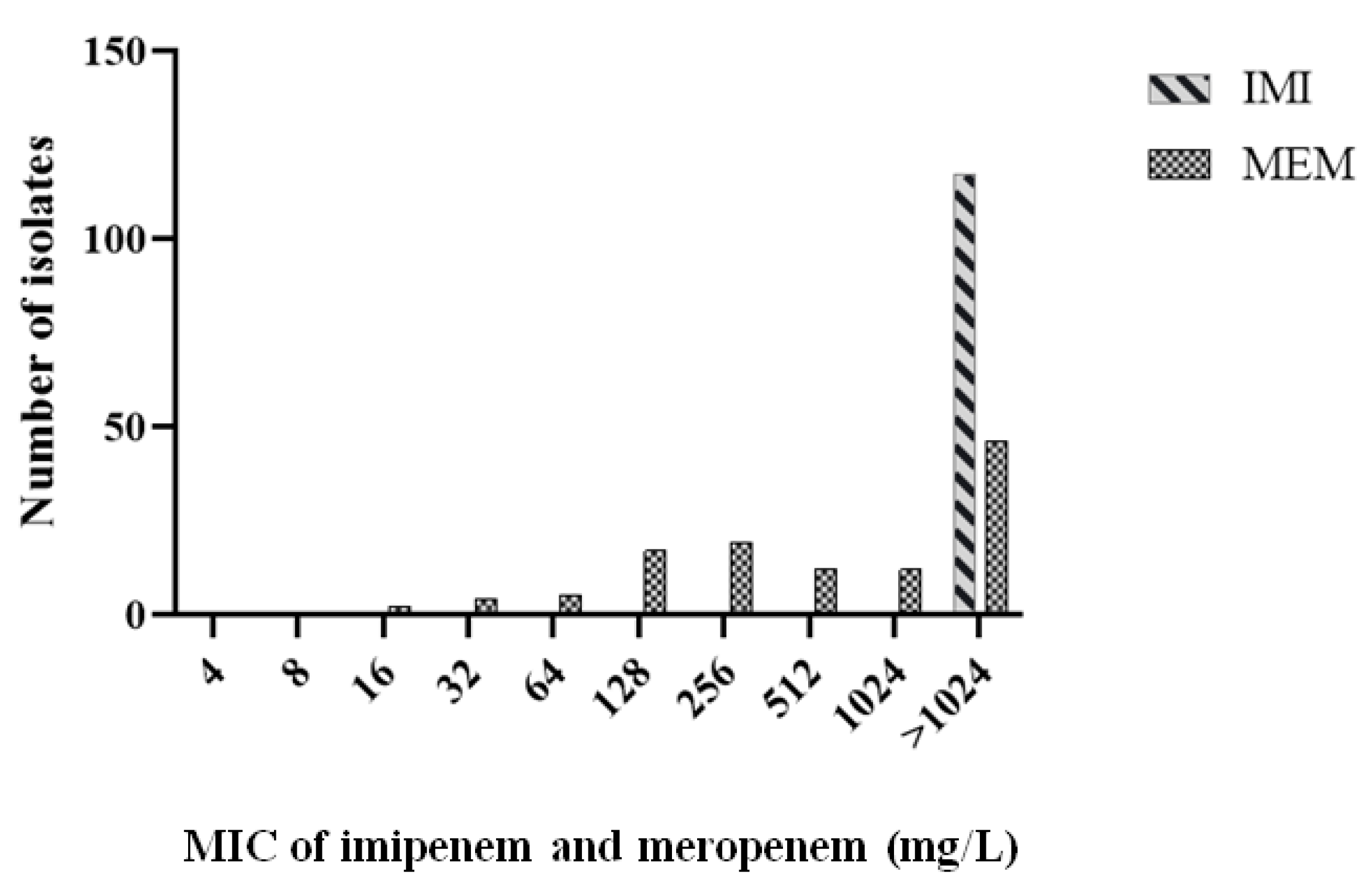
2.3. Phenotypic and Genotypic Characterization of Carbapenem-Resistant Isolates
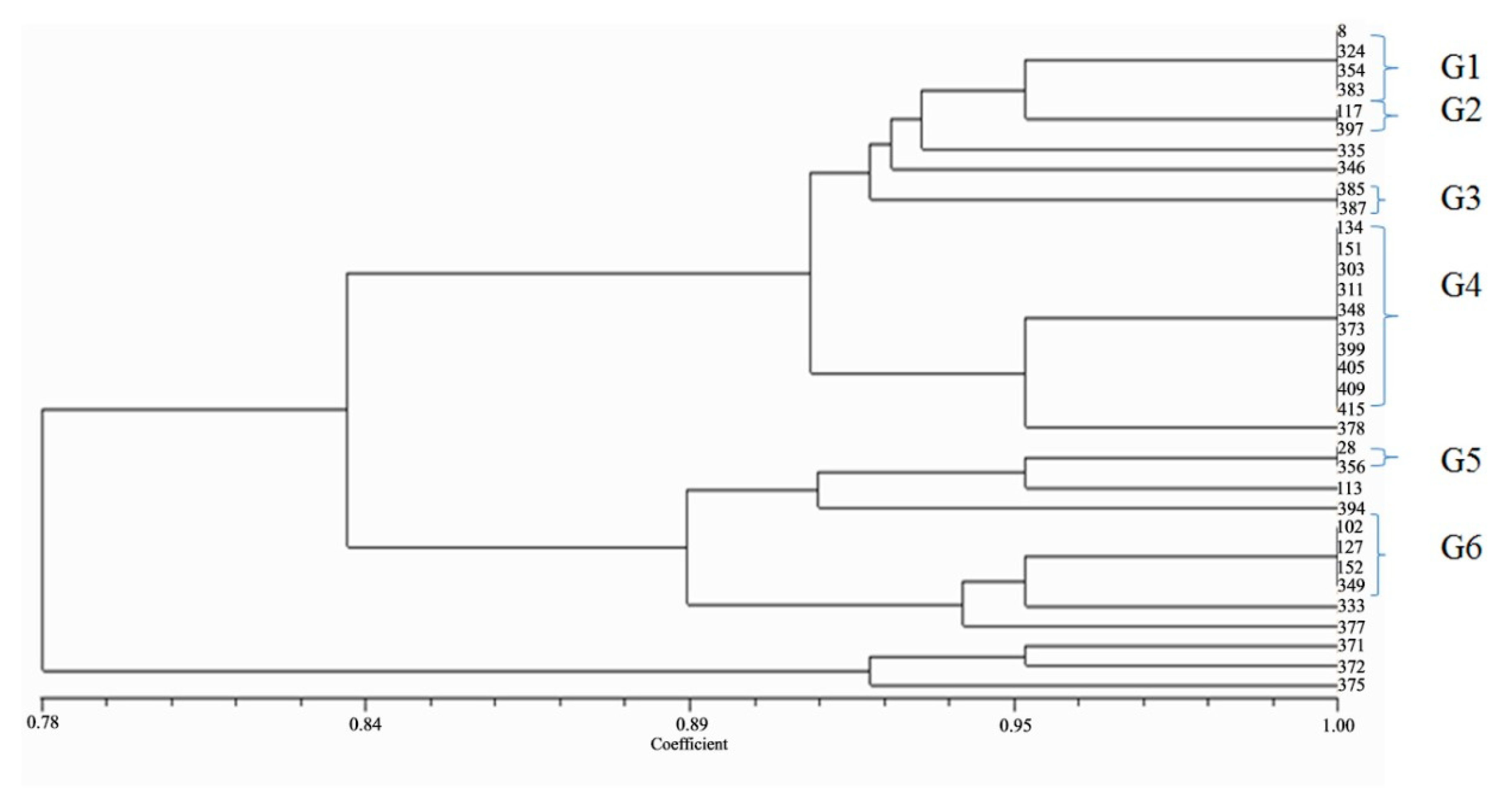
2.4. Genotyping of Isolates Harboring blaOXA-48-like Genes
2.5. Transferability of blaOXA-48 Gene
2.6. Assessment of Biofilm Formation and Detection of Adhesion Encoding Genes in Carbapenem-Resistant K. pneumoniae Isolates
2.7. The Correlation between Non-Susceptibility Pattern and Biofilm Formation Category of Carbapenem-Resistant K. pneumoniae Isolates
3. Discussion
4. Materials and Methods
4.1. Isolates Collection and Confirmation
4.2. Antimicrobial Susceptibility Testing
4.3. Phenotypic Confirmation of Carbapenemases Production
4.4. Molecular Detection of Carbapenemases Encoding Genes
4.5. Genotyping of Isolates Harboring blaOXA-48-like Genes
4.5.1. Enterobacterial Repetitive Intergenic Consensus (ERIC-PCR)
4.5.2. Plasmid Profile Analysis
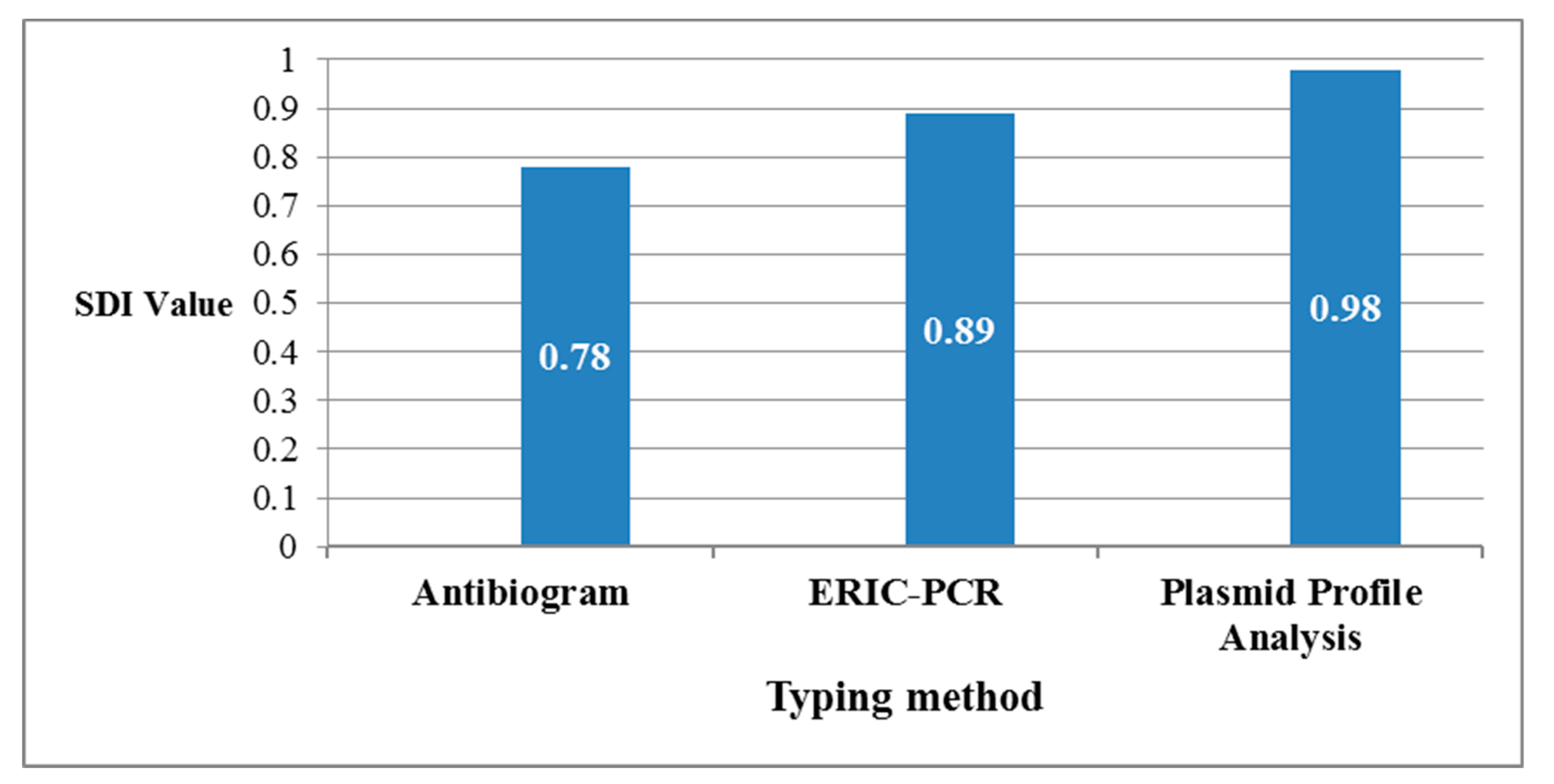
4.6. Calculation of the Discriminatory Power of Different Applied Typing Methods
4.7. Genetic Transfer of blaOXA-48 Gene by Transformation
4.8. Quantitative Biofilm Formation Assay for the Carbapenem-Resistant Isolates by Microtitre Plate Method
4.9. Molecular Detection of Type 1 (fimH-1) and Type 3 (mrkD) Adhesion Encoding Genes
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- O’Neill, J.I.M. Antimicrobial resistance: Tackling a crisis for the health and wealth of nations. Rev. Antimicrob. Resist. 2014, 20, 1–16. [Google Scholar]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014; pp. 1–256. Available online: http://apps.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf?ua=1 (accessed on 18 June 2020).
- Centers for Disease Control and Prevention. About Antimicrobial Resistance. 2018. Available online: https://www.cdc.gov/drugresistance/about.html (accessed on 18 June 2020).
- World Bank. Drug-Resistant Infections: A Threat to Our Economic Future; World Bank: Washington, DC, USA, 2017; Available online: http://www.worldbank.org/en/topic/health/publication/drug-resistant-infections-a-threat-to-our-economic-future (accessed on 20 June 2020).
- Rice, L.B. Progress and Challenges in Implementing the Research on ESKAPE Pathogens. Infect. Control. Hosp. Epidemiol. 2010, 31, S7–S10. [Google Scholar] [CrossRef]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 22 June 2020).
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Halat, D.H.; Moubareck, C.A. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef]
- Bratu, S.; Landman, D.; Haag, R.; Recco, R.; Eramo, A.; Alam, M.; Quale, J. Rapid Spread of Carbapenem-Resistant Klebsiella pneumoniae in New York City. Arch. Intern. Med. 2005, 165, 1430–1435. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.H.; Hu, Z.Q. Acquired metallo-β-lactamases and their genetic association with class 1 integrons and ISCRelements in Gram-negative bacteria. Futur. Microbiol. 2015, 10, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Genet. 2020, 18, 344–359. [Google Scholar] [CrossRef]
- Pitout, J.; Peirano, G.; Kock, M.M.; Strydom, K.A.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, 1–48. [Google Scholar] [CrossRef]
- Potron, A.; Poirel, L.; Nordmann, P. Derepressed Transfer Properties Leading to the Efficient Spread of the Plasmid Encoding Carbapenemase OXA-48. Antimicrob. Agents Chemother. 2013, 58, 467–471. [Google Scholar] [CrossRef] [Green Version]
- Van Belkum, A.; Tassios, P.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. 2007, 13, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Vert, M.; Doi, Y.; Hellwich, K.H.; Hess, M.; Hodge, P.E.; Kubisa, P.; Rinaudo, M.; Schué, F. Terminology for biorelated polymers and applications (IUPAC Recommendations 2012). Pure Appl. Chem. 2012, 84, 377–410. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic Resistance Related to Biofilm Formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcántar-Curiel, M.D.; Blackburn, D.; Saldaña, Z.; Gayosso-Vázquez, C.; Iovine, N.M.; De La Cruz, M.A.; Girón, J.A. Multi-functional analysis ofKlebsiella pneumoniaefimbrial types in adherence and biofilm formation. Virulence 2013, 4, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.J.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Manenzhe, R.I.; Zar, H.J.; Nicol, M.P.; Kaba, M. The spread of carbapenemase-producing bacteria in Africa: A systematic review. J. Antimicrob. Chemother. 2015, 70, 23–40. [Google Scholar] [CrossRef] [Green Version]
- Sekyere, J.O.; Govinden, U.; Essack, S. The Molecular Epidemiology and Genetic Environment of Carbapenemases Detected in Africa. Microb. Drug Resist. 2016, 22, 59–68. [Google Scholar] [CrossRef]
- Touati, A.; Mairi, A. Epidemiology of Carbapenemase-producing Enterobacterales in the Middle East: A systematic review. Expert Rev. Anti-Infect. Ther. 2020, 18, 241–250. [Google Scholar] [CrossRef]
- Yan, Z.; Zhou, Y.; Du, M.; Bai, Y.; Liu, B.; Gong, M.; Song, H.; Tong, Y.; Liu, Y. Prospective investigation of carbapenem-resistant Klebsiella pneumonia transmission among the staff, environment and patients in five major intensive care units, Beijing. J. Hosp. Infect. 2019, 101, 150–157. [Google Scholar] [CrossRef]
- Lerminiaux, N.A.; Cameron, A.D. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Skalova, A.; Chudejova, K.; Rotova, V.; Medvecky, M.; Studentova, V.; Chudackova, E.; Lavicka, P.; Bergerova, T.; Jakubu, V.; Zemlickova, H.; et al. Molecular Characterization of OXA-48-Like-Producing Enterobacteriaceae in the Czech Republic and Evidence for Horizontal Transfer of pOXA-48-Like Plasmids. Antimicrob. Agents Chemother. 2016, 61. [Google Scholar] [CrossRef] [Green Version]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance—The Need for Global Solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- Zakaa El-din, M.; Samy, F.; Mohamed, A.; Hamdy, F.; Yasser, S.; Ehab, M. Egyptian community pharmacists’ attitudes and practices towards antibiotic dispensing and antibiotic resistance; a cross-sectional survey in Greater Cairo. Curr. Med. Res. Opin. 2018, 35, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Argente, M.; Miró, E.; Martí, C.; Vilamala, A.; Alonso-Tarrés, C.; Ballester, F.; Calderon, A.; Galles, C.; Gasós, A.; Mirelis, B.; et al. Molecular characterization of OXA-48 carbapenemase-producing Klebsiella pneumoniae strains after a carbapenem resistance increase in Catalonia. Enferm. Infecc. Microbiol. Clínica 2019, 37, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Karampatakis, T.; Geladari, A.; Politi, L.; Antachopoulos, C.; Iosifidis, E.; Tsiatsiou, O.; Karyoti, A.; Papanikolaou, V.; Tsakris, A.; Roilides, E. Cluster-distinguishing genotypic and phenotypic diversity of carbapenem-resistant Gram-negative bacteria in solid-organ transplantation patients: A comparative study. J. Med. Microbiol. 2017, 66, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Azimi, L.; Talebi, M.; Owlia, P.; Pourshafie, M.-R.; Najafi, M.; Lari, E.R.; Lari, A.R.; Najafi, M. Tracing of false negative results in phenotypic methods for identification of carbapenemase by Real-time PCR. Gene 2016, 576, 166–170. [Google Scholar] [CrossRef]
- EUCAST Subcommittee for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. 2017. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance, Version 2.0. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf (accessed on 25 June 2020).
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- El Fertas-Aissani, R.; Messai, Y.; Alouache, S.; Bakour, R. Virulence profiles and antibiotic susceptibility patterns of Klebsiella pneumoniae strains isolated from different clinical specimens. Pathol. Biol. 2013, 61, 209–216. [Google Scholar] [CrossRef]
- Köser, C.U.; Ellington, M.J.; Cartwright, E.J.P.; Gillespie, S.H.; Brown, N.M.; Farrington, M.; Holden, M.T.G.; Dougan, G.; Bentley, S.D.; Parkhill, J.; et al. Routine Use of Microbial Whole Genome Sequencing in Diagnostic and Public Health Microbiology. PLoS Pathog. 2012, 8, e1002824. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive Web: User-Friendly Capsule and Lipopolysaccharide Serotype Prediction for Klebsiella Genomes. J. Clin. Microbiol. 2018, 56, 00197-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Pinto, N.A.; Kim, C.Y.; Yang, S.; D’Souza, R.; Yong, D.; Lee, I. Network Integrative Genomic and Transcriptomic Analysis of Carbapenem-Resistant Klebsiella pneumoniae Strains Identifies Genes for Antibiotic Resistance and Virulence. mSystems 2019, 4, e00202-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zowawi, H.M.; Forde, B.M.; Alfaresi, M.; Alzarouni, A.; Farahat, Y.; Chong, T.-M.; Yin, W.F.; Chan, K.-G.; Li, J.; Schembri, M.A.; et al. Stepwise evolution of pandrug-resistance in Klebsiella pneumoniae. Sci. Rep. 2015, 5, 15082. [Google Scholar] [CrossRef]
- Daikos, G.L.; Tsaousi, S.; Tzouvelekis, L.S.; Anyfantis, I.; Psichogiou, M.; Argyropoulou, A.; Stefanou, I.; Sypsa, V.; Miriagou, V.; Nepka, M.; et al. Carbapenemase-Producing Klebsiella pneumoniae Bloodstream Infections: Lowering Mortality by Antibiotic Combination Schemes and the Role of Carbapenems. Antimicrob. Agents Chemother. 2014, 58, 2322–2328. [Google Scholar] [CrossRef] [Green Version]
- Kęsik-Szeloch, A.; Drulis-Kawa, Z.; Weber-Dąbrowska, B.; Kassner, J.; Majkowska-Skrobek, G.; Augustyniak, D.; Łusiak-Szelachowska, M.; Żaczek, M.; Górski, A.; Kropinski, A.M. Characterising the biology of novel lytic bacteriophages infecting multidrug resistant Klebsiella pneumoniae. Virol. J. 2013, 10, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, T.A.; El-Sayed, L.H.; Haroun, M.; Hussein, A.A.; El Ashry, E.S.H. Development of immunization trials against Klebsiella pneumoniae. Vaccine 2012, 30, 2411–2420. [Google Scholar] [CrossRef] [PubMed]
- Somboro, A.M.; Sekyere, J.O.; Amoako, D.G.; Kumalo, H.M.; Khan, R.; Bester, L.A.; Essack, S. In vitro potentiation of carbapenems with tannic acid against carbapenemase-producing enterobacteriaceae: Exploring natural products as potential carbapenemase inhibitors. J. Appl. Microbiol. 2018, 126, 452–467. [Google Scholar] [CrossRef] [Green Version]
- Cassir, N.; Rolain, J.-M.; Brouqui, P. A new strategy to fight antimicrobial resistance: The revival of old antibiotics. Front. Microbiol. 2014, 5, 551. [Google Scholar] [CrossRef] [Green Version]
- Younis, W.; Abdelkhalek, A.; Mayhoub, A.S.; Seleem, M.N. In Vitro Screening of an FDA-Approved Library Against ESKAPE Pathogens. Curr. Pharm. Des. 2017, 23, 2147–2157. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, V.D.; Bat-Erdene, I.; Gupta, D.; Belkebir, S.; Rajhans, P.; Zand, F.; Myatra, S.N.; Afeef, M.; Tanzi, V.L.; Muralidharan, S.; et al. Six-year multicenter study on short-term peripheral venous catheters-related bloodstream infection rates in 727 intensive care units of 268 hospitals in 141 cities of 42 countries of Africa, the Americas, Eastern Mediterranean, Europe, South East Asia, and Western Pacific Regions: International Nosocomial Infection Control Consortium (INICC) findings. Infect. Control Hosp. Epidemiol. 2020, 41, 553–563. [Google Scholar] [CrossRef]
- Al-Hassan, L.; Roemer-Mahler, A.; Price, J.; Islam, J.; El-Mahallawy, H.; Higgins, P.G.; Hussein, A.F.A.; Roca, I.; Newport, M. The TACTIC experience: Establishing an international, interdisciplinary network to tackle antimicrobial resistance. J. Med. Microbiol. 2020, 69, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Document M100-S26; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Lutgring, J.D.; Limbago, B. The Problem of Carbapenemase-Producing-Carbapenem-Resistant-Enterobacteriaceae Detection. J. Clin. Microbiol. 2016, 54, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaneechoutte, M.; Dijkshoorn, L.; Tjernberg, I.; Elaichouni, A.; De Vos, P.; Claeys, G.; Verschraegen, G. Identification of Acinetobacter genomic species by amplified ribosomal DNA restriction analysis. J. Clin. Microbiol. 1995, 33, 11–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akpaka, P.E.; Swanston, W.H.; Ihemere, H.N.; Correa, A.; Torres, J.A.; Tafur, J.D.; Montealegre, M.C.; Quinn, J.P.; Villegas, M.V. Emergence of KPC-Producing Pseudomonas aeruginosa in Trinidad and Tobago. J. Clin. Microbiol. 2009, 47, 2670–2671. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Aktaş, Z.; Kayacan, C.B.; Schneider, I.; Can, B.; Midilli, K.; Bauernfeind, A. Carbapenem-Hydrolyzing Oxacillinase, OXA-48, Persists in Klebsiella pneumoniae in Istanbul, Turkey. Chemotherapy 2008, 54, 101–106. [Google Scholar] [CrossRef]
- Potron, A.; Nordmann, P.; Lafeuille, E.; Al Maskari, Z.; Al Rashdi, F.; Poirel, L. Characterization of OXA-181, a Carbapenem-Hydrolyzing Class D β-Lactamase from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 4896–4899. [Google Scholar] [CrossRef] [Green Version]
- Wilson, L.A. Sharp, P.M. Enterobacterial Repetitive Intergenic Consensus (ERIC) Sequences in Escherichia coli: Evolution and Implications for ERIC-PCR. Mol. Biol. Evol. 2006, 23, 1156–1168. [Google Scholar] [CrossRef] [Green Version]
- Mayer, L.W. Use of plasmid profiles in epidemiologic surveillance of disease outbreaks and in tracing the transmission of antibiotic resistance. Clin. Microbiol. Rev. 1988, 1, 228–243. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Lupski, J.R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Abdulall, A.K.; Tawfick, M.M.; El-Manakhly, A.R.; El-Kholy, A. Carbapenem-resistant Gram-negative bacteria associated with catheter-related bloodstream infections in three intensive care units in Egypt. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.N.; Chang, A.C.Y.; Hsu, L. Nonchromosomal Antibiotic Resistance in Bacteria: Genetic Transformation of Escherichia coli by R-Factor DNA. Proc. Natl. Acad. Sci. USA 1972, 69, 2110–2114. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Hennequin, C.; Forestier, C. Influence of capsule and extended-spectrum beta-lactamases encoding plasmids upon Klebsiella pneumoniae adhesion. Res. Microbiol. 2007, 158, 339–347. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | AUG | TZP | FOX | CTX | CTC | CAZ | CZC | CPM | ATM | MEM | ETR | GN | AK | CIP | STX | TGC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DH5α | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| transformant 383 | R | R | R | R | R | R | R | R | R | I | R | S | S | S | R | R |
| transformant 409 | R | R | R | R | R | R | R | R | R | R | R | S | S | S | R | R |
| Non-Susceptibility Pattern | Biofilm Pattern | p-Value (>0.05) | |||
|---|---|---|---|---|---|
| Strong | Moderate | Weak | |||
| A | Non-susceptible to all antimicrobials. | 1 | 21 | 52 | 0.3333 |
| B | Non-susceptible to all Except AK. | 0 | 0 | 2 | 0.6667 |
| C | Non-susceptible to all Except GN | 0 | 0 | 4 | 0.6667 |
| D | Non-susceptible to all Except CIP | 0 | 0 | 1 | 0.6667 |
| E | Non-susceptible to all Except STX | 0 | 3 | 2 | >0.9999 |
| F | Non-susceptible to all Except TGC | 0 | 7 | 13 | 0.3333 |
| G | Non-susceptible to all Except ATM & AK | 0 | 1 | 0 | >0.9999 |
| H | Non-susceptible to all Except GN & AK | 0 | 0 | 2 | 0.6667 |
| I | Non-susceptible to all Except TGC & AK | 0 | 1 | 0 | >0.9999 |
| J | Non-susceptible to all Except STX & TGC | 0 | 1 | 0 | >0.9999 |
| K | Non-susceptible to all Except GN & TGC | 0 | 0 | 3 | 0.6667 |
| L | Non-susceptible to all Except GN, CIP & STX | 0 | 0 | 1 | 0.6667 |
| M | Non-susceptible to all Except GN, STX & TGC | 0 | 0 | 1 | 0.6667 |
| N | Non-susceptible to all Except GN, CIP, STX & TGC | 0 | 0 | 1 | 0.6667 |
| Disc | Content | Interpretative Criteria |
|---|---|---|
| Meropenem (10 µg). | Confirmation of Carbapenem-non-susceptibility. | |
| B | Meropenem + MβL inhibitor. | Confirmation of MβL production (if only B-A ≥ 5 mm). |
| C | Meropenem + KPC inhibitor. | Confirmation of KPC production (if only C-A ≥ 4 mm). |
| D | Meropenem + AmpC inhibitor. | Confirmation of AmpC+ porin loss (if both C-A ≥ 4 mm and D-A ≥ 5 mm) |
| TEM | Temocillin (30 µg). | Indicative for OXA-48 (if there was no synergy detected and the inhibition zone of Temocillin was <11 mm) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragheb, S.M.; Tawfick, M.M.; El-Kholy, A.A.; Abdulall, A.K. Phenotypic and Genotypic Features of Klebsiella pneumoniae Harboring Carbapenemases in Egypt: OXA-48-Like Carbapenemases as an Investigated Model. Antibiotics 2020, 9, 852. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120852
Ragheb SM, Tawfick MM, El-Kholy AA, Abdulall AK. Phenotypic and Genotypic Features of Klebsiella pneumoniae Harboring Carbapenemases in Egypt: OXA-48-Like Carbapenemases as an Investigated Model. Antibiotics. 2020; 9(12):852. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120852
Chicago/Turabian StyleRagheb, Suzan Mohammed, Mahmoud Mohamed Tawfick, Amani Ali El-Kholy, and Abeer Khairy Abdulall. 2020. "Phenotypic and Genotypic Features of Klebsiella pneumoniae Harboring Carbapenemases in Egypt: OXA-48-Like Carbapenemases as an Investigated Model" Antibiotics 9, no. 12: 852. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120852