A Novel, Integron-Regulated, Class C β-Lactamase
by

Maria-Elisabeth Böhm
1,2, 
Mohammad Razavi
1,2,
Carl-Fredrik Flach
1,2 and
D. G. Joakim Larsson
1,2,* 1
Centre for Antibiotic Resistance Research (CARe), University of Gothenburg, Gothenburg, Sweden
2
Department of Infectious Diseases, Institute of Biomedicine, Sahlgrenska Academy, University of Gothenburg, 405 30 Gothenburg, Sweden
*
Author to whom correspondence should be addressed.
Antibiotics 2020, 9(3), 123; https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030123
Submission received: 12 February 2020
/
Revised: 10 March 2020
/
Accepted: 11 March 2020
/
Published: 14 March 2020
(This article belongs to the Special Issue Microbial Drug Resistance Genes)
Abstract
:AmpC-type β-lactamases severely impair treatment of many bacterial infections, due to their broad spectrum (they hydrolyze virtually all β-lactams, except fourth-generation cephalosporins and carbapenems) and the increasing incidence of plasmid-mediated versions. The original chromosomal AmpCs are often tightly regulated, and their expression is induced in response to exposure to β-lactams. Regulation of mobile ampC expression is in many cases less controlled, giving rise to constitutively resistant strains with increased potential for development or acquisition of additional resistances. We present here the identification of two integron-encoded ampC genes, blaIDC-1 and blaIDC-2 (integron-derived cephalosporinase), with less than 85% amino acid sequence identity to any previously annotated AmpC. While their resistance pattern identifies them as class C β-lactamases, their low isoelectric point (pI) values make differentiation from other β-lactamases by isoelectric focusing impossible. To the best of our knowledge, this is the first evidence of an ampC gene cassette within a class 1 integron, providing a mobile context with profound potential for transfer and spread into clinics. It also allows bacteria to adapt expression levels, and thus reduce fitness costs, e.g., by cassette-reshuffling. Analyses of public metagenomes, including sewage metagenomes, show that the discovered ampCs are primarily found in Asian countries.
1. Introduction
AmpC-type β-lactamases, also called class C β-lactamases (Ambler structural classification) or class 1 cephalosporinases (functional classification), have gained clinical relevance due to their broad spectrum of activity against penicillins, cephalosporins, cephamycins, and monobactams, while not being inhibited by common β-lactamase inhibitors like clavulanic acid and tazobactam [1,2]. Major clinical concerns are the spread of plasmid-mediated AmpCs, the development of high-level resistance against a broad spectrum of β-lactams by mutations in the regulatory systems that control their expression, and the subsequent accumulation of resistance genes to several antibiotics [3].
After the introduction of later-generation cephalosporins, monobactams, and carbapenems in the 1980s to avoid resistance by known β-lactamases, overproduction of chromosomal AmpC was the first detected broad-spectrum resistance mechanism [3,4]. Nowadays, AmpCs are less prevalent than extended spectrum β-lactamases (ESBLs), but they have a similar substrate spectrum [2,5]. In addition, several AmpC β-lactamases cause resistance to carbapenems above the clinical threshold in a porin-deficient background [6,7,8]. This potential, in combination with the frequent occurrence of AmpCs in multidrug-resistant isolates, is rather worrisome [9,10,11].
AmpCs and other penicillin-binding proteins (not necessarily β-lactamases) are chromosomally encoded in both gram-positive and gram-negative bacteria [3]. Research on their expression and regulation has mostly focused on gram-negatives, where ampC expression normally is tightly regulated in response to β-lactam exposure and cell wall damage [3,12,13]. In many Enterobacteriaceae and in P. aeruginosa, the repressor AmpR, one or several versions of the regulatory enzyme AmpD, and the inner membrane oligopeptide permease AmpG control the inducible expression of ampC [14,15,16]. Different β-lactams induce ampC expression at different levels, and they are to varying degrees vulnerable to hydrolyzation. Penicillins and first-generation cephalosporins are strong inducers, and at the same time very good substrates for AmpCs. Consequently, many later-generation β-lactams are weak inducers and substrates [3,17]. In contrast, cefoxitin and imipenem are both strong inducers, leading to increased resistance against other β-lactams and inhibitors, but are poor substrates themselves (especially imipenem) [17,18]. Production of large quantities of AmpC achieves high-level resistance, even against the poorer substrates, such as cephalosporins and monobactams.
Overexpression or de-repression of chromosomal AmpC is mainly caused by mutations in the genes ampD or ampR, or more rarely by mutations in ampG [3]. In E. coli, A. baumannii, and Shigella, expression is non-inducible, due to the lack of the ampR gene, and ampC expression is instead regulated by promoter and growth rate-dependent attenuation [15,19,20,21]. Mutations in the promoter or attenuator sequence can lead to increased expression [19,22,23]. In Aeromonas, ampC expression was first found to be controlled by the two-component regulatory system BlrAB (β-lactam resistance) [24]. BlrAB-like systems were later shown to be global metabolic regulators—the homolog in E. coli and P. aeruginosa is the CreBC (carbon source responsive) system—that affect cell wall recycling, biofilm formation, and general fitness as well [13,25].
Chromosomally encoded AmpCs have been known for decades in Enterobacteriaceae, before the first plasmid-borne AmpC was reported in 1989 [26]. Since then, plasmid-mediated AmpCs have spread quickly to various bacteria, and often resulted in multi-resistant pathogens [4,27]. Plasmid-encoded AmpCs are now common all over the world; they originate from the chromosomal versions, contain additional regulatory possibilities, and appear both inducible and in a perpetually de-repressed state [11,23]. They have been mobilized by different mechanisms, such as transposons and insertion sequence (IS) elements (either with their original expression regulation [28] or controlled by a promoter provided by the mobile element [29,30,31,32]), and are often associated with integrons [3,4,28,33,34,35]. These mobile elements, e.g., IS26, ISEcp1 or ISCR1, have inserted in close vicinity or within an integron, and form larger composite structures with other transposable elements. However, to the best of our knowledge, ampC genes have not yet been reported as a gene cassette in an integron.
Integrons are genetic elements that acquire, shuffle, and express promoter-less gene cassettes to regulate the expression of a variety of accessory factors, including virulence, secondary metabolism, and antimicrobial or metal resistance genes [36]. Integrons consist of an integrase gene (intI) responsible for the specific excision and integration of gene cassettes at the attI recombination site. Gene cassettes are expressed from the constitutive promoter Pc embedded in intI and the region between intI and attI. The distance from this promoter determines the level of their expression [37]. All gene cassettes include attC, an imperfect inverted repeat, at the 3’ end, which is recognized as target recombination site by the integrase [38]. Class 1 integrons, developed from chromosomal structures, have been mobilized and spread rapidly as a consequence of natural selection and co-selection [39]. A large number of available resistance gene cassettes and the linkage to transposable elements have made this process possible, immensely accelerated by the human use of antibiotics and disinfectants [38,40,41]. The known pool of resistance gene cassettes is comprised of at least 130 different genes [42]. They include a variety of aminoglycoside modifying enzymes, chloramphenicol, quinolone, macrolide, fosfomycin, and trimethoprim resistances, as well as quarternary ammonium compound efflux proteins and β-lactamases. Some class A β-lactamases, like GES, KPC, and BEL, as well as the class D β-lactamase OXA are well known as resistance gene cassettes [43,44,45]. Class B metallo-β-lactamases, like IMP or VIM [46,47], are also abundant in class 1 integron cassettes, but to the best of our knowledge, no class C β-lactamase has been reported in integrons so far.
If class C β-lactamases are indeed present as gene cassettes, they can take advantage of a successful transmission and expression system already harbored by a multitude of mobile elements and bacteria (both pathogenic and non-pathogenic). The ability of integrons to accumulate resistances also favors co-selection, and would enhance the spread of cassette-encoded ampC. To answer the question of whether ampC genes appear as gene cassettes, we searched all sequences encoding potential ampC genes from NCBI’s non-redundant protein database and the metagenomic datasets from a previous study, which have focused specifically on the identification of antibiotic resistance genes in class 1 integrons [48]. We discovered two novel AmpC proteins that are encoded as gene cassettes and verified their functionality in E. coli. Expression under control of a class 1 integron constitutes an additional way to escape tight regulation control and to facilitate fast spread.
2. Results
2.1. A Novel Class C β-lactamase Occurring in Gene Cassettes
To determine the general abundance of ampC gene cassettes, we searched all complete and draft bacterial genomes in NCBI’s assembly database for the 311 known ampC genes from the comprehensive antibiotic resistance database (CARD) [49] with identity greater than 70% and coverage greater than 50%, as thresholds. We then checked the DNA sequences 2.5 kb up- and downstream of the 20,836 resulting open reading frames (ORFs) for integron attachment sites (attC) using IntegronFinder [50]. Only one ampC gene was detected, blaCMY-2 of Salmonella enterica (GenBank: AAGERX010000065.1), with attC sites in close vicinity. A more detailed analysis showed that blaCMY-2 is merely located adjacent to the last cassette of a class 1 integron. Furthermore, blaCMY-2 and the downstream region have appeared in several bacterial genomes without the integron, suggesting that the detected blaCMY-2 is not a gene cassette (Figure S1, Supplementary Materials).
Since no known ampC gene was discovered as a gene cassette in sequenced isolates, we chose to search the functional metagenomics amplicons containing only integron gene cassettes that were selected on cefotaxime [48] for ampC genes with the same method. Preparation of the functional metagenomic amplicons is summarized briefly as follows: DNA was isolated from river sediments contaminated with municipal and hospital sewage. Gene cassettes were amplified from these samples, with primer pairs targeting the gene cassette array of class 1 integrons [48,51]. The amplification products were cloned into the vector pZE21-Pbla and screened for resistance against different antibiotics in E. coli DH10β. The resistant clones were scraped off the selection plates and used as templates to prepare barcoded amplicons for long-read sequencing (PacBio). Any putative or known ampC recovered from the cefotaxime selection reads can be considered mobile and prone to occur in pathogens. This search resulted in 101 ORFs. After assuring that they were complete and flanked by attachment sites, 15 unique but still similar ORFs (at least 94% nucleotide identity) remained, and none of them was a known ampC. Twelve ORFs appeared each on a single PacBio read. They had sporadic mismatches and more than 99% nucleotide similarity to one of the other three ORFs, suggesting that the emergence of these variants was due to possible sequencing errors. Moreover, two of the three abundant ORFs differed consistently by just one nucleotide in all their identified reads. We chose to verify the functionality of one of these almost identical ORFs, as well as the other abundant ORF (the number of unique reads carrying them are listed in Table S1 in the Supplementary Materials). The two selected ORFs share 95% amino acid sequence identity between them, but are less than 85% identical to the closest related sequences that are currently available in the NCBI non-redundant protein database: WP_123086336.1 and WP_095207490.1. Consequently, we considered them a novel family of class C β-lactamases and named them blaIDC-1 and blaIDC-2 (integron-derived cephalosporinase). The two new AmpC variants were not recognized earlier by sequence homology-based analyses of the same metagenomic DNA samples using entire genes as templates [51,52,53], since the identity threshold was too stringent.
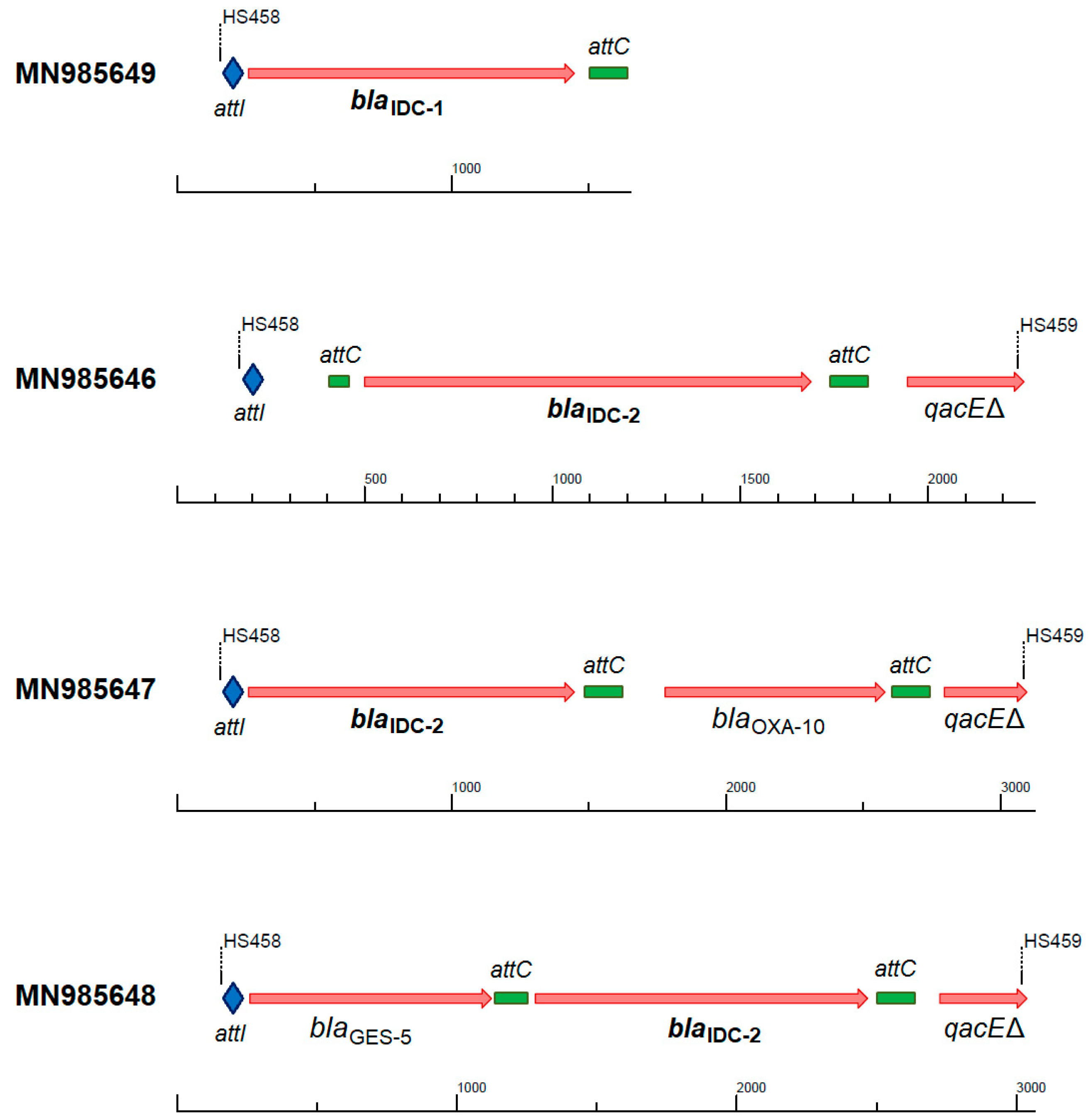
The blaIDC-2 gene cassette was identified in three different contexts in class 1 integrons, while blaIDC-1 always occurred as the first cassette in truncated reads (Figure 1). All three cassette arrays were also successfully identified by IntegronFinder from the amplicon reads. The ampC gene appeared as the first or second cassette, often combined with the class D β-lactamase OXA-10 [54] or the carbapenemase GES-5 [55], which can confer resistance against almost all β-lactam antibiotics to any strain harboring this integron.
2.2. Evidence for Class C β-lactamase Activity: Resistance Profile
Functional screening of the metagenomic libraries included four β-lactam antibiotics (cefotaxime, ertapenem, imipenem, and meropenem [48]). The two identified novel ampC genes were both clearly selected on cefotaxime, but not on carbapenems (Table S1 in the Supplementary Materials), and were assumed to be AmpCs on the basis of sequence similarities. AmpCs have a broad substrate spectrum, but as their expression is variable in clinical isolates, and consequently the corresponding resistance levels, there is no standardized detection method [4,57,58]. We therefore chose to test β-lactamase activity with both the disk diffusion method and Etest for the determination of minimal inhibitory concentration (MIC) values against a selection of different cephalosporins, carbapenems, monobactams, and inhibitors.
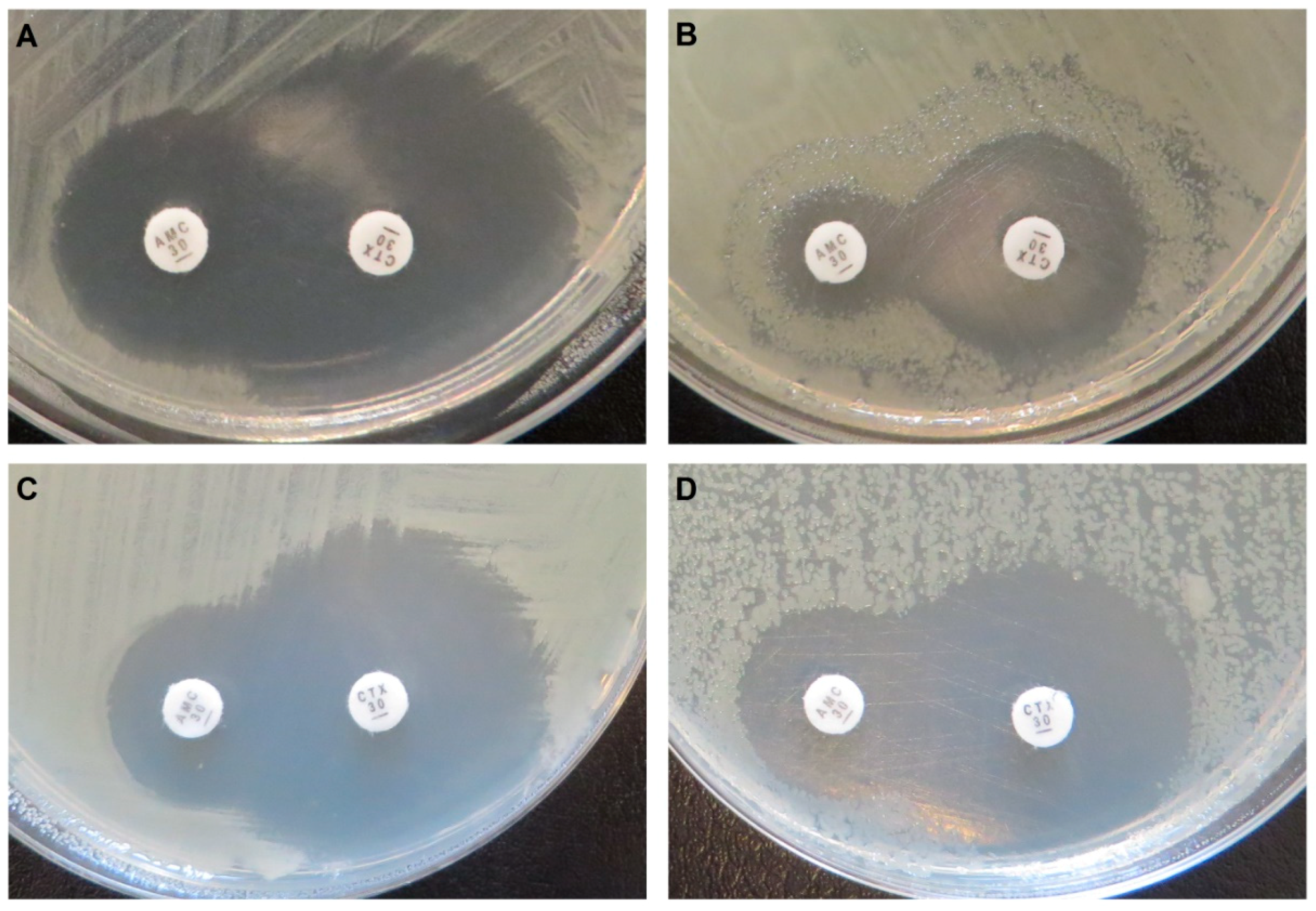
Both novel AmpCs present the typical class C β-lactamase resistance profile (Table 1 and Table 2). Integron-derived cephalosporinase 1 (IDC-1) is active against penicillin, second- and third-generation cephalosporins (cefotaxime, ceftazidime, cefoxitin), and monobactams (aztreonam), but showed no clear activity against fourth-generation cephalosporins (cefepime) and carbapenems. In addition, it is unimpaired by clavulanic acid (Figure 2), but inhibited by cloxacillin [57,59,60]. The overexpression of blaIDC-1 caused a four-fold increase in the ertapenem MIC (Table 2). However, the strain remains susceptible (clinical breakpoint of Enterobacterales = 0.5 µg/mL; EUCAST breakpoint tables v9.0) under the tested conditions. The initial functional selections were carried out at concentrations designed to identify high-level ertapenem resistance, and were therefore too high to detect this small increase (0.032 µg/mL and 1 µg/mL, [48]). Expression of blaIDC-2 resulted in a generally minor increase of resistance against the same antibiotics, and was also not inhibited by clavulanate, but cloxacillin abolished the resistance phenotype. Decreased susceptibility to ceftazidime, amoxicillin, and aztreonam could not be revealed by disk diffusion tests (Table 1). When tested with Etest stripes, blaIDC-2 caused six-fold (aztreonam), four-fold (ceftazidime), and three-fold (amoxicillin) MIC increases, while it showed no activity against cefepime and carbapenems (Table 2).
Most known AmpCs have high isoelectric points [3,4], but the newly discovered AmpCs are predicted to have exceptionally low isoelectric point (pI) values (IDC-1: pI = 5.7, and IDC-2: pI = 5.1), indicating polarity changes compared to known AmpCs. Isoelectric focusing has been used to discern the different classes of β-lactamases, since the majority of AmpC proteins show considerably higher isoelectric points (pI ≥ 8) than class A β-lactamases [3,4,61,62]. The closely related AmpCs LRA-10 and LRA-18 (Figure 3) from the CARD database have predicted pI values of 9.4. Thus, the novel AmpC proteins cannot be distinguished from class A β-lactamases by isoelectric focusing.
2.3. Phylogenetic Affiliation within Class C β-lactamases
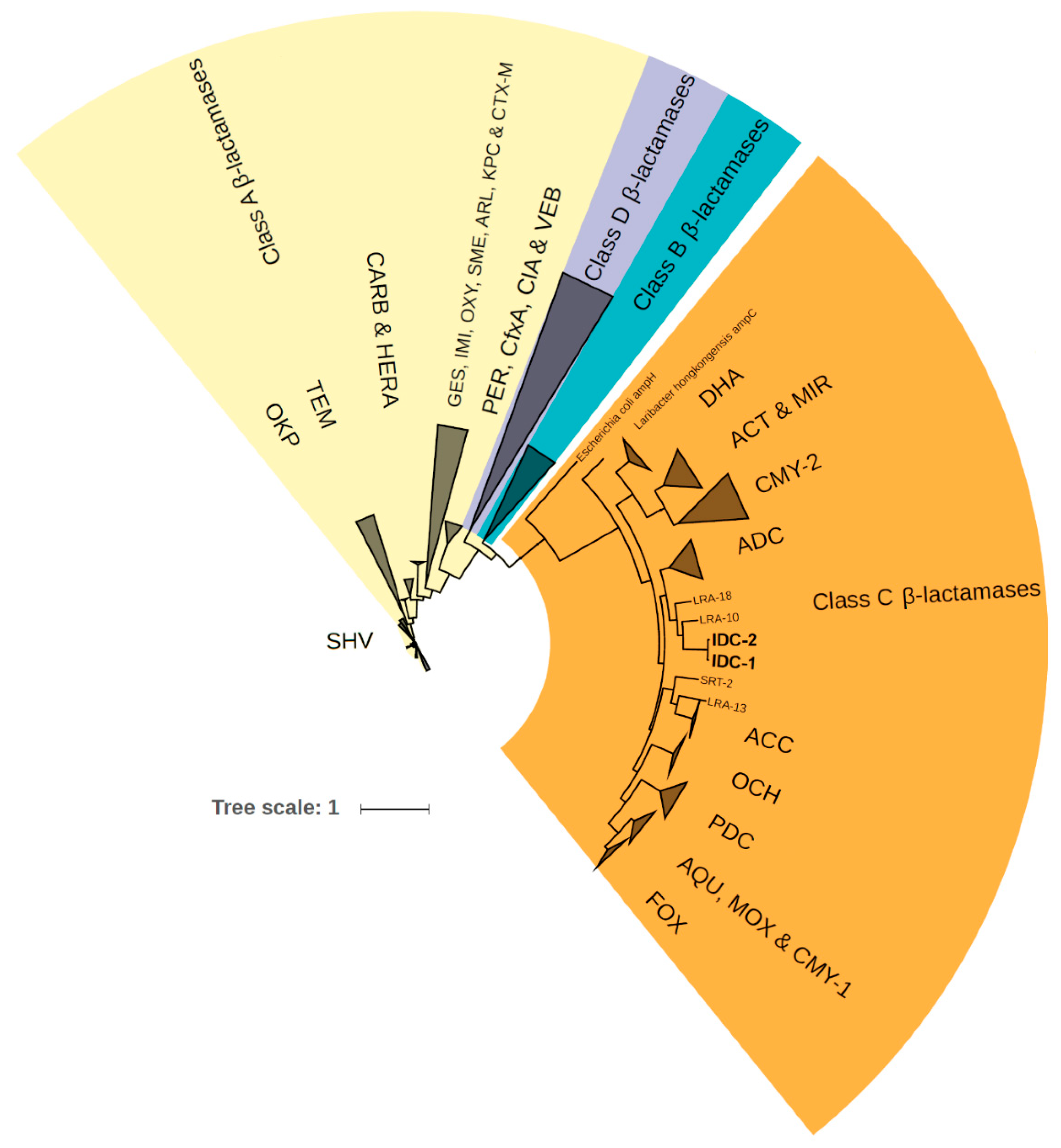
The two novel AmpC variants named IDC-1 and IDC-2 formed a distinct cluster in the phylogenetic tree of β-lactamases from the CARD database (Figure 3), which led us to consider them as a new family of class C β-lactamases. The closest related proteins from the NCBI non-redundant protein database are WP_123086336.1 and WP_095207490.1, which are encoded by chromosomal genes in the Chinese soil isolate Lysobacter sp. ZS60 [63] and Luteimonas sp. JM171 from an Hawaiian coral [64]. These showed 85% and 72% amino acid identity to IDC-1, respectively, and have been annotated as class C β-lactamases, even though their functionalities are not proven. The closest homologous proteins with proven class C β-lactam resistance are LRA-10 and LRA-18. These proteins are encoded by genes recovered from Alaskan soil [65] (Figure 3) and share 55% identity with the two novel AmpCs.
In order to detect the spread of the blaIDC family, 1251 metagenomic datasets were searched for reads indicating their presence (Table S2 in the Supplementary Materials). The search revealed that reads mapping to blaIDC are currently not common, but they appear in metagenomes prepared from wastewaters or river sediments contaminated with urban and pharmaceutical production wastewaters [53,66,67,68,69,70] especially in Asia (India, China, Vietnam, Cambodia). The gene seems to be still rare in the rest of the world.
3. Discussion
Here we show an additional mode of ampC transmission. This is the first description of a class C β-lactamase, the blaIDC family, expressed as gene cassettes under the control of a class 1 integron. The pool of gene cassettes, from which integrons can assemble their cassette array, contains class A, B, and D β-lactamases [42], and is now shown to be comprised of class C β-lactamases, too. The ability of integrons to acquire several resistance gene cassettes [36] provides ample opportunities for co-selection and further spread of a cassette. AmpC gene cassettes discovered in this study occur in combination with blaGES-5 or blaOXA-10, showing the potential for transmission of resistance genes against virtually all β-lactams in a single transfer event. Class 1 integrons are often further embedded into transposable elements and conjugative plasmids that have facilitated their spread into many versatile bacterial species thriving in various geographical and environmental conditions [71]. The ampC cassette is in all likelihood not limited to the contexts identified in this study, since integrons constantly acquire, excise, or shuffle their cassettes to adapt to changing conditions [72].
The two novel ampC gene cassettes are divergent from the known clinical ampC genes, but they are already mobilized and appear in an environment strongly impacted by hospital wastewater [53]. While not (yet) detected in the clinics, they occur in samples contaminated with municipal and hospital wastewater, indicating their potential to spread to human pathogens—or that they are already there but have escaped discovery, as was the case with the gar gene [48]. A systematic search for blaIDC-1 in available metagenomes proved that it is still rare and mainly present in Asian wastewaters (Table S2 in the Supplementary Materials). The integron-borne ampC genes were thus found in environments that both contain pathogens, which might already carry them, and may supply the selective forces necessary to drive their spread into and among human pathogens [73].
The most similar proteins currently available (January 2020) in the public databases show 85% or less identity to IDC-1. Hence, the origin of the novel AmpCs remains elusive. The novel class C β-lactamase genes were therefore named blaIDC-1 and blaIDC-2 (integron-derived cephalosporinase) upon consultation with the NCBI and in agreement with the current β-lactamase nomenclature.
Penicillin-binding proteins and β-lactamases show very different binding affinities and hydrolyzing activities against various β-lactams, despite having similar structures and conserved motifs [3,74,75]. While both IDCs display a resistance pattern consistent with that of an AmpC, IDC-1 causes much higher resistance levels than IDC-2 in an isogenic background. All three conserved sequence motifs necessary for class C β-lactamase activity are identical in IDC-1 and IDC-2 [76,77]. However, 18 amino acid changes were detected outside of the conserved motifs, of which one or several might be responsible for the observed differences in activity. Modelling of the protein structures revealed that IDC-1 contains a positively charged Arg-238 close to the active site cavity, whereas IDC-2 harbors proline at this position (Figure S2, Supplementary Materials). Furthermore, only overexpression of IDC-1 caused a slight increase of resistance against ertapenem. This phenomenon has been previously reported in Enterobacter cloacae, Citrobacter freundii, and some plasmid-mediated AmpCs [27,78,79]. It points to an increased hydrolyzation activity towards carbapenems, but such a slight decrease in carbapenem susceptibility as caused by IDC-1 alone might not be of any clinical relevance. However, IDC-1 could very well (like other AmpCs) show enhanced resistance against an extended spectrum of β-lactams, including carbapenems, when occurring in a membrane porin-deficient background [6,79,80]. Several residues are involved in determining substrate affinity and hydrolyzation activity of class C β-lactamases [79,81]. Mutations in these positions could influence their ability to hydrolyze carbapenems. For example, Asn-346 is conserved in the AmpC proteins CMY-2, ACT-1, and DHA-1 that can confer carbapenem resistance, and was shown to be important for enhanced carbapenem hydrolysis, while Ile-346 occurs in AmpCs that fail to confer carbapenem resistance [82]. Both new AmpC proteins harbor Ile at the corresponding position (here: 374) (Figure S2, Supplementary Materials). The Ala-105 residue in some PDCs from P. aeruginosa also contributes to increased carbapenem resistance [83]. IDC-1 indeed harbors alanine at the corresponding position (here 109) and showed slightly reduced ertapenem susceptibility, in contrast to IDC-2 containing threonine at this position. Therefore, it is possible that the increased carbapenemase activity of IDC-1 is due to Ala-109, and might be further enhanced by a single point mutation at position 374.
Since gene cassettes accumulate within an integron, the combination of ampC with other β-lactamase genes can lead to resistance against (almost) all β-lactams. In this study, we identified three different contexts of the blaIDC-2 gene, while the extended context of blaIDC-1 remained elusive (Figure 1). However, many more contexts are conceivable. GES-5 is an extended-spectrum class A β-lactamase active against penicillins, cephalosporins, cephamycins, and carbapenems; it is inhibited by clavulanate, sulbactam, and tazobactam, and is weakly/not active against ceftazidime and aztreonam [84]. These “gaps” in the resistance profile are covered by class C β-lactamases like IDC, which are not incapacitated by the mentioned inhibitors. OXA-10 enzymes (class D β-lactamases) generally show a narrow spectrum of hydrolysis, but some expanded spectrum variants are known [54,85]. The combination with an AmpC-type enzyme extends the resistance spectrum to all β-lactams except carbapenems. Single amino acid changes or a different genetic context can then complete the spectrum of activity [3,54]. Furthermore, class D β-lactamases hydrolyze cloxacillin [1], an inhibitor of class C enzymes, which leads to prolonged activity of AmpC during therapy.
Hyperproduction of AmpC due to one or several mutations/insertions or mobilization into a different context causes between eight-fold and up to 1000-fold increased expression [3,32,86,87,88,89]. These mechanisms lead to a resistance advantage offered by ampC expression prior to antibiotic exposure, but could be a greater fitness burden for the host in comparison to the more restricted chromosomal ampC expression. Gene cassettes, however, are expressed under the control of a constitutive promoter Pc, which is specific for the gene cassette array [38]. The cassettes’ expression level is not just determined by the strength of the promoter, which can vary up to 30-fold [90,91], but also by the distance from the promoter [37], and can thus be changed by insertion or excision of other gene cassettes. The possible removal or reshuffling of ampC gene cassettes within the integron may provide a less costly control mechanism than the constant de-repression or even overexpression of many other (plasmid-borne) AmpCs. Moreover, gene cassettes could potentially be integrated into all integrons available within a bacterial genome, which would lead to an adaptable expression level. The interplay of many factors determines fitness and hampers our ability to judge if or how fast the integron-regulated ampC can spread. However, considering the enormous success of (mobile) integron-borne antibiotic resistances [41], this mode of ampC transmission has the potential to become more abundant.
4. Materials and Methods
4.1. Detection of ampC Gene Cassettes
All complete and draft bacterial genomes in the NCBI assembly database (downloaded on 15 August 2019, containing 333,456 assembly reports) were searched for known ampC genes in gene cassettes. Reference ampC genes were identified by searching the AmpC serine hydrolase domain (conserved protein domain: PRK11289) against the CARD database [49] (v3.0.5) using Hmmer3.0 [92]. The search resulted in 311 proteins with e-values less than 10−5 (Supplementary File 2). By using Diamond [93] (v0.9.24.125), all genomes with at least one antibiotic resistance gene from the CARD database were extracted; all ORFs on the selected genomes were identified using Prodigal [94] (v2.6.3). Then, the predicted ORFs were searched against the reference AmpC dataset, using Diamond, with identity greater than 70% and coverage greater than 50% as thresholds. These thresholds were chosen to capture all known ampCs and allow for detection of new ampCs, while still limiting noise from penicillin binding proteins that do not confer β-lactam resistance. The DNA sequences ±2.5 kb up- and downstream of the 20,836 resulting ORFs were searched for integron attachment sites (attC) and the integrase gene using IntegronFinder [50].
In addition, the functional metagenomics dataset (NCBI BioProject database PRJNA555822) was searched for presence of ampC as gene cassettes. These amplicons contain gene cassettes amplified with primers specific for class 1 integrons, followed by functional selection for different antibiotic resistances [48]. We focused on the reads recovered from cefotaxime selection plates. First, the ORFs in the selected reads were identified using Prodigal, followed by a search against the reference AmpC dataset applying the aforementioned thresholds. To remove duplicates, the detected ampC were clustered with 100% identity using CD-HIT software [95,96]. To be considered an ampC gene cassette, the ORF needed to be complete and flanked by attachment sites. From the resulting 15 candidates, and according to the rationale described in the results section, two ORFs were chosen for synthesis, functional verification, and determination of a detailed resistance profile. Nucleotide sequences containing the novel ampC variants were deposited in GenBank (accession number MN985649: blaIDC-1; MN985646–MN985648: blaIDC-2 in different gene cassette arrangements; see Figure 1).
4.2. Resistance Profile
The genes blaIDC-1 and blaIDC-2 were synthesized and subcloned into pZE21-MCS1 using KpnI and BamHI restriction sites by GeneArt Gene Synthesis (ThermoFisher Scientific, Regensburg, Germany), as described earlier [97]. The recombinant plasmids were electroporated into E. coli C600Z1 (Expressys, Bammental, Germany) and selected using the kanamycin resistance of pZE21-MCS1. The same strain with the empty vector was used as negative control.
4.2.1. Minimal Inhibitory Concentration Determination
Minimal inhibitory concentrations (MICs) were determined using Etest stripes (bioMérieux SA, Marcy l’Etoile, France) on MH (Mueller Hinton) agar plates supplemented with 50 µg/mL kanamycin, with the addition of 250 ng/ml anhydrotetracycline as an inducer of the PLtetO-1 promoter to ensure maximal expression of the blaIDC genes.
4.2.2. Disk Diffusion Tests
Disk diffusion tests (Oxoid, Hampshire, United Kingdom) were performed according to EUCAST recommendations version 7.0 (2019). MH agar plates were supplemented with 50 µg/mL kanamycin, 250 ng/mL of the inducer anhydrotetracycline, and/or 50 µg/mL cloxacillin (class C β-lactamase inhibitor). Cultures were incubated at 37 °C for 24 h before measuring inhibition zones. Each disk of cefotaxime, ceftazidime, cefoxitin, cefepime, and aztreonam contained 30 µg of the antibiotic. Ertapenem disks contained 10 µg, and the amoxicillin–clavulanate combination disk contained 30 µg of the antibiotic and 10 µg of the inhibitor. Potential inhibition of the β-lactamase by clavulanic acid was controlled in a double disk synergy test by placing the cefotaxime and the amoxicillin–clavulanate combination disk at a distance of 20 mm center to center.
4.3. Metagenome Search
The abundance of the blaIDC family was searched in 1251 public metagenomics datasets, as described in [48]. The reads were mapped with Diamond [93] (v0.9.24.125) to the reference protein IDC-1, with thresholds of 95% identity and an ORF length greater than 20 amino acids to recover both blaIDC-1 and blaIDC-2 containing reads. Integron attachment sites were detected by mapping the short paired-end reads to the gene cassettes from Figure 1 using bowtie2 [98] (v2.2.9), and then manually analyzing the marginal paired-end reads using the Tablet software [99] (v1.19.05).
4.4. Phylogenetic Tree
Amino acid sequences of blaIDC-1, blaIDC-2 and all available 1632 β-lactamases from CARD [49] (v3.0.5) were aligned using MAFFT [100] (v7.310; --maxiterate 1000 --localpair). The phylogenetic tree was calculated by FastTree [101] (v2.1.9) using the maximum likelihood algorithm, Jones–Taylor–Thornton model with 1000 times bootstrap. The full version of the tree is available in Supplementary File 3 (Newick format) and the proteins are listed in Supplementary File 4. The Interactive Tree Of Life (iTOL v4) online tool [102] (https://itol.embl.de; last access: January 2020) was used to prepare the phylogenetic tree for display.
4.5. Amino Acid Sequence Analysis and Protein Model
The EMBOSS pepstats software (https://www.ebi.ac.uk/Tools/seqstats/emboss_pepstats/; last access: November 2019) was used to estimate the isoelectric point (pI) of the AmpC proteins. The signal peptide cleavage site was predicted by the SignalP 5.0 server [103]. The I-TASSER server for protein structure and function prediction was used to create models of the two AmpCs [104,105,106]. The models with the highest confidence scores (C-score IDC-1: 1.00 and IDC-2: 0.85) are shown in Figure S2. PyMOL v2.3.3 (https://pymol.org/2/; last access: January 2020) was used to create the ribbon presentation.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2079-6382/9/3/123/s1, Figure S1: Context and alignment of blaCMY-2 and an adjacent class 1 integron; Figure S2: Amino acid sequence and comparison of the predicted protein structures of IDC-1 and IDC-2; Table S1: Frequencies of unique PacBio long reads (97% identity threshold) that harbor novel ampC genes; Table S2: Occurrence of the newly discovered blaIDC family in 1251 metagenomic datasets. Supplementary File 2: 311 AmpC proteins from CARD. Supplementary File 3: Phylogenetic tree of 1632 β-lactamases plus IDC-1 and IDC-2 in Newick format. Supplementary File 4: List of the 1632 β-lactamases from CARD.
Author Contributions
Conceptualization, M.-E.B., M.R., C.-F.F., and D.G.J.L.; data curation, M.R.; funding acquisition, D.G.J.L.; investigation, M.-E.B.; resources, D.G.J.L.; software, M.R.; supervision, D.G.J.L.; validation, M.-E.B., and M.R.; visualization, M.B. and M.R.; writing—original draft, M.-E.B.; writing—review and editing, M.-E.B., M.R., C.-F.F., and D.G.J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Swedish Research Council for Environment, Agriculture and Spatial Planning (FORMAS) (2018-00787 to D.G.J.L), the Swedish Research Council (VR) (2018-02835 and 2018-05771 to D.G.J.L), and the Centre for Antibiotic Resistance Research (CARe) at the University of Gothenburg.
Acknowledgments
We thank the NCBI for assistance with choosing a suitable name for the novel gene family within the established β-lactamase nomenclature. NCBI kindly takes responsibility for future allele designation of the blaIDC family.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bush, K.; Jacoby, G.A. Updated functional classification of β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Thomson, K.S. Extended-Spectrum-β-Lactamase, AmpC, and Carbapenemase Issues. J. Clin. Microbiol. 2010, 48, 1019–1025. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. AmpC β-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-determined AmpC-type β-lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuland, E.A.; Halaby, T.; Hays, J.P.; de Jongh, D.M.C.; Snetselaar, H.D.R.; van Keulen, M.; Elders, P.J.M.; Savelkoul, P.H.M.; Vandenbroucke-Grauls, C.M.J.E.; Al Naiemi, N. Plasmid-mediated AmpC: Prevalence in community-acquired isolates in Amsterdam, The Netherlands, and risk factors for carriage. PLoS ONE 2015, 10, e0113033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, G.A.; Mills, D.M.; Chow, N. Role of β-lactamases and porins in resistance to ertapenem and other β-lactams in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 3203–3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapleton, P.D.; Shannon, K.P.; French, G.L. Carbapenem resistance in Escherichia coli associated with plasmid-determined CMY-4 β-lactamase production and loss of an outer membrane protein. Antimicrob. Agents Chemother. 1999, 43, 1206–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, P.A.; Urban, C.; Mariano, N.; Projan, S.J.; Rahal, J.J.; Bush, K. Imipenem resistance in Klebsiella pneumoniae is associated with the combination of ACT-1, a plasmid-mediated AmpC β-lactamase, and the loss of an outer membrane protein. Antimicrob. Agents Chemother. 1997, 41, 563–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64 (Suppl. 1), i29–i36. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Kawahara, R.; Harada, K.; Teruya, S.; Nakayama, T.; Motooka, D.; Nakamura, S.; Nguyen, P.D.; Kumeda, Y.; Van Dang, C.; et al. The presence of colistin resistance gene mcr-1 and -3 in ESBL producing Escherichia coli isolated from food in Ho Chi Minh City, Vietnam. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Hennequin, C.; Ravet, V.; Robin, F. Plasmids carrying DHA-1 β-lactamases. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.W.; Fisher, J.F.; Mobashery, S. Bacterial cell-wall recycling. Ann. N. Y. Acad. Sci. 2013, 1277, 54–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Lin, J. β-lactamase induction and cell wall metabolism in Gram-negative bacteria. Front. Microbiol. 2013, 4, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadlamani, G.; Thomas, M.D.; Patel, T.R.; Donald, L.J.; Reeve, T.M.; Stetefeld, J.; Standing, K.G.; Vocadlo, D.J.; Mark, B.L. The β-lactamase gene regulator AmpR is a tetramer that recognizes and binds the D-Ala-D-Ala motif of its repressor UDP-N-acetylmuramic acid (MurNAc)-pentapeptide. J. Biol. Chem. 2015, 290, 2630–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honoré, N.; Nicolas, M.H.; Cole, S.T. Inducible cephalosporinase production in clinical isolates of Enterobacter cloacae is controlled by a regulatory gene that has been deleted from Escherichia coli. EMBO J. 1986, 5, 3709–3714. [Google Scholar] [CrossRef] [PubMed]
- Moya, B.; Juan, C.; Albertí, S.; Pérez, J.L.; Oliver, A. Benefit of having multiple ampD genes for acquiring β-lactam resistance without losing fitness and virulence in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2008, 52, 3694–3700. [Google Scholar] [CrossRef] [Green Version]
- Galleni, M.; Amicosante, G.; Frère, J.M. A survey of the kinetic parameters of class C β-lactamases: Cephalosporins and other β-lactam compounds. Biochem. J. 1988, 255, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M. Clinical significance of beta-lactamase induction and stable derepression in gram-negative rods. Eur. J. Clin. Microbiol. 1987, 6, 439–445. [Google Scholar] [CrossRef]
- Jaurin, B.; Grundström, T.; Edlund, T.; Normark, S. The E. coli β-lactamase attenuator mediates growth rate-dependent regulation. Nature 1981, 290, 221–225. [Google Scholar] [CrossRef]
- Bou, G.; Martínez-Beltrán, J. Cloning, nucleotide sequencing, and analysis of the gene encoding an AmpC β-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2000, 44, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Mulvey, M.R.; Bryce, E.; Boyd, D.A.; Ofner-Agostini, M.; Land, A.M.; Simor, A.E.; Paton, S. Molecular characterization of cefoxitin-resistant Escherichia coli from Canadian hospitals. Antimicrob. Agents Chemother. 2005, 49, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corvec, S.; Caroff, N.; Espaze, E.; Marraillac, J.; Reynaud, A. 11 Mutation in the ampC promoter increasing resistance to β-lactams in a clinical Escherichia coli strain. Antimicrob. Agents Chemother. 2002, 46, 3265–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisbig, M.D.; Hossain, A.; Hanson, N.D. Factors influencing gene expression and resistance for Gram-negative organisms expressing plasmid-encoded ampC genes of Enterobacter origin. J. Antimicrob. Chemother. 2003, 51, 1141–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alksne, L.E.; Rasmussen, B.A. Expression of the AsbA1, OXA-12, and AsbM1 β-lactamases in Aeromonas jandaei AER 14 is coordinated by a two-component regulon. J. Bacteriol. 1997, 179, 2006–2013. [Google Scholar] [CrossRef] [Green Version]
- Zamorano, L.; Moyà, B.; Juan, C.; Mulet, X.; Blázquez, J.; Oliver, A. The Pseudomonas aeruginosa CreBC two-component system plays a major role in the response to β-lactams, fitness, biofilm growth, and global regulation. Antimicrob. Agents Chemother. 2014, 58, 5084–5095. [Google Scholar] [CrossRef] [Green Version]
- Bauernfeind, A.; Chong, Y.; Schweighart, S. Extended broad spectrum β-lactamase in Klebsiella pneumoniae including resistance to cephamycins. Infection 1989, 17, 316–321. [Google Scholar] [CrossRef]
- Thomson, K.S.; Smith Moland, E. Version 2000: The new β-lactamases of Gram-negative bacteria at the dawn of the new millennium. Microbes Infect. 2000, 2, 1225–1235. [Google Scholar] [CrossRef]
- Verdet, C.; Benzerara, Y.; Gautier, V.; Adam, O.; Ould-Hocine, Z.; Arlet, G. Emergence of DHA-1-producing Klebsiella spp. in the Parisian region: Genetic Organization of the ampC and ampR genes originating from Morganella morganii. Antimicrob. Agents Chemother. 2006, 50, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Zamorano, L.; Miró, E.; Juan, C.; Gómez, L.; Bou, G.; González-López, J.J.; Martínez-Martínez, L.; Aracil, B.; Conejo, M.C.; Oliver, A.; et al. Mobile genetic elements related to the diffusion of plasmid-mediated AmpC β-lactamases or carbapenemases from Enterobacteriaceae: Findings from a multicenter study in Spain. Antimicrob. Agents Chemother. 2015, 59, 5260–5266. [Google Scholar] [CrossRef] [Green Version]
- Toleman, M.A.; Walsh, T.R. Combinatorial events of insertion sequences and ICE in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 912–935. [Google Scholar] [CrossRef] [Green Version]
- Mollet, B.; Iida, S.; Arber, W. Gene organization and target specificity of the prokaryotic mobile genetic element IS26. Mol. Gen. Genet. 1985, 201, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Jaurin, B.; Normark, S. Insertion of IS2 creates a novel ampC promoter in Escherichia coli. Cell 1983, 32, 809–816. [Google Scholar] [CrossRef]
- Nadjar, D.; Rouveau, M.; Verdet, C.; Donay, L.; Herrmann, J.; Lagrange, P.H.; Philippon, A.; Arlet, G. Outbreak of Klebsiella pneumoniae producing transferable AmpC-type β-lactamase (ACC-1) originating from Hafnia alvei. FEMS Microbiol. Lett. 2000, 187, 35–40. [Google Scholar] [CrossRef]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR elements: Novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 2006, 70, 296–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Moreno, M.O.; Estepa, V.; Sáenz, Y.; Cortell-Ortolá, M.; Fort-Gallifa, I.; Ruiz, J.; Torres, C. Intrahospitalary dissemination of Klebsiella pneumoniae carrying blaDHA-1 and qnrB4 genes within a novel complex class 1 integron. Diagn. Microbiol. Infect. Dis. 2012, 73, 210–211. [Google Scholar] [CrossRef] [PubMed]
- Cambray, G.; Guerout, A.-M.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Collis, C.M.; Hall, R.M. Expression of antibiotic resistance genes in the integrated cassettes of integrons. Antimicrob. Agents Chemother. 1995, 39, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Chow, L.; Asher, A.J.; Waldron, L.S.; Gillings, M.R. Evolution of class 1 integrons: Mobilization and dispersal via food-borne bacteria. PLoS ONE 2017, 12, e0179169. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [Green Version]
- Gillings, M.R. DNA as a Pollutant: The Clinical Class 1 Integron. Curr. Pollut. Rep. 2018, 4, 49–55. [Google Scholar] [CrossRef]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatica, J.; Tripathi, V.; Green, S.; Manaia, C.M.; Berendonk, T.; Cacace, D.; Merlin, C.; Kreuzinger, N.; Schwartz, T.; Fatta-Kassinos, D.; et al. High Throughput Analysis of Integron Gene Cassettes in Wastewater Environments. Environ. Sci. Technol. 2016, 50, 11825–11836. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhu, J.; Zhou, K.; Chen, J.; Yin, Z.; Feng, J.; Ma, L.; Zhou, D. Genetic characterization of novel class 1 Integrons In0, In1069 and In1287 to In1290, and the inference of In1069-associated integron evolution in Enterobacteriaceae. Antimicrob. Resist. Infect. Control 2017, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouheraoua, N.; Poirel, L.; Bourreau, B.; Bonnin, R.; Laroche, L.; Naas, T.; Nordmann, P. Integrase-Mediated Recombination of the bel-1 Gene Cassette Encoding the Extended-Spectrum β-Lactamase BEL-1. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleman, M.A.; Biedenbach, D.; Bennett, D.; Jones, R.N.; Walsh, T.R. Genetic characterization of a novel metallo-β-lactamase gene, blaIMP-13, harboured by a novel Tn5051-type transposon disseminating carbapenemase genes in Europe: Report from the SENTRY worldwide antimicrobial surveillance programme. J. Antimicrob. Chemother. 2003, 52, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kim, M.-N.; Park, K.S.; Lee, J.H.; Karim, A.M.; Park, M.; Kim, J.H.; Lee, S.H. Complex Class 1 Integron Carrying qnrB62 and blaVIM-2 in a Citrobacter freundii Clinical Isolate. Antimicrob. Agents Chemother. 2016, 60, 6937–6940. [Google Scholar] [CrossRef] [Green Version]
- Böhm, M.-E.; Razavi, M.; Marathe, N.P.; Flach, C.-F.; Larsson, D.G.J. Discovery of a novel integron-borne aminoglycoside resistance gene present in clinical pathogens by screening environmental bacterial communities. Microbiome 2020. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Cury, J.; Jové, T.; Touchon, M.; Néron, B.; Rocha, E.P. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef] [Green Version]
- Razavi, M.; Marathe, N.P.; Gillings, M.R.; Flach, C.-F.; Kristiansson, E.; Joakim Larsson, D.G. Discovery of the fourth mobile sulfonamide resistance gene. Microbiome 2017, 5, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marathe, N.P.; Janzon, A.; Kotsakis, S.D.; Flach, C.-F.; Razavi, M.; Berglund, F.; Kristiansson, E.; Larsson, D.G.J. Functional metagenomics reveals a novel carbapenem-hydrolyzing mobile β-lactamase from Indian river sediments contaminated with antibiotic production waste. Environ. Int. 2018, 112, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Marathe, N.P.; Pal, C.; Gaikwad, S.S.; Jonsson, V.; Kristiansson, E.; Larsson, D.G.J. Untreated urban waste contaminates Indian river sediments with resistance genes to last resort antibiotics. Water Res. 2017, 124, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Antunes, N.T.; Lamoureaux, T.L.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Class D β-lactamases: Are They All Carbapenemases? Antimicrob. Agents Chemother. 2014, 58, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, I.K.; Lee, Y.-N.; Jeong, S.H.; Hong, S.G.; Lee, J.H.; Lee, S.H.; Kim, H.J.; Youn, H. Genetic and biochemical characterization of GES-5, an extended-spectrum class A β-lactamase from Klebsiella pneumoniae. Diagn. Microbiol. Infect. Dis. 2007, 58, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.J.; Gillings, M.R.; Nield, B.S.; Mabbutt, B.C.; Nevalainen, K.M.H.; Stokes, H.W. The gene cassette metagenome is a basic resource for bacterial genome evolution. Environ. Microbiol. 2003, 5, 383–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, P.R.; Inglis, T.J.J.; Vanzetti, T.R.; Henderson, B.A.; Harnett, G.B.; Murray, R.J. Comparison of methods for AmpC β-lactamase detection in Enterobacteriaceae. J. Med. Microbiol. 2011, 60, 715–721. [Google Scholar] [CrossRef]
- Tan, T.Y.; Ng, L.S.Y.; He, J.; Koh, T.H.; Hsu, L.Y. Evaluation of screening methods to detect plasmid-mediated AmpC in Escherichia coli, Klebsiella pneumoniae, and Proteus mirabilis. Antimicrob. Agents Chemother. 2009, 53, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Drieux, L.; Brossier, F.; Sougakoff, W.; Jarlier, V. Phenotypic detection of extended-spectrum β-lactamase production in Enterobacteriaceae: Review and bench guide. Clin. Microbiol. Infect. 2008, 14 (Suppl. 1), 90–103. [Google Scholar] [CrossRef] [Green Version]
- Polsfuss, S.; Bloemberg, G.V.; Giger, J.; Meyer, V.; Böttger, E.C.; Hombach, M. Practical approach for reliable detection of AmpC β-lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2011, 49, 2798–2803. [Google Scholar] [CrossRef] [Green Version]
- Coudron, P.E.; Moland, E.S.; Thomson, K.S. Occurrence and detection of AmpC β-lactamases among Escherichia coli, Klebsiella pneumoniae, and Proteus mirabilis isolates at a veterans medical center. J. Clin. Microbiol. 2000, 38, 1791–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Catry, B.; Herman, L.; Haesebrouck, F.; Butaye, P. Diversity of extended-spectrum β-lactamases and class C β-lactamases among cloacal Escherichia coli Isolates in Belgian broiler farms. Antimicrob. Agents Chemother. 2008, 52, 1238–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Dong, H.; Zhou, M.; Huang, Y.; Zhang, H.; He, W.; Sheng, H.; An, L. Lysobacter psychrotolerans sp. nov., isolated from soil in the Tianshan Mountains, Xinjiang, China. Int. J. Syst. Evol. Microbiol. 2019, 69, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Miller, J.M.; Rowley, S.J.; Hou, S.; Donachie, S.P. Draft Genome Sequence of a Novel Luteimonas sp. Strain from Coral Mucus, Hawai’i. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Allen, H.K.; Moe, L.A.; Rodbumrer, J.; Gaarder, A.; Handelsman, J. Functional metagenomics reveals diverse β-lactamases in a remote Alaskan soil. ISME J. 2009, 3, 243–251. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Marathe, N.P.; Berglund, F.; Razavi, M.; Pal, C.; Dröge, J.; Samant, S.; Kristiansson, E.; Larsson, D.G.J. Sewage effluent from an Indian hospital harbors novel carbapenemases and integron-borne antibiotic resistance genes. Microbiome 2019, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Kristiansson, E.; Fick, J.; Janzon, A.; Grabic, R.; Rutgersson, C.; Weijdegård, B.; Söderström, H.; Larsson, D.G.J. Pyrosequencing of antibiotic-contaminated river sediments reveals high levels of resistance and gene transfer elements. PLoS ONE 2011, 6, e17038. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. The structure and diversity of human, animal and environmental resistomes. Microbiome 2016, 4, 54. [Google Scholar] [CrossRef] [Green Version]
- Ibarbalz, F.M.; Orellana, E.; Figuerola, E.L.M.; Erijman, L. Shotgun Metagenomic Profiles Have a High Capacity to Discriminate Samples of Activated Sludge According to Wastewater Type. Appl. Environ. Microbiol. 2016, 82, 5186–5196. [Google Scholar] [CrossRef] [Green Version]
- Gillings, M.R. Class 1 integrons as invasive species. Curr. Opin. Microbiol. 2017, 38, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Escudero, J.A.; Loot, C.; Nivina, A.; Mazel, D. The Integron: Adaptation on Demand. Microbiol. Spect. 2015, 3, 139–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2017, 42. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.R.; Moews, P.C.; Frere, J.M. Molecular evolution of bacterial β-lactam resistance. Chem. Biol. 1996, 3, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Joris, B.; Ledent, P.; Dideberg, O.; Fonzé, E.; Lamotte-Brasseur, J.; Kelly, J.A.; Ghuysen, J.M.; Frère, J.M. Comparison of the sequences of class A β-lactamases and of the secondary structure elements of penicillin-recognizing proteins. Antimicrob. Agents Chemother. 1991, 35, 2294–2301. [Google Scholar] [CrossRef] [Green Version]
- Pozzi, C.; Di Pisa, F.; De Luca, F.; Benvenuti, M.; Docquier, J.D.; Mangani, S. Atomic-Resolution Structure of a Class C β-Lactamase and Its Complex with Avibactam. ChemMedChem 2018, 13, 1437–1446. [Google Scholar] [CrossRef]
- Feller, G.; Zekhnini, Z.; Lamotte-Brasseur, J.; Gerday, C. Enzymes from cold-adapted microorganisms. The class C β-lactamase from the antarctic psychrophile Psychrobacter immobilis A5. Eur. J. Biochem. 1997, 244, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Oakton, K.J.; Carter, M.W.; Warner, M. Activity of ertapenem (MK-0826) versus Enterobacteriaceae with potent β-lactamases. Antimicrob. Agents Chemother. 2001, 45, 2831–2837. [Google Scholar] [CrossRef] [Green Version]
- Mammeri, H.; Guillon, H.; Eb, F.; Nordmann, P. Phenotypic and biochemical comparison of the carbapenem-hydrolyzing activities of five plasmid-borne AmpC β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 4556–4560. [Google Scholar] [CrossRef] [Green Version]
- Mainardi, J.L.; Mugnier, P.; Coutrot, A.; Buu-Hoï, A.; Collatz, E.; Gutmann, L. Carbapenem resistance in a clinical isolate of Citrobacter freundii. Antimicrob. Agents Chemother. 1997, 41, 2352–2354. [Google Scholar] [CrossRef] [Green Version]
- Mammeri, H.; Poirel, L.; Nordmann, P. Extension of the hydrolysis spectrum of AmpC β-lactamase of Escherichia coli due to amino acid insertion in the H-10 helix. J. Antimicrob. Chemother. 2007, 60, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Dahyot, S.; Broutin, I.; de Champs, C.; Guillon, H.; Mammeri, H. Contribution of asparagine 346 residue to the carbapenemase activity of CMY-2 β-lactamase. FEMS Microbiol. Lett. 2013, 345, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Martínez, J.-M.; Poirel, L.; Nordmann, P. Extended-spectrum cephalosporinases in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2009, 53, 1766–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsakis, S.D.; Miriagou, V.; Tzelepi, E.; Tzouvelekis, L.S. Comparative biochemical and computational study of the role of naturally occurring mutations at Ambler positions 104 and 170 in GES β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 4864–4871. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [Green Version]
- Juan, C.; Maciá, M.D.; Gutiérrez, O.; Vidal, C.; Pérez, J.L.; Oliver, A. Molecular Mechanisms of β-Lactam Resistance Mediated by AmpC Hyperproduction in Pseudomonas aeruginosa Clinical Strains. Antimicrob. Agents Chemother. 2005, 49, 4733–4738. [Google Scholar] [CrossRef] [Green Version]
- Juan, C.; Moyá, B.; Pérez, J.L.; Oliver, A. Stepwise Upregulation of the Pseudomonas aeruginosa Chromosomal Cephalosporinase Conferring High-Level β-Lactam Resistance Involves Three AmpD Homologues. Antimicrob. Agents Chemother. 2006, 50, 1780–1787. [Google Scholar] [CrossRef] [Green Version]
- Tracz, D.M.; Boyd, D.A.; Hizon, R.; Bryce, E.; McGeer, A.; Ofner-Agostini, M.; Simor, A.E.; Paton, S.; Mulvey, M.R. Canadian Nosocomial Infection Surveillance Program ampC gene expression in promoter mutants of cefoxitin-resistant Escherichia coli clinical isolates. FEMS Microbiol. Lett. 2007, 270, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Corvec, S.; Caroff, N.; Espaze, E.; Giraudeau, C.; Drugeon, H.; Reynaud, A. AmpC cephalosporinase hyperproduction in Acinetobacter baumannii clinical strains. J. Antimicrob. Chemother. 2003, 52, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Jové, T.; Da Re, S.; Denis, F.; Mazel, D.; Ploy, M.-C. Inverse correlation between promoter strength and excision activity in class 1 integrons. PLoS Genet. 2010, 6, e1000793. [Google Scholar] [CrossRef] [Green Version]
- Lévesque, C.; Brassard, S.; Lapointe, J.; Roy, P.H. Diversity and relative strength of tandem promoters for the antibiotic-resistance genes of several integrons. Gene 1994, 142, 49–54. [Google Scholar] [CrossRef]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.-L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Flach, C.-F.; Boulund, F.; Kristiansson, E.; Larsson, D.J. Functional verification of computationally predicted qnr genes. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Milne, I.; Bayer, M.; Cardle, L.; Shaw, P.; Stephen, G.; Wright, F.; Marshall, D. Tablet-next generation sequence assembly visualization. Bioinformatics 2010, 26, 401–402. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Genetic context and nucleotide accession numbers of the blaIDC gene cassettes recovered from the amplified functional metagenomics dataset of class 1 integrons (primer pair HS458-HS459) [48,56]. Attachment sites: attI marked as blue rhomb, attC marked as green rectangle; blaIDC-1 and blaIDC-2: novel integron-derived cephalosporinases; blaOXA-10: class D β-lactamase OXA-10; blaGES-5: class A β-lactamase GES-5 (carbapenemase); qacEΔ: quarternary ammonium compound resistance protein (truncated).
Figure 1.
Genetic context and nucleotide accession numbers of the blaIDC gene cassettes recovered from the amplified functional metagenomics dataset of class 1 integrons (primer pair HS458-HS459) [48,56]. Attachment sites: attI marked as blue rhomb, attC marked as green rectangle; blaIDC-1 and blaIDC-2: novel integron-derived cephalosporinases; blaOXA-10: class D β-lactamase OXA-10; blaGES-5: class A β-lactamase GES-5 (carbapenemase); qacEΔ: quarternary ammonium compound resistance protein (truncated).

Figure 2.
Disk diffusion tests. E. coli C600Z1 pZE21-blaIDC-1 with expression not induced (A) and induced (B), and E. coli C600Z1 pZE21-blaIDC-2 with expression not induced (C) and induced (D). AMC: amoxicillin + clavulanic acid; CTX: cefotaxime. The tests show that the activity of IDC-1 and IDC-2 is not inhibited by clavulanic acid.
Figure 2.
Disk diffusion tests. E. coli C600Z1 pZE21-blaIDC-1 with expression not induced (A) and induced (B), and E. coli C600Z1 pZE21-blaIDC-2 with expression not induced (C) and induced (D). AMC: amoxicillin + clavulanic acid; CTX: cefotaxime. The tests show that the activity of IDC-1 and IDC-2 is not inhibited by clavulanic acid.

Figure 3.
Phylogenetic tree of all four β-lactamase classes and the two newly discovered integron-derived AmpC variants IDC-1 and IDC-2.
Figure 3.
Phylogenetic tree of all four β-lactamase classes and the two newly discovered integron-derived AmpC variants IDC-1 and IDC-2.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Resistance profile of blaIDC-1 and blaIDC-2 (disk diffusion).
| β-Lactam | Empty Vector | blaIDC-1 | blaIDC-2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cloxacillin | Cloxacillin | cloxacillin | ||||||||||
| Not Induced | Induced | Not Induced | Induced | Not Induced | Induced | Not Induced | Induced | Not Induced | Induced | Not Induced | Induced | |
| Amoxicillin + clavulanate | 21 | 22 | n.d. | n.d. | 24 | 13 | n.d. | n.d. | 21 | 20 | n.d. | n.d. |
| Cefotaxime | 35 | 35 | 36 | 37 | 38 | 24 | 38 | 36 | 35 | 32 | 35 | 43 |
| Ceftazidime | 32 | 31 | 32 | 33 | 32 | 27 | 34 | 35 | 31 | 32 | 32 | 41 |
| Cefoxitin | 27 | 29 | 31 | 31 | 28 | 17 | 32 | 29 | 28 | 25 | 31 | 29 |
| Cefepime | 33 | 33 | 34 | 34 | 36 | 37 | 36 | 37 | 33 | 43 | 35 | 44 |
| Ertapenem | 35 | 34 | 36 | 36 | 38 | 38 | 37 | 38 | 33 | 41 | 35 | 46 |
| Aztreonam | 32 | 32 | 35 | 35 | 36 | 30 | 37 | 37 | 33 | 36 | 34 | 43 |
Inhibition zone sizes are in millimeters; the increased resistance phenotype is underlined; n.d. = not determined. The expression of blaIDC-1 and blaIDC-2 was induced by supplementing the agar with anhydrotetracycline. Cloxacillin is an inhibitor of class C β-lactamases, and is shown to impair the resistance conferred by both IDCs.
Table 2.
Resistance profile of blaIDC-1 and blaIDC-2 (Etest).
| β-Lactam | Empty Vector | blaIDC-1 | blaIDC-2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Not Induced | Induced | MIC Increase (Fold-Change) | Not Induced | Induced | MIC increase (Fold-Change) | Not Induced | Induced | MIC Increase (Fold-Change) | |
| Amoxicillin | 6 | 6 | - | 6 | 48 | 8 | 6 | 16 | 3 |
| Cefotaxime | 0.047 | 0.047 | - | 0.064 | 8 | 125 | 0.032 | 0.75 | 23 |
| Ceftazidime | 0.064 | 0.094 | - | 0.094 | 1.5 | 16 | 0.094 | 0.38 | 4 |
| Cefoxitin | 3 | 3 | - | 2 | 16 | 8 | 3 | 8 | 3 |
| Cefepime | 0.032 | 0.032 | - | 0.023 | 0.032 | - | 0.023 | 0.032 | - |
| Ertapenem | 0.003 | 0.003 | - | 0.004 | 0.016 | 4 | 0.003 | 0.004 | - |
| Imipenem | 0.50 | 0.50 | - | 0.38 | 0.50 | - | 0.38 | 0.38 | - |
| Aztreonam | 0.094 | 0.094 | - | 0.094 | 1.5 | 16 | 0.064 | 0.38 | 6 |
Minimal inhibitory concentration (MIC) values are in µg/ml. The expression of blaIDC-1 and blaIDC-2 was induced by supplementing the agar with anhydrotetracycline.The increased resistance phenotype is underlined.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Böhm, M.-E.; Razavi, M.; Flach, C.-F.; Larsson, D.G.J. A Novel, Integron-Regulated, Class C β-Lactamase. Antibiotics 2020, 9, 123. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030123
AMA Style
Böhm M-E, Razavi M, Flach C-F, Larsson DGJ. A Novel, Integron-Regulated, Class C β-Lactamase. Antibiotics. 2020; 9(3):123. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030123
Chicago/Turabian StyleBöhm, Maria-Elisabeth, Mohammad Razavi, Carl-Fredrik Flach, and D. G. Joakim Larsson. 2020. "A Novel, Integron-Regulated, Class C β-Lactamase" Antibiotics 9, no. 3: 123. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030123
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.