Pigs Overexpressing Porcine β-Defensin 2 Display Increased Resilience to Glaesserella parasuis Infection

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
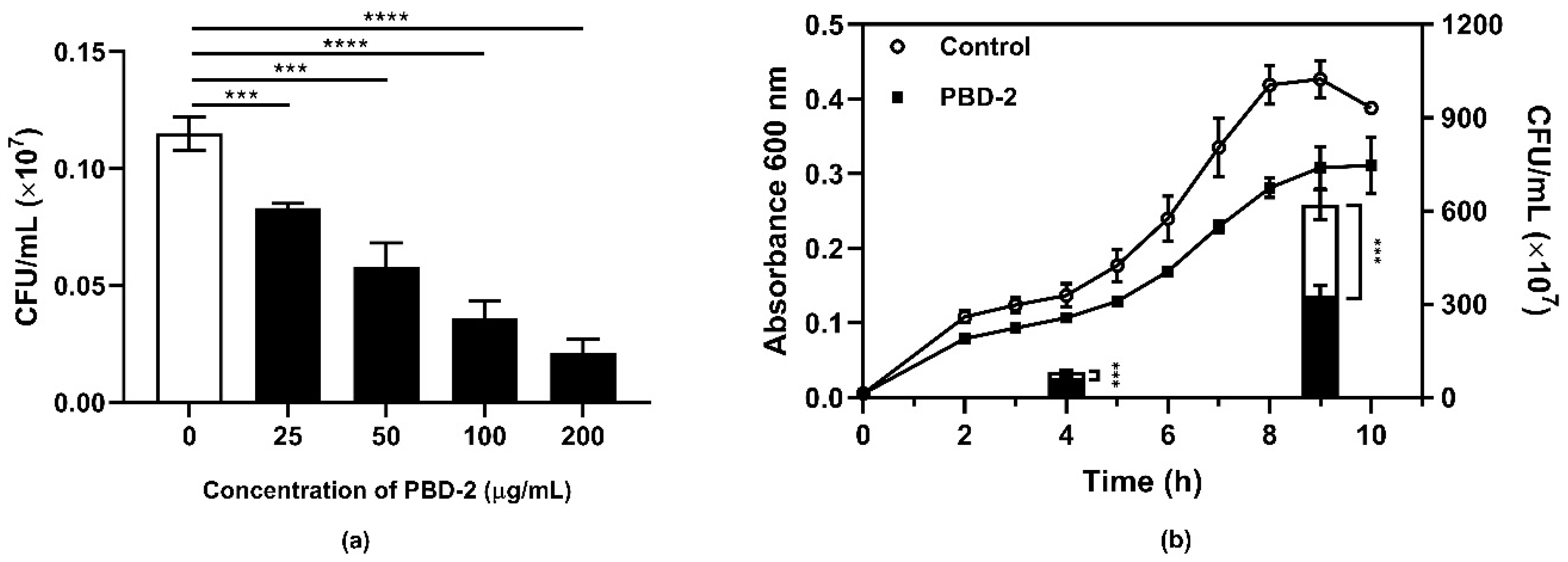
2.1. Antibacterial Activity of Synthetic PBD-2
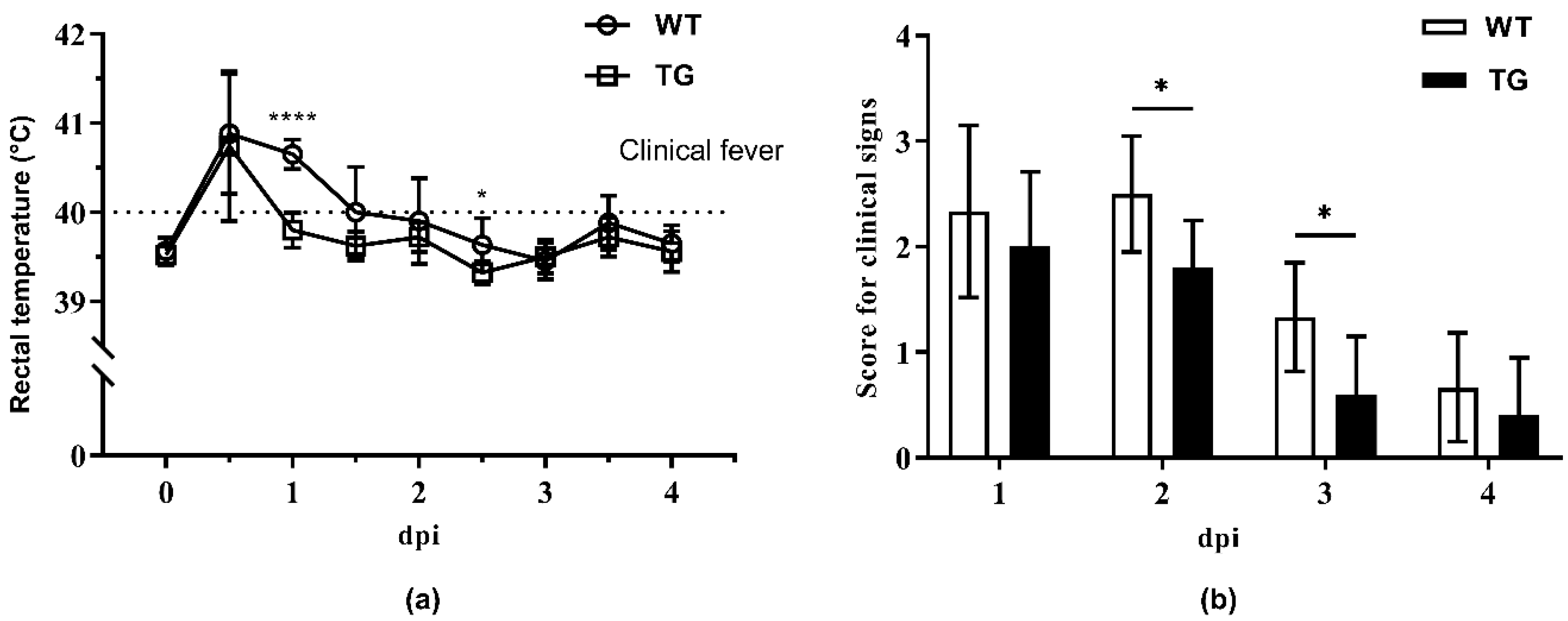
2.2. Clinical Monitoring during Cohabitation
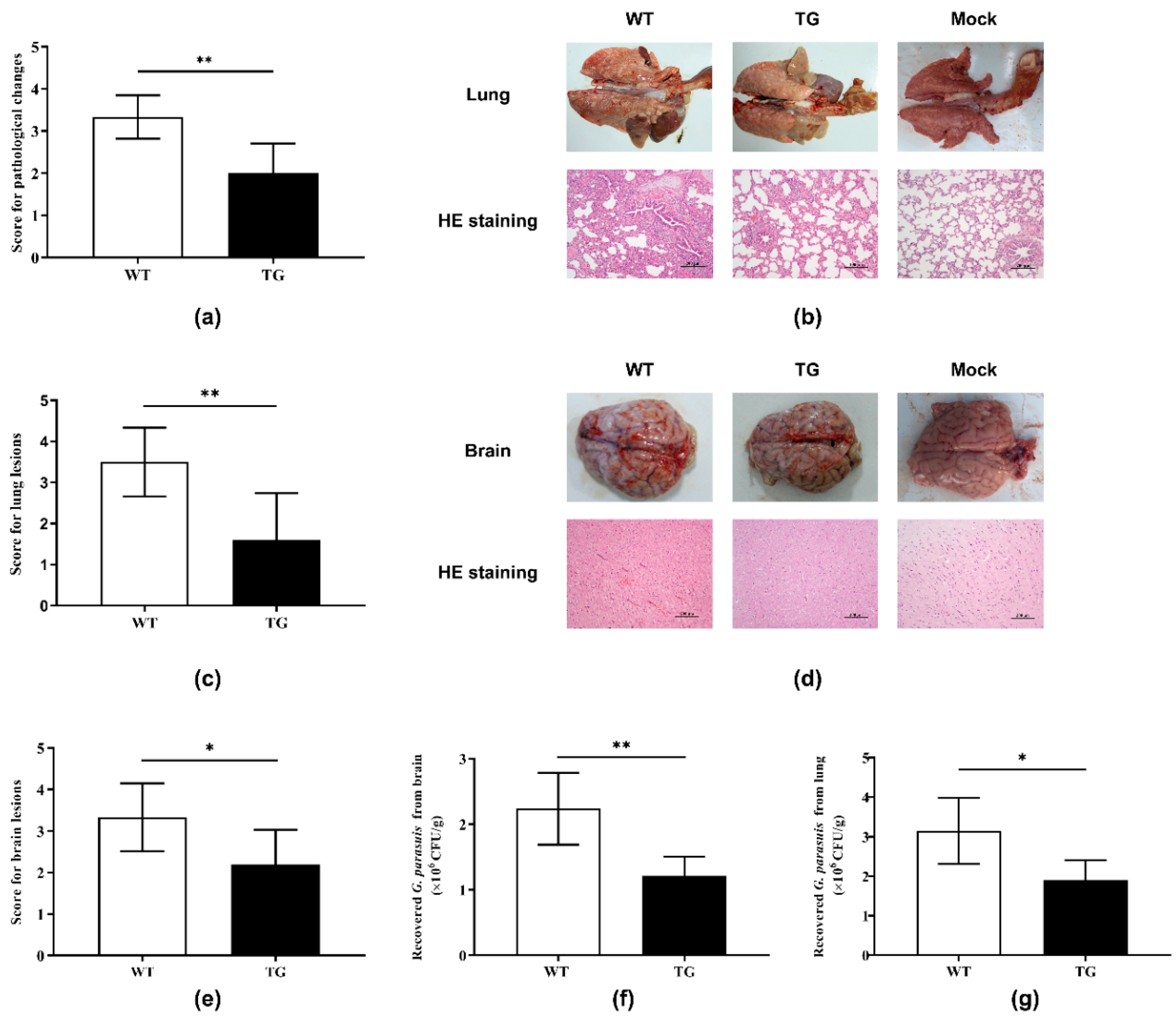
2.3. Postmortem Analysis
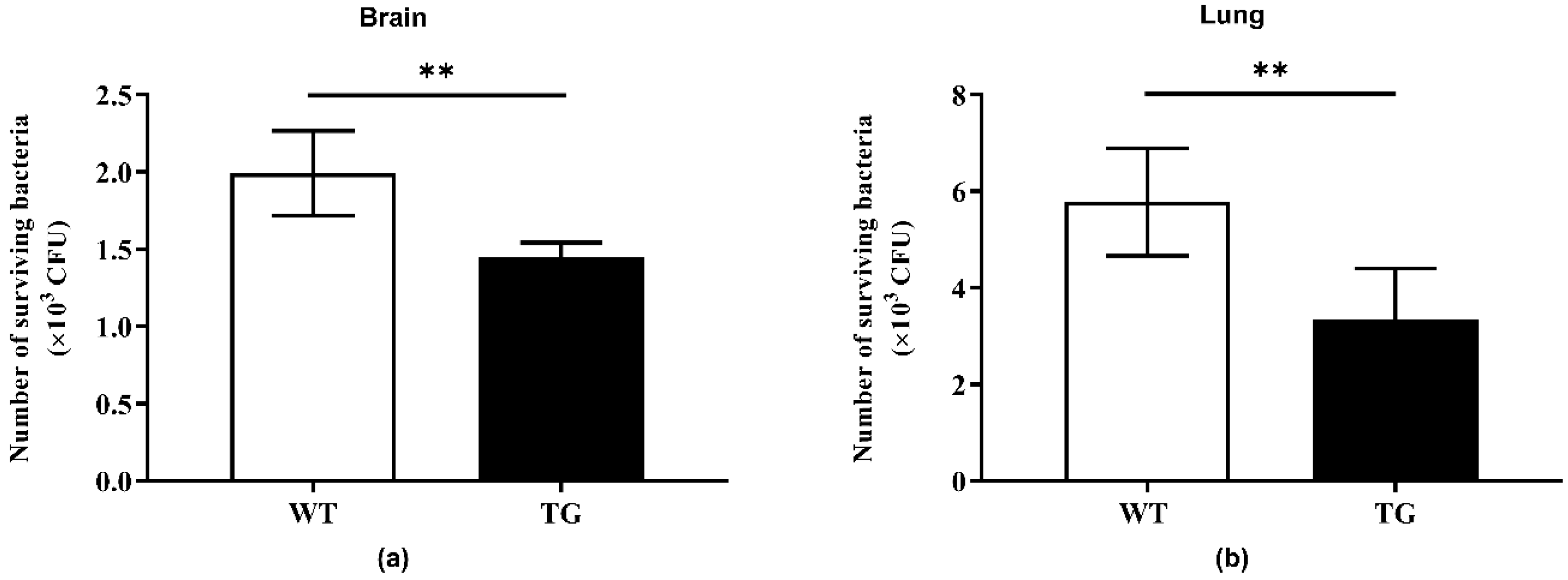
2.4. Bactericidal Activity of Tissue Homogenates
3. Discussion
4. Materials and Methods
4.1. Peptides, Bacterial Strains, and Animals
4.2. Determination of Antibacterial Activity of PBD-2
4.3. Cohabitation Challenge Trial
4.4. Histopathological Analysis
4.5. Quantification of Bacterial Loads in Pig Tissues
4.6. Detection of Bactericidal Effect of Tissue Homogenates from Pigs
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.; Chen, Z.; Cao, Z.; Li, W.; Wu, Y. Defensins, a novel type of animal toxin-like potassium channel inhibitor. Toxicon 2019, 157, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Ganz, T.; Lehrer, R.I. Defensins and other endogenous peptide antibiotics of vertebrates. J. Leukoc. Biol. 1995, 58, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Brown, K.L.; Mookherjee, N. Host defence peptides from invertebrates--emerging antimicrobial strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.; Hedegaard, C.J.; Haagsman, H.P.; Heegaard, P.M.H. The potential for immunoglobulins and host defense peptides (HDPs) to reduce the use of antibiotics in animal production. Vet. Res. 2018, 49, 68. [Google Scholar] [CrossRef] [Green Version]
- Mattar, E.H.; Almehdar, H.A.; Yacoub, H.A.; Uversky, V.N.; Redwan, E.M. Antimicrobial potentials and structural disorder of human and animal defensins. Cytokine Growth Factor Rev. 2016, 28, 95–111. [Google Scholar] [CrossRef]
- Veldhuizen, E.J.; Rijnders, M.; Claassen, E.A.; van Dijk, A.; Haagsman, H.P. Porcine beta-defensin 2 displays broad antimicrobial activity against pathogenic intestinal bacteria. Mol. Immunol. 2008, 45, 386–394. [Google Scholar] [CrossRef]
- Contreras, G.; Shirdel, I.; Braun, M.S.; Wink, M. Defensins: Transcriptional regulation and function beyond antimicrobial activity. Dev. Comp. Immunol. 2020, 104, 103556. [Google Scholar] [CrossRef]
- Fei, Z.; Hu, S.; Xiao, L.; Zhou, J.; Diao, H.; Yu, H.; Fang, S.; Wang, Y.; Wan, Y.; Wang, W.; et al. mBin1b transgenic mice show enhanced resistance to epididymal infection by bacteria challenge. Genes Immun. 2012, 13, 445–451. [Google Scholar] [CrossRef]
- Salzman, N.H.; Ghosh, D.; Huttner, K.M.; Paterson, Y.; Bevins, C.L. Protection against enteric salmonellosis in transgenic mice expressing a human intestinal defensin. Nature 2003, 422, 522–526. [Google Scholar] [CrossRef]
- Su, F.; Wang, Y.; Liu, G.; Ru, K.; Liu, X.; Yu, Y.; Liu, J.; Wu, Y.; Quan, F.; Guo, Z.; et al. Generation of transgenic cattle expressing human beta-defensin 3 as an approach to reducing susceptibility to Mycobacterium bovis infection. FEBS J. 2016, 283, 776–790. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Blecha, F. Porcine host defense peptides: Expanding repertoire and functions. Dev. Comp. Immunol. 2009, 33, 334–343. [Google Scholar] [CrossRef]
- Choi, M.K.; Le, M.T.; Nguyen, D.T.; Choi, H.; Kim, W.; Kim, J.H.; Chun, J.; Hyeon, J.; Seo, K.; Park, C. Genome-level identification, gene expression, and comparative analysis of porcine ss-defensin genes. BMC Genet. 2012, 13, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Wang, A.; Feng, Q.; Wang, Z.; Ivanova, I.V.; He, X.; Zhang, B.; Song, W. High-level expression, purification and characterisation of porcine beta-defensin 2 in Pichia pastoris and its potential as a cost-efficient growth promoter in porcine feed. Appl. Microbiol. Biotechnol. 2014, 98, 5487–5497. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, A.; Xie, L.; Song, W.; Wang, J.; Yin, Z.; Zhou, D.; Li, F. Use of recombinant porcine beta-defensin 2 as a medicated feed additive for weaned piglets. Sci. Rep. 2016, 6, 26790. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yao, S.; Chen, Y.; Gao, S.; Yang, Y.; Deng, J.; Ren, Z.; Shen, L.; Cui, H.; Hu, Y.; et al. Use of antimicrobial peptides as a feed additive for juvenile goats. Sci. Rep. 2017, 7, 12254. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Yang, X.; Huang, J.; Liu, X.; Yang, X.; Jin, H.; Huang, Q.; Li, L.; Zhou, R. Porcine Beta-Defensin 2 Provides Protection Against Bacterial Infection by a Direct Bactericidal Activity and Alleviates Inflammation via Interference With the TLR4/NF-kappaB Pathway. Front. Immunol. 2019, 10, 1673. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Qi, Y.; Wang, A.; Huang, C.; Liu, X.; Yang, X.; Li, L.; Zhou, R. Porcine beta-defensin 2 inhibits proliferation of pseudorabies virus in vitro and in transgenic mice. Virol. J. 2020, 17, 18. [Google Scholar] [CrossRef]
- Yang, X.; Cheng, Y.T.; Tan, M.F.; Zhang, H.W.; Liu, W.Q.; Zou, G.; Zhang, L.S.; Zhang, C.Y.; Deng, S.M.; Yu, L.; et al. Overexpression of Porcine Beta-Defensin 2 Enhances Resistance to Actinobacillus pleuropneumoniae Infection in Pigs. Infect. Immun. 2015, 83, 2836–2843. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, A.; Huang, C.; Sun, Y.; Song, B.; Zhou, R.; Li, L. Generation of Marker-Free pbd-2 Knock-in Pigs Using the CRISPR/Cas9 and Cre/loxP Systems. Genes 2020, 11, 951. [Google Scholar] [CrossRef]
- Aragon, V.; Segalés, J.; Tucker, A.W.D. Glässer’s Disease. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons: Ames, IA, USA, 2019; pp. 844–853. [Google Scholar] [CrossRef]
- Morozumi, T.; Hiramune, T.; Kobayashi, K. Glässer‘s disease in piglets produced by intraperitoneal inoculation with Haemophilus parasuis. Natl. Inst. Anim. Health Q. 1981, 21, 121–128. [Google Scholar]
- Del‘Arco, A.E.; Santos, J.L.; Bevilacqua, P.D.; Faria, J.E.; Guimarães, W.V. Swine infection by Streptococcus suis: A retrospective study. Arq. Bras. Med. Vet. Zootec. 2008, 60, 878–883. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Li, C.; Wang, C.; Liu, Y.; Wang, G.; He, X.; Hu, L.; Liu, Y.; Cui, M.; et al. Secondary Haemophilus parasuis infection enhances highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection-mediated inflammatory responses. Vet. Microbiol. 2017, 204, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, W.; Wang, Y.; Gu, C.; Liu, X.; Charreyre, C.; Fan, S.; He, Q. Coinfection with Haemophilus parasuis serovar 4 increases the virulence of porcine circovirus type 2 in piglets. Virol. J. 2017, 14, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Wang, C.; Xue, Y.; Tang, X.; Wu, B.; Cheng, X.; He, Q.; Chen, H. The occurrence of Bordetella bronchiseptica in pigs with clinical respiratory disease. Vet. J. 2011, 188, 337–340. [Google Scholar] [CrossRef]
- Kielstein, P.; Rapp-Gabrielson, V.J. Designation of 15 serovars of Haemophilus parasuis on the basis of immunodiffusion using heat-stable antigen extracts. J. Clin. Microbiol. 1992, 30, 862–865. [Google Scholar] [CrossRef] [Green Version]
- Angen, O.; Svensmark, B.; Mittal, K.R. Serological characterization of Danish Haemophilus parasuis isolates. Vet. Microbiol. 2004, 103, 255–258. [Google Scholar] [CrossRef]
- Rubies, X.; Kielstein, P.; Costa, L.; Riera, P.; Artigas, C.; Espuna, E. Prevalence of Haemophilus parasuis serovars isolated in Spain from 1993 to 1997. Vet. Microbiol. 1999, 66, 245–248. [Google Scholar] [CrossRef]
- Castilla, K.S.; de Gobbi, D.D.; Moreno, L.Z.; Paixao, R.; Coutinho, T.A.; dos Santos, J.L.; Moreno, A.M. Characterization of Haemophilus parasuis isolated from Brazilian swine through serotyping, AFLP and PFGE. Res. Vet. Sci. 2012, 92, 366–371. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, C.; Guo, L.; Shen, H.; Deng, X.; Ke, C.; Ke, B.; Zhang, B.; Li, A.; Ren, T.; et al. Prevalence and characterization of genotypic diversity of Haemophilus parasuis isolates from southern China. Can. J. Vet. Res. 2012, 76, 224–229. [Google Scholar] [PubMed]
- Zhao, Y.; Wang, Q.; Li, J.; Lin, X.; Huang, X.; Fang, B. Epidemiology of Haemophilus parasuis isolates from pigs in China using serotyping, antimicrobial susceptibility, biofilm formation and ERIC-PCR genotyping. PeerJ 2018, 6, e5040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, C.N.; Thanh, T.V.T.; Zou, G.; Jia, M.; Wang, Q.; Zhang, L.; Ding, W.; Huang, Q.; Zhou, R. Characterization of serotypes and virulence genes of Haemophilus parasuis isolates from Central Vietnam. Vet. Microbiol. 2019, 230, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, H.; Xue, Y.; Chen, K.; Liu, Z.; Xue, Q.; Wang, C. Analysis of efficacy obtained with a trivalent inactivated Haemophilus parasuis serovars 4, 5, and 12 vaccine and commercial vaccines against Glässer‘s disease in piglets. Can. J. Vet. Res. 2017, 81, 22–27. [Google Scholar] [PubMed]
- Nielsen, R. Pathogenicity and immunity studies of Haemophilus parasuis serotypes. Acta Vet. Scand. 1993, 34, 193–198. [Google Scholar] [PubMed]
- de la Fuente, A.J.; Tucker, A.W.; Navas, J.; Blanco, M.; Morris, S.J.; Gutierrez-Martin, C.B. Antimicrobial susceptibility patterns of Haemophilus parasuis from pigs in the United Kingdom and Spain. Vet. Microbiol. 2007, 120, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xu, C.; Shen, H.; Li, J.; Guo, L.; Cao, G.; Feng, S.; Liao, M. Biofilm formation in Haemophilus parasuis: Relationship with antibiotic resistance, serotype and genetic typing. Res. Vet. Sci. 2014, 97, 171–175. [Google Scholar] [CrossRef]
- Brogden, S.; Pavlovic, A.; Tegeler, R.; Kaspar, H.; De Vaan, N.; Kehrenberg, C. Antimicrobial susceptibility of Haemophilus parasuis isolates from Germany by use of a proposed standard method for harmonized testing. Vet. Microbiol. 2018, 217, 32–35. [Google Scholar] [CrossRef]
- Xie, K.; Xie, H.; Su, G.; Chen, D.; Yu, B.; Mao, X.; Huang, Z.; Yu, J.; Luo, J.; Zheng, P.; et al. Beta-Defensin 129 Attenuates Bacterial Endotoxin-Induced Inflammation and Intestinal Epithelial Cell Apoptosis. Front. Immunol. 2019, 10, 2333. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, G.; Wu, H.; Ross, C.; Blecha, F.; Ganz, T. Porcine epithelial beta-defensin 1 is expressed in the dorsal tongue at antimicrobial concentrations. Infect. Immun. 1999, 67, 3121–3127. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Patil, A.A.; Zhang, G.; Ross, C.R.; Blecha, F. Bioinformatic and expression analysis of novel porcine beta-defensins. Mamm. Genome 2006, 17, 332–339. [Google Scholar] [CrossRef]
- Xu, J.; Zhong, F.; Zhang, Y.; Zhang, J.; Huo, S.; Lin, H.; Wang, L.; Cui, D.; Li, X. Construction of Bacillus subtilis strain engineered for expression of porcine beta-defensin-2/cecropin P1 fusion antimicrobial peptides and its growth-promoting effect and antimicrobial activity. Asian-Australas J. Anim. Sci. 2017, 30, 576–584. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Fu, S.; Hu, J.; Li, J.; Chang, H.; Hu, L.; Chen, H.; Tian, Y.; Bei, W. Evaluation of recombinant proteins of Haemophilus parasuis strain SH0165 as vaccine candidates in a mouse model. Res. Vet. Sci. 2012, 93, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xu, X.; Zhao, Y.; Chen, P.; Zhang, X.; Chen, H.; Cai, X. Distribution of antimicrobial resistance among different serovars of Haemophilus parasuis isolates. Vet. Microbiol. 2010, 141, 168–173. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Yang, P.; Wang, L.; Jiang, X.; Zhang, W.; Liang, X.; Yin, L.; Yin, Z.; Geng, Y.; Zhong, Z.; et al. Antibacterial effect of Blumea balsamifera DC. essential oil against Haemophilus parasuis. Arch. Microbiol. 2020, 202, 2499–2508. [Google Scholar] [CrossRef]
- Li, L.; Song, X.; Yin, Z.; Jia, R.; Li, Z.; Zhou, X.; Zou, Y.; Li, L.; Yin, L.; Yue, G.; et al. The antibacterial activity and action mechanism of emodin from Polygonum cuspidatum against Haemophilus parasuis in vitro. Microbiol. Res. 2016, 186–187, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tian, Y.; Yu, J.; Song, X.; Jia, R.; Cui, Q.; Tong, W.; Zou, Y.; Li, L.; Yin, L.; et al. iTRAQ-based quantitative proteomic analysis reveals multiple effects of Emodin to Haemophilus parasuis. J. Proteom. 2017, 166, 39–47. [Google Scholar] [CrossRef]
- Ye, C.; Li, R.; Xu, L.; Qiu, Y.; Fu, S.; Liu, Y.; Wu, Z.; Hou, Y.; Hu, C.A. Effects of Baicalin on piglet monocytes involving PKC-MAPK signaling pathways induced by Haemophilus parasuis. BMC Vet. Res. 2019, 15, 98. [Google Scholar] [CrossRef]
- Nakajima, Y.; Ishibashi, J.; Yukuhiro, F.; Asaoka, A.; Taylor, D.; Yamakawa, M. Antibacterial activity and mechanism of action of tick defensin against Gram-positive bacteria. Biochim. Biophys. Acta 2003, 1624, 125–130. [Google Scholar] [CrossRef]
- Yenugu, S.; Hamil, K.G.; Radhakrishnan, Y.; French, F.S.; Hall, S.H. The androgen-regulated epididymal sperm-binding protein, human beta-defensin 118 (DEFB118) (formerly ESC42), is an antimicrobial beta-defensin. Endocrinology 2004, 145, 3165–3173. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Han, Y.; Yang, D.; Zhao, J.; Wang, C.; Mu, C. Antibacterial activities and mechanisms of action of a defensin from manila clam Ruditapes philippinarum. Fish Shellfish Immunol. 2020, 103, 266–276. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, C.; Zhang, X.; Zhang, M.Z.; Rottinghaus, G.E.; Zhang, S. Structure-function analysis of Avian beta-defensin-6 and beta-defensin-12: Role of charge and disulfide bridges. BMC Microbiol. 2016, 16, 210. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, P.; Wilmes, M.; Pugniere, M.; Aumelas, A.; Bachere, E.; Sahl, H.G.; Schneider, T.; Destoumieux-Garzon, D. Insight into invertebrate defensin mechanism of action: Oyster defensins inhibit peptidoglycan biosynthesis by binding to lipid II. J. Biol. Chem. 2010, 285, 29208–29216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chairatana, P.; Nolan, E.M. Molecular basis for self-assembly of a human host-defense peptide that entraps bacterial pathogens. J. Am. Chem. Soc. 2014, 136, 13267–13276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Xie, Z.; Zhang, Q.; Li, Y.; Yang, F.; Chen, Z.; Li, W.; Cao, Z.; Wu, Y. Scorpion Potassium Channel-blocking Defensin Highlights a Functional Link with Neurotoxin. J. Biol. Chem. 2016, 291, 7097–7106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.B.; Zhang, K.; Zhang, H.; Gao, C.Y.; Li, C.L. Analysis of the antimicrobial mechanism of porcine beta defensin 2 against E. coli by electron microscopy and differentially expressed genes. Sci. Rep. 2018, 8, 14711. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Zhang, H.; Gao, C.; Chen, R.; Li, C. Antimicrobial Mechanism of pBD2 against Staphylococcus aureus. Molecules 2020, 25, 3513. [Google Scholar] [CrossRef]
- Lu, T.; Song, Z.; Li, Q.; Li, Z.; Wang, M.; Liu, L.; Tian, K.; Li, N. Overexpression of Histone Deacetylase 6 Enhances Resistance to Porcine Reproductive and Respiratory Syndrome Virus in Pigs. PLoS ONE 2017, 12, e0169317. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Bao, Y.; Li, J.; Chen, Z.; Yu, X.; Zhao, Y.; Tian, K.; Li, N. RNAi-based inhibition of porcine reproductive and respiratory syndrome virus replication in transgenic pigs. J. Biotechnol. 2014, 171, 17–24. [Google Scholar] [CrossRef]
- Hu, S.; Qiao, J.; Fu, Q.; Chen, C.; Ni, W.; Wujiafu, S.; Ma, S.; Zhang, H.; Sheng, J.; Wang, P.; et al. Transgenic shRNA pigs reduce susceptibility to foot and mouth disease virus infection. Elife 2015, 4, e06951. [Google Scholar] [CrossRef]
- Xie, Z.; Pang, D.; Yuan, H.; Jiao, H.; Lu, C.; Wang, K.; Yang, Q.; Li, M.; Chen, X.; Yu, T.; et al. Genetically modified pigs are protected from classical swine fever virus. PLoS Pathog. 2018, 14, e1007193. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Jiao, H.; Xiao, H.; Jiang, Y.; Liu, Z.; Qi, C.; Zhao, D.; Jiao, S.; Yu, T.; Tang, X.; et al. Generation of pRSAD2 gene knock-in pig via CRISPR/Cas9 technology. Antiviral. Res. 2020, 174, 104696. [Google Scholar] [CrossRef]
- Huang, G.; Liu, X.; Tang, X.; Du, L.; Feng, W.; Hu, X.; Zhu, L.; Li, Q.; Suo, X. Increased Neutralizing Antibody Production and Interferon-gamma Secretion in Response to Porcine Reproductive and Respiratory Syndrome Virus Immunization in Genetically Modified Pigs. Front. Immunol. 2017, 8, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Sun, Y.; Carretero, O.A.; Zhu, L.; Harding, P.; Shesely, E.G.; Dai, X.; Rhaleb, N.E.; Peterson, E.; Yang, X.P. Effects of cardiac overexpression of the angiotensin II type 2 receptor on remodeling and dysfunction in mice post-myocardial infarction. Hypertension 2014, 63, 1251–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, M.; Yang, F.; Yang, J.; Bei, W.; Cai, X.; Chen, L.; Dong, J.; Zhou, R.; Jin, M.; Jin, Q.; et al. Complete genome sequence of Haemophilus parasuis SH0165. J. Bacteriol. 2009, 191, 1359–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, H.; Riising, H.J. Protection of vaccinated pigs against experimental infections with homologous and heterologous Haemophilus parasuis. Vet. Rec. 2002, 151, 502–505. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Yang, X.; Wang, A.; Huang, C.; Tang, H.; Zhang, Q.; Fang, Q.; Yu, Z.; Liu, X.; Huang, Q.; et al. Pigs Overexpressing Porcine β-Defensin 2 Display Increased Resilience to Glaesserella parasuis Infection. Antibiotics 2020, 9, 903. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120903
Huang J, Yang X, Wang A, Huang C, Tang H, Zhang Q, Fang Q, Yu Z, Liu X, Huang Q, et al. Pigs Overexpressing Porcine β-Defensin 2 Display Increased Resilience to Glaesserella parasuis Infection. Antibiotics. 2020; 9(12):903. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120903
Chicago/Turabian StyleHuang, Jing, Xiaoyu Yang, Antian Wang, Chao Huang, Hao Tang, Qiuhong Zhang, Qiong Fang, Zuming Yu, Xiao Liu, Qi Huang, and et al. 2020. "Pigs Overexpressing Porcine β-Defensin 2 Display Increased Resilience to Glaesserella parasuis Infection" Antibiotics 9, no. 12: 903. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120903