Antibacterial and Antivirulence Activity of Glucocorticoid PYED-1 against Stenotrophomonas maltophilia

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
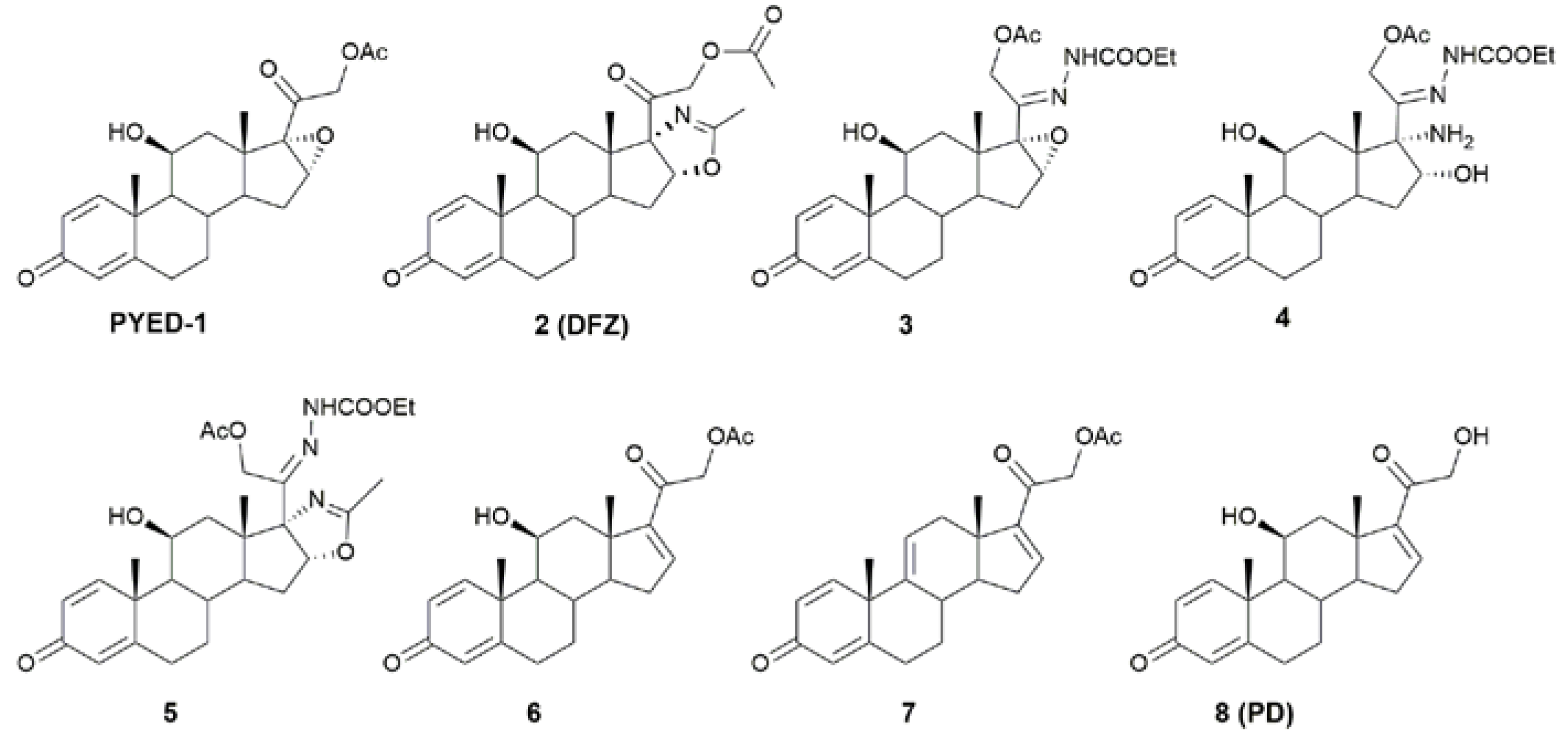
2.1. Antimicrobial Activity of a Panel of Steroid Derivatives
2.2. Checkerboard Assay
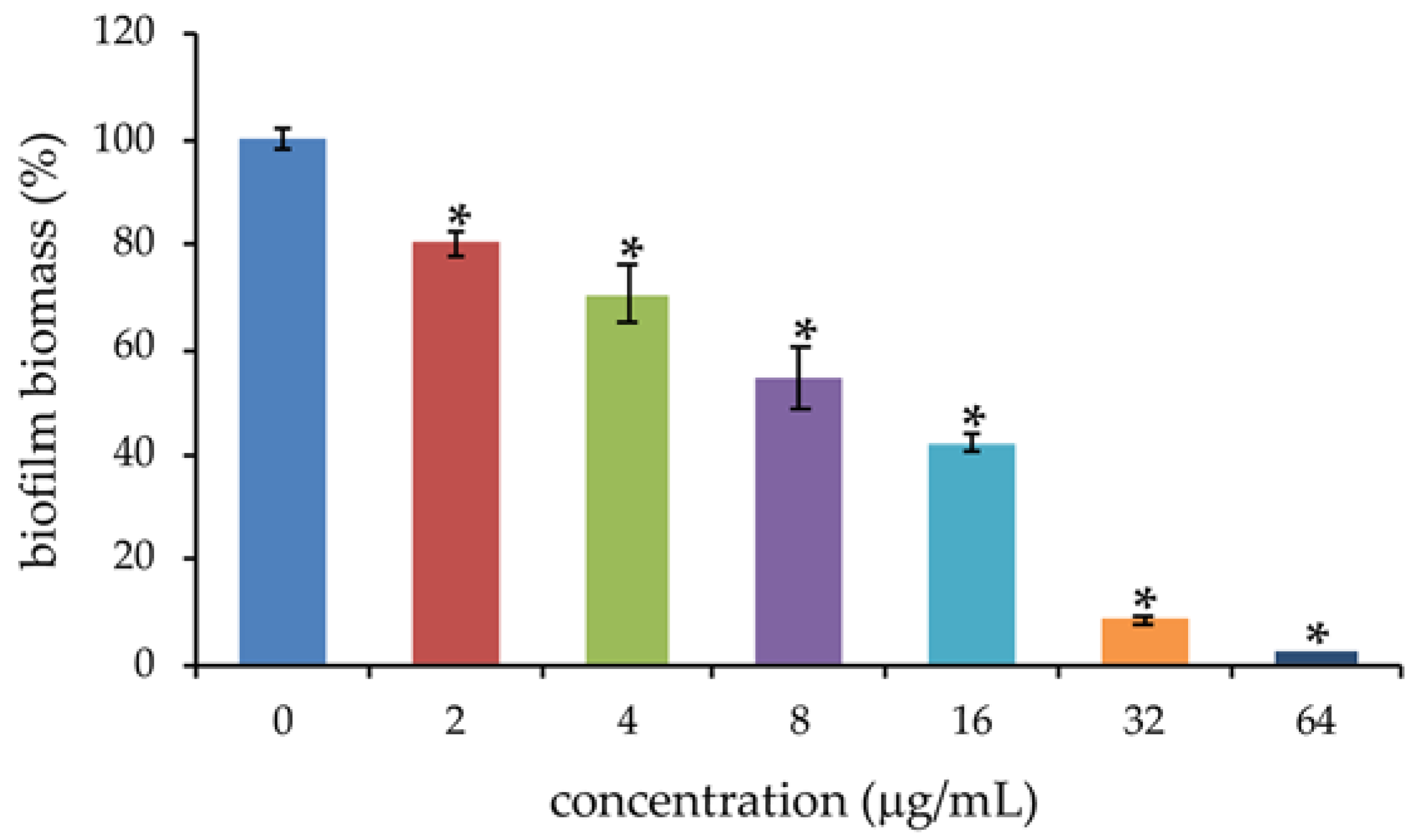
2.3. Effects of PYED-1 on the Formation of S. maltophilia Biofilm
2.4. Transcriptional Changes Induced by PYED-1 in S. maltophilia K279a
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Antimicrobial Activity
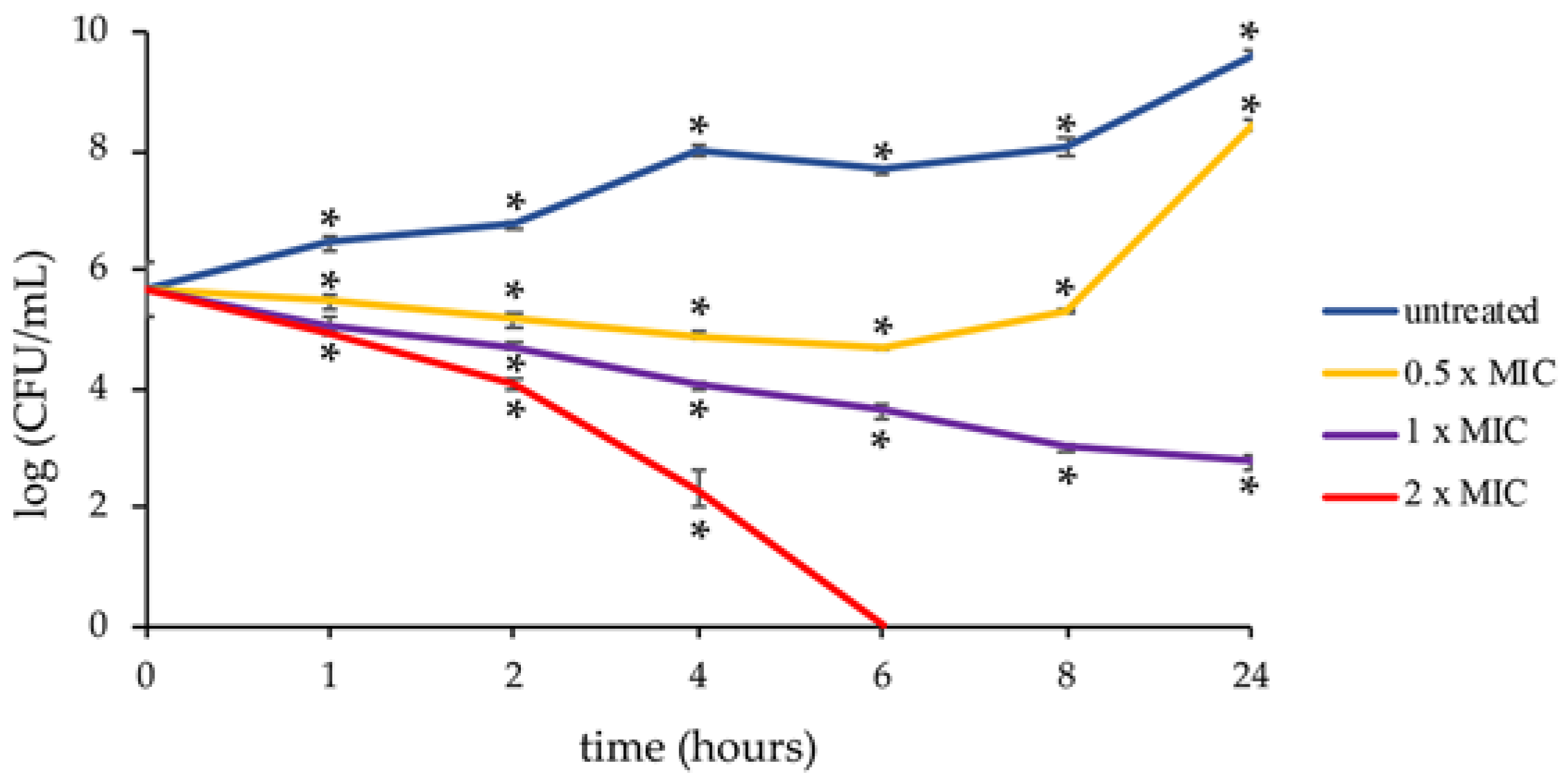
3.3. Time Killing Assay
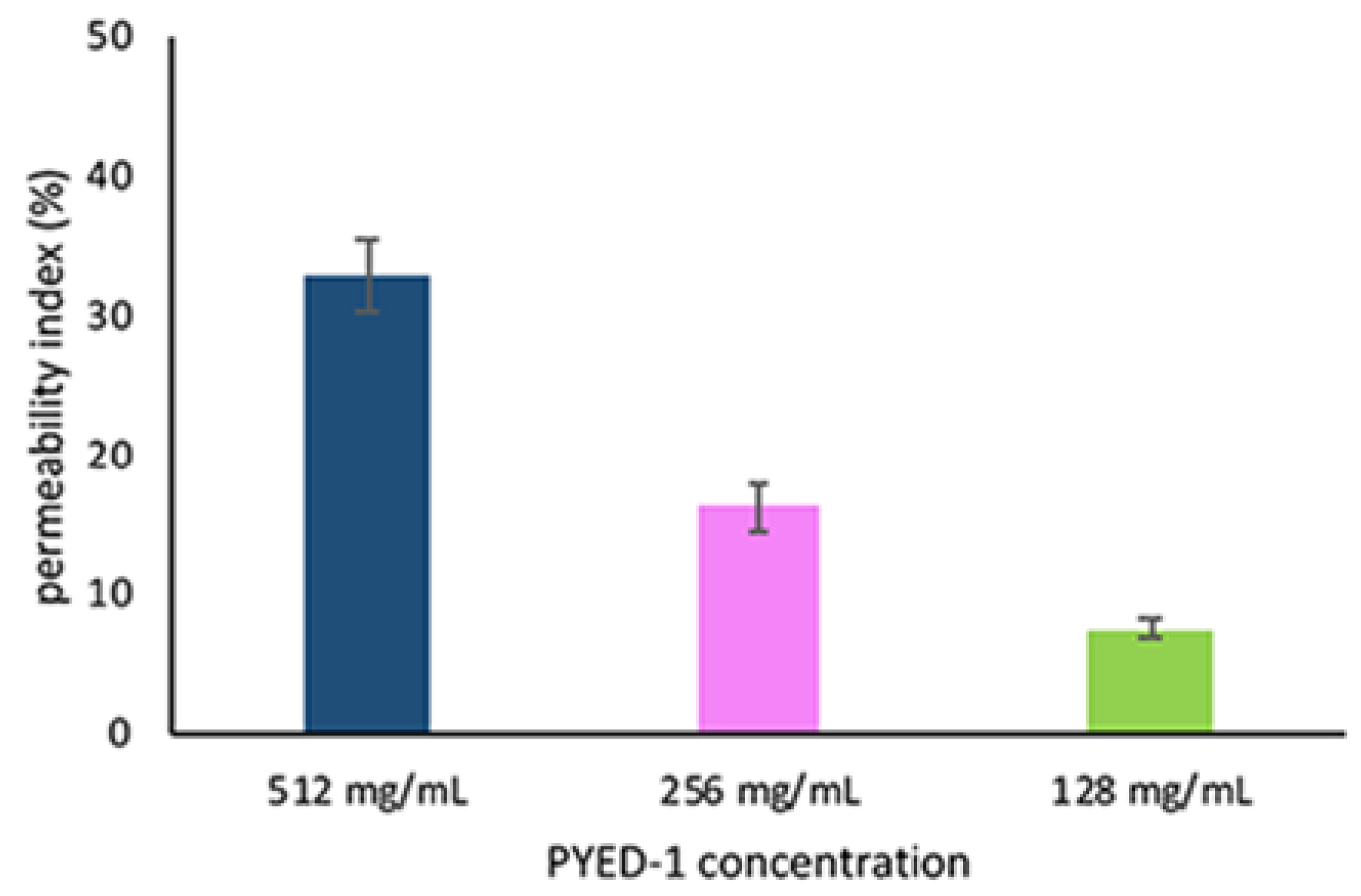
3.4. Propidium Iodide Uptake Assay
3.5. Checkerboard Assay
3.6. Biofilm Assay
3.7. RNA
3.8. RT-PCR
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fihman, V.; Le Monnier, A.; Corvec, S.; Jaureguy, F.; Tankovic, J.; Jacquier, H.; Carbonnelle, E.; Bille, E.; Illiaquer, M.; Cattoir, V.; et al. Stenotrophomonas maltophilia—the most worrisome threat among unusual non-fermentative gram-negative bacilli from hospitalized patients: A prospective multicenter study. J. Infect. 2012, 64, 391–398. [Google Scholar] [CrossRef]
- Looney, W.J.; Narita, M.; Mühlemann, K. Stenotrophomonas maltophilia: An emerging opportunist human pathogen. Lancet Infect. Dis. 2009, 9, 312–323. [Google Scholar] [CrossRef]
- Crispino, M.; Boccia, M.C.; Bagattini, M.; Villari, P.; Triassi, M.; Zarrilli, R. Molecular epidemiology of Stenotrophomonas maltophilia in a university hospital. J. Hosp. Infect. 2002, 52, 88–92. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Rev. Clin. Microbiol. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, Y.D.; Jeong, W.Y.; Kim, M.H.; Jung, I.Y.; Ahn, M.Y.; Ann, H.W.; Ahn, J.Y.; Han, S.H.; Choi, J.Y.; Song, Y.G.; et al. Risk factors for mortality in patients with Stenotrophomonas maltophilia bacteremia. Medicine 2016, 95, e4375. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.; Nazareth, D.; Shaw, M.; Walshaw, M.J. Cystic fibrosis related diabetes is not independently associated with increased Stenotrophomonas maltophilia infection: Longitudinal data from the UK CF Registry. J. Cyst. Fibros. 2019, 18, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Barsky, E.E.; Williams, K.A.; Priebe, G.P.; Sawicki, G.S. Incident Stenotrophomonas maltophilia infection and lung function decline in cystic fibrosis. Pediatr Pulmonol. 2017, 52, 1276–1282. [Google Scholar] [CrossRef]
- Waters, V.; Atenafu, E.G.; Lu, A.; Yau, Y.; Tullis, E.; Ratjen, F. Chronic Stenotrophomonas maltophilia infection and mortality or lung transplantation in cystic fibrosis patients. J. Cyst. Fibros. 2013, 12, 482–486. [Google Scholar] [CrossRef] [Green Version]
- Adegoke, A.A.; Stenström, T.A.; Okoh, A.I. Stenotrophomonas maltophilia as an Emerging Ubiquitous Pathogen: Looking Beyond Contemporary Antibiotic Therapy. Front. Microbiol. 2017, 8, 2276. [Google Scholar] [CrossRef] [Green Version]
- Bayer-Santos, E.; Cenens, W.; Matsuyama, B.Y.; Oka, G.U.; Di Sessa, G.; Mininel, I.; Alves, T.; Farah, C.S. The opportunistic pathogen Stenotrophomonas maltophilia utilizes a type IV secretion system for interbacterial killing. PLoS Pathog. 2019, 15, e1007651. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Prosseda, G.; Del Chierico, F.; Cannavacciuolo, S.; Cipriani, P.; Petrucca, A.; Superti, F.; Ammendolia, M.G.; Concato, C.; Fiscarelli, E.; et al. Molecular characterization of virulence determinants of Stenotrophomonas maltophilia strains isolated from patients affected by cystic fibrosis. Int. J. Immunopathol. Pharmacol. 2007, 20, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Nas, M.Y.; White, R.C.; DuMont, A.L.; Lopez, A.E.; Cianciotto, N.P. Stenotrophomonas maltophilia encodes a VirB/VirD4 Type IV Secretion System that modulates apoptosis in human cells and promotes competition against heterologous bacteria, including Pseudomonas aeruginosa. Infect. Immun. 2019, 87, e00457-19. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, M.; Iacobino, A.; Prosseda, G.; Fiscarelli, E.; Zarrilli, R.; De Carolis, E.; Petrucca, A.; Nencioni, L.; Colonna, B.; Casalino, M. Stenotrophomonas maltophilia strains from cystic fibrosis patients: Genomic variability and molecular characterization of some virulence determinants. Int. J. Med. Microbiol. 2011, 301, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Pomponio, S.; Crocetta, V.; Gherardi, G.; Verginelli, F.; Fiscarelli, E.; Dicuonzo, G.; Savini, V.; D’Antonio, D.; Di Bonaventura, G. Phenotypic and genotypic characterization of Stenotrophomonas maltophilia isolates from patients with cystic fibrosis: Genome diversity, biofilm formation, and virulence. BMC Microbiol. 2011, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifonova, A.; Strateva, T. Stenotrophomonas maltophilia—A low-grade pathogen with numerous virulence factors. Infect. Dis. 2019, 51, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.T.; Lin, C.Y.; Chen, Y.H.; Hsueh, P.R. Update on infections caused by Stenotrophomonas maltophilia with particular attention to resistance mechanisms and therapeutic options. Front. Microbiol 2015, 6, 893. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.B. Antibiotic resistance in the opportunistic pathogen Stenotrophomonas maltophilia. Front. Microbiol. 2015, 6, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, E.; Liang, G.; Wang, L.; Wei, W.; Lei, M.; Song, S.; Han, R.; Wang, Y.; Qi, W. Antimicrobial susceptibility of hospital acquired Stenotrophomonas maltophilia isolate biofilms. Braz. J. Infect. Dis. 2016, 20, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Toleman, M.A.; Bennett, P.M.; Bennett, D.M.; Jones, R.N.; Walsh, T.R. Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerg. Infect. Dis. 2007, 13, 559–565. [Google Scholar] [CrossRef]
- Alcaraz, E.; Garcıa, C.; Friedman, L.; Passerini de Rossi, B. The rpf/DSF signalling system of Stenotrophomonas maltophilia positively regulates biofilm formation, production of virulence-associated factors and β-lactamase induction. FEMS Microbiol. Lett. 2019, 366, fnz069. [Google Scholar] [CrossRef]
- Flores-Treviño, S.; Bocanegra-Ibarias, P.; Camacho-Ortiz, A.; Morfín-Otero, R.; Salazar-Sesatty, H.A.; Garza-González, E. Stenotrophomonas maltophilia biofilm: Its role in infectious diseases. Expert. Rev. Anti Infect. Ther. 2019, 17, 877–893. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Savini, V.; Fiscarelli, E.; Gherardi, G.; Di Bonaventura, G. Clonal diversity, biofilm formation, and antimicrobial resistance among Stenotrophomonas maltophilia strains from cystic fibrosis and non-cystic fibrosis patients. Antibiotics (Basel) 2020, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Perez, O.; Luquin, S.; Garcia-Estrada, J.; Ramos-Remus, C. Deflazacort: A glucocorticoid with few metabolic adverse effects but important immunosuppressive activity. Adv. Ther. 2007, 24, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Parente, L. Deflazacort: Therapeutic index, relative potency and equivalent doses versus other corticosteroids. BMC Pharmacol. Toxicol. 2017, 18, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, A.; De Gregorio, E.; De Fenza, M.; D’Alonzo, D.; Satawani, A.; Guaragna, A. Expeditious synthesis and preliminary antimicrobial activity of deflazacort and its precursors. RSC Advances. 2019, 9, 21519–21524. [Google Scholar] [CrossRef] [Green Version]
- Levison, M.E. Pharmacodynamics of antimicrobial drugs. Infect. Dis Clin. North. Am. Rev. 2004, 18, 451–465. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 29th Informational CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Karunanidhi, A.; Thomas, R.; van Belkum, A.; Neela, V. In vitro antibacterial and antibiofilm activities of chlorogenic acid against clinical isolates of Stenotrophomonas maltophilia including the trimethoprim/sulfamethoxazole resistant strain. Biomed. Res. Int. 2013, 2013, 392058. [Google Scholar] [CrossRef] [Green Version]
- Karunanidhi, A.; Ghaznavi-Rad, E.; Hamat, R.A.; Pichika, M.R.; Lung, L.T.T.; Mohd Fauzi, F.; Chigurupati, S.; van Belkum, A.; Neela, V. Antibacterial and Antibiofilm Activities of Nonpolar Extracts of Allium stipitatum Regel. against Multidrug Resistant Bacteria. Biomed. Res. Int. 2018, 2018, 9845075. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Lee, D.; Eom, Y.B. Anti-biofilm and Anti-Virulence Efficacy of Celastrol against Stenotrophomonas maltophilia. Int. J. Med. Sci. 2018, 15, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Pollini, S.; Di Pilato, V.; Landini, G.; Di Maggio, T.; Cannatelli, A.; Sottotetti, S.; Cariani, L.; Aliberti, S.; Blasi, F.; Sergio, F.; et al. In vitro activity of N-acetylcysteine against Stenotrophomonas maltophilia and Burkholderia cepacia complex grown in planktonic phase and biofilm. PLoS ONE. 2018, 13, e0203941. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; Scocchi, M.; Pomponio, S.; Di Vincenzo, V.; Mardirossian, M.; Gherardi, G.; Fiscarelli, E.; Dicuonzo, G.; Gennaro, R.; et al. Potential novel therapeutic strategies in cystic fibrosis: Antimicrobial and anti-biofilm activity of natural and designed α-helical peptides against Staphylococcus aureus, Pseudomonas aeruginosa, and Stenotrophomonas maltophilia. BMC Microbiol. 2012, 12, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidigal, P.G.; Müsken, M.; Becker, K.A.; Häussler, S.; Wingender, J.; Steinmann, E.; Kehrmann, J.; Gulbins, E.; Buer, J.; Rath, P.M.; et al. Effects of green tea compound epigallocatechin-3-gallate against Stenotrophomonas maltophilia infection and biofilm. PLoS ONE. 2014, 9, e92876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heras, B.; Scanlon, M.J.; Martin, J.L. Targeting virulence not viability in the search for future antibacterials. Br. J. Clin. Pharmacol. 2015, 79, 208–215. [Google Scholar] [CrossRef]
- Totsika, M. Disarming pathogens: Benefits and challenges of antimicrobials that target bacterial virulence instead of growth and viability. Future Med. Chem. 2017, 9, 267–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.M.; Wang, F.F.; Zhang, H.; Zhang, Q.; Qiana, W. Genome-wide identification of genes necessary for biofilm formation by nosocomial pathogen Stenotrophomonas maltophilia reveals that orphan response regulator FsnR is a critical modulator. Appl. Environ. Microbiol. 2015, 81, 1200–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhenga, L.; Wang, F.F.; Ren, B.Z.; Liu, W.; Liu, Z.; Qian, W. Systematic mutational analysis of histidine kinase genes in the nosocomial pathogen Stenotrophomonas maltophilia identifies BfmAK systemcontrol of biofilm development. Appl. Environ. Microbiol. 2016, 82, 2444–2456. [Google Scholar] [CrossRef] [Green Version]
- Molloy, K.; Smith, S.G.; Cagney, G.; Dillon, E.T.; Greene, C.M.; McElvaney, N.G. Characterisation of the major extracellular proteases of Stenotrophomonas maltophilia and their effects on pulmonary antiproteases. Pathogens 2019, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- DuMont, A.L.; Karaba, S.M.; Cianciotto, N.P. Type II secretion-dependent degradative and cytotoxic activities mediated by Stenotrophomonas maltophilia serine proteases StmPr1 and StmPr2. Infect. Immun. 2015, 83, 3825–3837. [Google Scholar] [CrossRef] [Green Version]
- DuMont, A.L.; Cianciotto, N.P. Stenotrophomonas maltophilia serine protease StmPr1 induces matrilysis, anoikis, and protease-activated receptor 2 activation in human lung epithelial cells. Infect. Immun. 2017, 85, e00544-17. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.J.; Huang, Y.W.; Lin, Y.T.; Ning, H.C.; Yang, T.C. Inactivation of SmeSyRy two-component regulatory system inversely regulates the expression of SmeYZ and SmeDEF efflux pumps in Stenotrophomonas maltophilia. PLoS ONE 2016, 11, e0160943. [Google Scholar] [CrossRef]
- Lin, Y.T.; Huang, Y.W.; Chen, S.J.; Chang, C.W.; Yang, T.C. The SmeYZ efflux pump of Stenotrophomonas maltophilia contributes to drug resistance, virulence-related characteristics, and virulence in mice. Antimicrob. Agents and Chemother. 2015, 59, 4067–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jusko, W.J. Pharmacokinetics and receptor-mediated pharmacodynamics of corticosteroids. Toxicology 1995, 102, 189–196. [Google Scholar] [CrossRef]
- Di Nocera, P.P.; De Gregorio, E.; Rocco, F. GTAG- and CGTC-tagged palindromic DNA repeats in prokaryotes. BMC genomics 2013, 14, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, F.; De Gregorio, E.; Di Nocera, P.P. A giant family of short palindromic sequences in Stenotrophomonas maltophilia. FEMS Microbiol. Lett. 2010, 308, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pane, K.; Cafaro, V.; Avitabile, A.; Torres, M.T.; Vollaro, A.; De Gregorio, E.; Catania, M.R.; Di Maro, A.; Bosso, A.; Gallo, G. Identification of novel cryptic multifunctional antimicrobial peptides from the human stomach enabled by a computational-experimental platform. ACS Synth. Biol. 2018, 7, 2105. [Google Scholar] [CrossRef] [PubMed]
- Ayaz Ahmed, K.B.; Raman, T.; Veerappan, A. Platinum nanoparticles inhibit bacteria proliferation and rescue zebrafish from bacterial infection. RSC Adv. 2016, 50, 44415–44424. [Google Scholar] [CrossRef]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 5th ed; Lorian, V., Ed.; The Lippincott Williams & Wilkins Co.: Philadelphia, PA, USA, 2005; pp. 365–440. [Google Scholar]
- De Gregorio, E.; Esposito, E.P.; Zarrilli, R.; Di Nocera, P.P. Contact-Dependent Growth Inhibition Proteins in Acinetobacter baylyi ADP1. Curr. Microbiol. 2018, 75, 1434–1440. [Google Scholar] [CrossRef] [Green Version]
- Martinucci, M.; Roscetto, E.; Iula, V.D.; Votsi, A.; Catania, M.R.; De Gregorio, E. Accurate identification of members of the Burkholderia cepacia complex in cystic fibrosis sputum. Lett. Appl. Microbiol. 2016, 62, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | MIC | MBC |
|---|---|---|
| PYED-1 | 256 | 512 |
| 2 (DFZ) | >1000 | >1000 |
| 3 | 750 | >1000 |
| 4 | >1000 | >1000 |
| 5 | 512 | >1000 |
| 6 | >1000 | >1000 |
| 7 | 1000 | >1000 |
| 8 (PD) | >1000 | >1000 |
| STX | 1 | 1 |
| Strain | MIC | MBC | KQ | Strain | MIC | MBC | KQ |
|---|---|---|---|---|---|---|---|
| K279a | 256 | 512 | 2 | Sm0707 | 256 | 512 | 2 |
| Sm0262 | 128 | 512 | 4 | Sm0916 | 256 | 512 | 2 |
| Sm0527 | 64 | 128 | 2 | Sm1053 | 512 | 2000 | 4 |
| Sm0528 | 128 | 256 | 2 | OBGTC3 | 64 | 128 | 2 |
| Sm0545 | 512 | 2000 | 4 | OBGTC9 | 256 | 512 | 2 |
| Sm0571 | 256 | 512 | 2 | OBGTC20 | 128 | 512 | 2 |
| Bacterial strain | Combination | MICa (μg/mL) | MICc (μg/mL) | FIC index |
|---|---|---|---|---|
| S. maltophilia K279a | PYED-1 /gentamicin | 256/16 | 64/4 | 0.5 |
| PYED-1 /amikacin | 256/16 | 64/2 | 0.375 |
| Gene | Fold Change ± SD | P Value | ||
|---|---|---|---|---|
| Smlt0648 | rmlA | glucose-1-phosphate thymidylyl transferase | −1.10 ± 0.045 * | 0.00002919 |
| Smlt0686 | StmPr1 | extracellular protease | −2.21 ± 0.108 * | <0.0000001 |
| Smlt0706 | smf-1 | fimbrial adhesin protein | −1.33± 0.144 * | 0.00003563 |
| Smlt1736 | hfq | host factor-I protein | −1.20± 0.241 | 0.07368922 |
| Smlt2120 | aph3’-IIc | aminoglycoside 3’-phosphotransferase II | −2.93 ± 0.193 * | <0.0000001 |
| Smlt2202 | smeZ | multidrug efflux pump | −3.97 ± 1.381 * | 0.00009759 |
| Smlt2299 | fsnR | response regulator protein | −2.03 ± 0.061 * | <0.0000001 |
| Smlt3524 | sphB | Serine-protease | −2.66 ± 0.067 * | <0.0000001 |
| Smlt3615 | aac6′-Iz | aminoglycoside 6’-N-acetyltransferase | −2.44± 0.486 * | 0.00000015 |
| Smlt3638 | transmembrane hemolysin protein | −1.23 ± 0.083 * | 0.00000143 | |
| Smlt4190 | sppA | protease IV | −1.63 ± 0.088 * | <0.0000001 |
| Smlt4209 | bfmA | two component response regulator | −2.36 ± 0.174 * | <0.0000001 |
| Smlt4395 | StmPr3 | Serine-protease | −2.90 ± 0.456 * | <0.0000001 |
| Primer Name | Primer Sequence |
|---|---|
| aac6-Iz fw | TGTGGACTGATGCCGATG |
| aac6-Iz rv | GCACTTCAGCGAAACCAAC |
| aph3-IIc fw | CCGATCATGAAGACCTGGTG |
| aph3-IIc rv | GTCGATGAAACCGCTGAAAC |
| bfmA fw | AGTGAACTGCGCTTTTCTGG |
| bfmA rv | TGAATTCACCACGGCTGAG |
| fsnR fw | TCCTGATGGACCTGTCATTG |
| fsnR rv | TGCATGGTCATCATCACAAC |
| Hfq fw | TCTACAAGCACGCCATTTCC |
| Hfq rv | TACTCGTCTGCTTCATCACCTG |
| rmlA fw | TGCTGGGTGACAACATCTTC |
| rmlA rv | CCGGATCATTCACCCAATAG |
| rpoB fw | AGGAAATGCTGACGGTGAAG |
| rpoB rv | ACGAGCACGTTGAAGGATTC |
| smeZ fw | GCAGTGATGTACCTGTTTCTGC |
| smeZ rv | CAGCACATTGATCGAGAAGC |
| smf-1 fw | ACCGTGTCCAAGAACACTCTG |
| smf-1 rv | TGCACTTGGTCAGGTTGATG |
| Smlt3638 fw | GGTTGAAGGTATTCGACCACTG |
| Smlt3638 rv | ATCAGGGTGAACGGGGTATAG |
| sphB fw | CGCATCTTTCAGTCACCAAC |
| sphB rv | GTAATTGAAGTTGGCCAGCAC |
| sppA fw | AGTTTCTTCATCGGGCTGTG |
| sppA rv | ATGACGAACATCACCAGCAG |
| StmPr1 fw | GCCGAAGTCATCAACCTCTC |
| StmPr1 rv | ACACGTTGGTGTTGCTGTTG |
| StmPr3 fw | ATCGACAGCACCTGCAACTAC |
| StmPr3 rv | TTCACATCGCGATAGGACAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, A.; Vollaro, A.; Esposito, E.P.; D’Alonzo, D.; Guaragna, A.; Zarrilli, R.; De Gregorio, E. Antibacterial and Antivirulence Activity of Glucocorticoid PYED-1 against Stenotrophomonas maltophilia. Antibiotics 2020, 9, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030105
Esposito A, Vollaro A, Esposito EP, D’Alonzo D, Guaragna A, Zarrilli R, De Gregorio E. Antibacterial and Antivirulence Activity of Glucocorticoid PYED-1 against Stenotrophomonas maltophilia. Antibiotics. 2020; 9(3):105. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030105
Chicago/Turabian StyleEsposito, Anna, Adriana Vollaro, Eliana Pia Esposito, Daniele D’Alonzo, Annalisa Guaragna, Raffaele Zarrilli, and Eliana De Gregorio. 2020. "Antibacterial and Antivirulence Activity of Glucocorticoid PYED-1 against Stenotrophomonas maltophilia" Antibiotics 9, no. 3: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030105