Natural Oregano Essential Oil May Replace Antibiotics in Lamb Diets: Effects on Meat Quality

 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
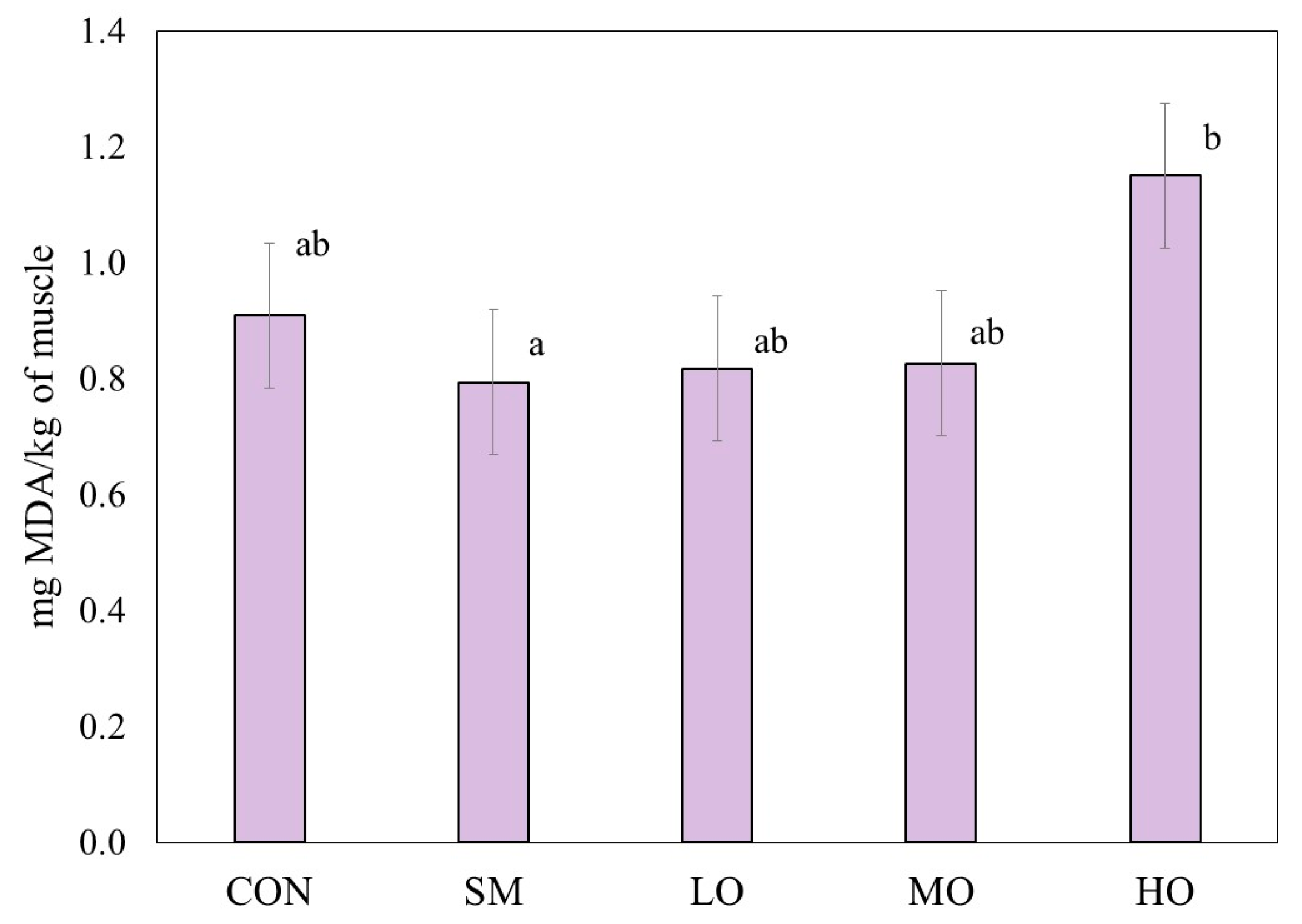
2.1. Lipid Oxidation
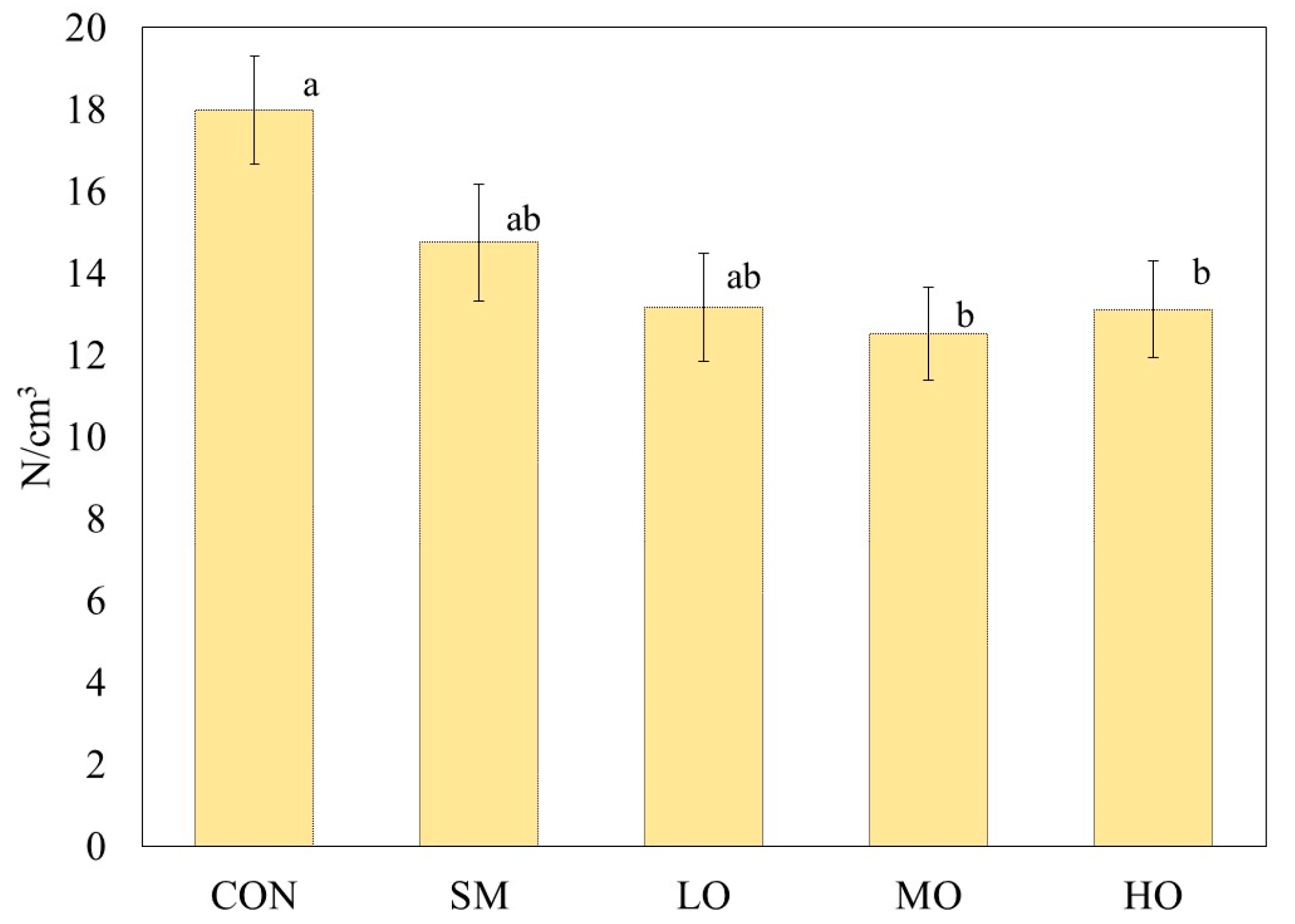
2.2. Compression Strength
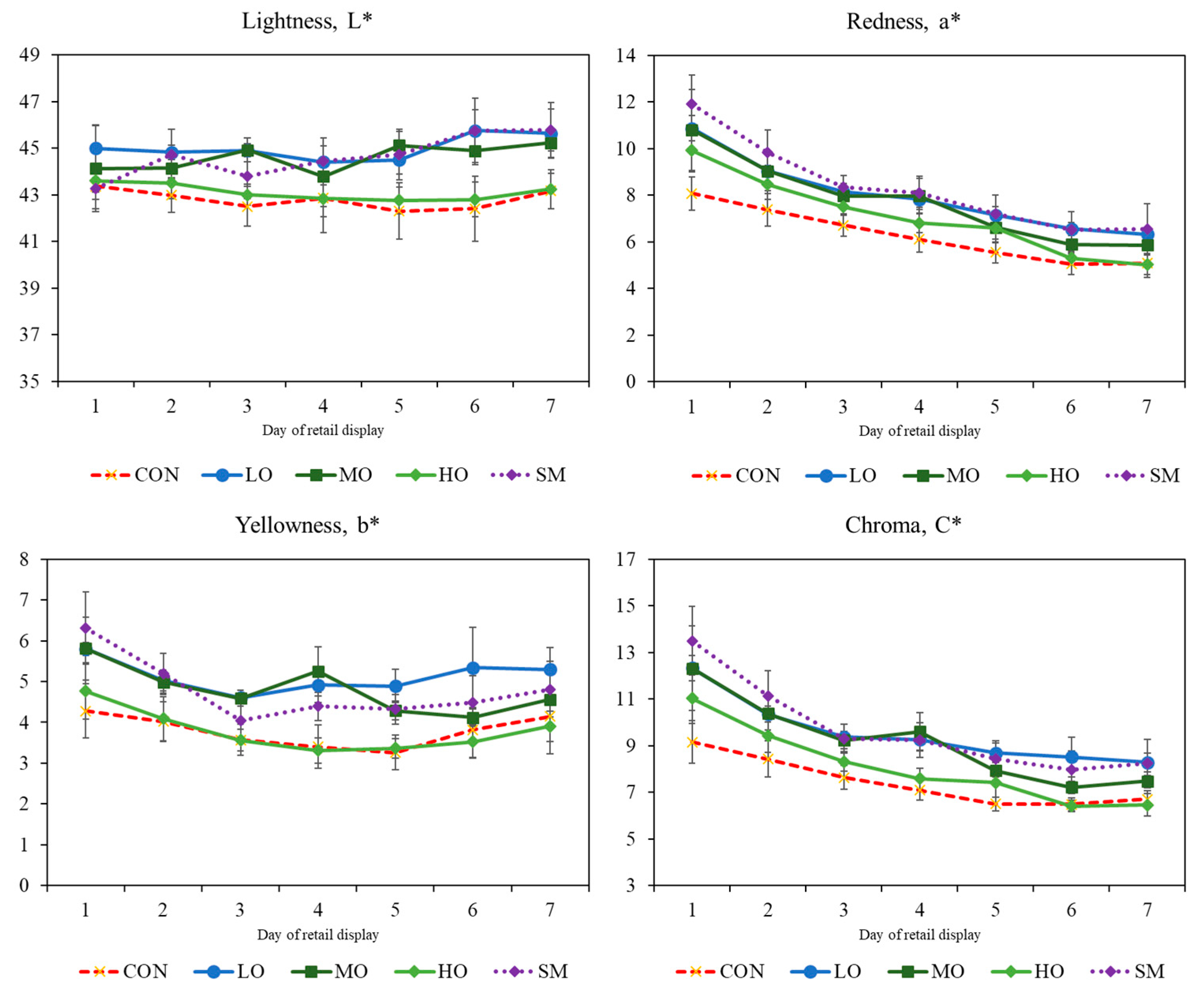
2.3. Colour
2.4. Fatty Acid Profile
3. Discussion
3.1. Lipid Oxidation
3.2. Compression Strength
3.3. Colour
3.4. Fatty Acid Profile
4. Materials and Methods
4.1. Animal Handling and Treatment Description
4.2. Lipid Oxidation
4.3. Compression Strenght Analysis
4.4. Colour
4.5. Fatty Acids (FA) Profile Analysis
4.6. Analysis of Data
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Resurreccion, A.V. Sensory aspects of consumer choices for meat and meat products. Meat Sci. 2004, 66, 11–20. [Google Scholar] [CrossRef]
- Kinnucan, H.W.; Xiao, H.; Hsia, C.-J.; Jackson, J.D. Effects of Health Information and Generic Advertising on U.S. Meat Demand. Am. J. Agric. Econ. 1997, 79, 13–23. [Google Scholar] [CrossRef]
- Schelling, G.T. Monensin mode of action in the rumen. J. Anim. Sci. 1984, 58, 1518–1527. [Google Scholar] [CrossRef]
- Millet, S.; Maertens, L. The European ban on antibiotic growth promoters in animal feed: From challenges to opportunities. Vet. J. 2011, 187, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.; Heritage, J. Antibiotic growth-promoters in food animals. In assessing quality and safety of animal feeds. In Assesing Quality and Safety of Animal Feeds; Jutzi, S., Ed.; Food and agriculture organization of the United Nations: Rome, Italy, 2004; pp. 129–153. [Google Scholar]
- Kumar, Y.; Yadav, D.N.; Ahmad, T.; Narsaiah, K. Recent Trends in the Use of Natural Antioxidants for Meat and Meat Products. Compr. Rev. Food Sci. Food Saf. 2015, 14, 796–812. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.A.; Bosco, S.J.D.; Mir, S.A. Plant extracts as natural antioxidants in meat and meat products. Meat Sci. 2014, 98, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, F.; Ji, B.P.; Pei, R.S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef]
- Aguirre, A.; Borneo, R.; León, A.E. Antimicrobial, mechanical and barrier properties of triticale protein films incorporated with oregano essential oil. Food Biosci. 2013, 1, 2–9. [Google Scholar] [CrossRef]
- Piñon, M.I.; Alarcon-Rojo, A.D.; Renteria, A.L.; Mendez, G.; Janacua-Vidales, H. Reduction of microorganisms in marinated poultry breast using oregano essential oil and power ultrasound. Acta Aliment. 2015, 44, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Gündüz, G.T.; Gönül, Ş.A.; Karapınar, M. Efficacy of oregano oil in the inactivation of Salmonella typhimurium on lettuce. Food Control 2010, 21, 513–517. [Google Scholar] [CrossRef]
- Valenzuela-Grijalva, N.V.; Pinelli-Saavedra, A.; Muhlia-Almazan, A.; Domínguez-Díaz, D.; González-Ríos, H. Dietary inclusion effects of phytochemicals as growth promoters in animal production. J. Anim. Sci. Technol. 2017, 59, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.E.; Maillard, J.Y.; Russel, A.D.; Catrenich, C.E.; Charbonneau, A.L.; Bartolo, R.G. Activity and mechanisms of action of selected biocidal agents on Gram-positive and -negative bacteria. J. Appl. Microbiol. 2003, 94, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, M.; Canbolat, O. Effect of carvacrol on intake, rumen fermentation, growth performance and carcass characteristics of growing lambs. J. Appl. Anim. Res. 2010, 38, 245–248. [Google Scholar] [CrossRef]
- Chikwanha, O.C.; Vahmani, P.; Muchenje, V.; Dugan, M.E.R.; Mapiye, C. Nutritional enhancement of sheep meat fatty acid profile for human health and wellbeing. Food Res. Int. 2018, 104, 25–38. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Larraín, R.E. Impacts of fat fromruminants’ meat on cardiovascular health and possible strategies to alter its lipid composition. J. Sci. Food Agric. 2017, 97, 1969–1978. [Google Scholar] [CrossRef]
- Xu, R. Important bioactive properties of omega-3 fatty acids. Ital. J. Food Sci. 2015, 27, 129–135. [Google Scholar]
- Chen, G.C.; Yang, J.; Eggersdorfer, M.; Zhang, W.; Qin, L.Q. N-3 long-chain polyunsaturated fatty acids and risk of all-cause mortality among general populations: A meta-analysis. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zárate, R.; el Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Le, H.; Nguyen, D.V.; Vu Nguyen, Q.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Fatty acid profiles of muscle, liver, heart and kidney of Australian prime lambs fed different polyunsaturated fatty acids enriched pellets in a feedlot system. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- World Health Organization Diet, Nutrition and the Prevention of Chronic Diseases; WHO Technical Report Series 916: Geneva, Switzerland, 2003.
- Miltko, R.; Majewska, M.P.; Bełżecki, G.; Kula, K.; Kowalik, B. Growth performance, carcass and meat quality of lambs supplemented different vegetable oils. Asian-Australas J. Anim. Sci. 2019, 32, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Camo, J.; Lorés, A.; Djenane, D.; Beltrán, J.A.; Roncalés, P. Display life of beef packaged with an antioxidant active film as a function of the concentration of oregano extract. Meat Sci. 2011, 88, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Chouliara, E.; Karatapanis, A.; Savvaidis, I.N.; Kontominas, M.G. Combined effect of oregano essential oil and modified atmosphere packaging on shelf-life extension of fresh chicken breast meat, stored at 4 °C. Food Microbiol. 2007, 24, 607–617. [Google Scholar] [CrossRef]
- Gallo, M.; Ferracane, R.; Naviglio, D. Antioxidant addition to prevent lipid and protein oxidation in chicken meat mixed with supercritical extracts of Echinacea angustifolia. J. Supercrit. Fluids 2012, 72, 198–204. [Google Scholar] [CrossRef]
- Karabagias, I.; Badeka, a.; Kontominas, M.G. Shelf life extension of lamb meat using thyme or oregano essential oils and modified atmosphere packaging. Meat Sci. 2011, 88, 109–116. [Google Scholar] [CrossRef]
- Petrou, S.; Tsiraki, M.; Giatrakou, V.; Savvaidis, I.N. Chitosan dipping or oregano oil treatments, singly or combined on modified atmosphere packaged chicken breast meat. Int. J. Food Microbiol. 2012, 156, 264–271. [Google Scholar] [CrossRef]
- Simitzis, P.E.; Symeon, G.K.; Charismiadou, M.A.; Bizelis, J.A.; Deligeorgis, S.G. The effects of dietary oregano oil supplementation on pig meat characteristics. Meat Sci. 2010, 84, 670–676. [Google Scholar] [CrossRef]
- Giannenas, I.; Tzora, A.; Bonos, E.; Sarakatsianos, I.; Karamoutsios, A.; Anastasiou, I.; Skoufos, I. Effects of dietary oregano essential oil, laurel essential oil and attapulgite on chemical composition, oxidative stability, fatty acid profile and mineral content of chicken breast and thigh meat. Eur. Poult. Sci. 2016, 80, 1–18. [Google Scholar]
- Cobellis, G.; Trabalza-Marinucci, M.; Yu, Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 2016, 545–546, 556–568. [Google Scholar] [CrossRef]
- Lourenço, M.; Cardozo, P.W.; Calsamiglia, S.; Fievez, V. Effects of saponins, quercetin, eugenol, and cinnamaldehyde on fatty acid biohydrogenation of forage polyunsaturated fatty acids in dual-flow continuous culture fermenters1. J. Anim. Sci. 2008, 86, 3045–3053. [Google Scholar] [CrossRef]
- Nieto, G.; Bañón, S.; Garrido, M.D. Effect of supplementing ewes’ diet with thyme (Thymus zygis ssp. gracilis) leaves on the lipid oxidation of cooked lamb meat. Food Chem. 2011, 125, 1147–1152. [Google Scholar]
- Rivaroli, D.C.; Guerrero, A.; Velandia Valero, M.; Zawadzki, F.; Eiras, C.E.; del Mar Campo, M.; Sañudo, C.; Mendes Jorge, A.; Nunes do Prado, I. Effect of essential oils on meat and fat qualities of crossbred young bulls finished in feedlots. Meat Sci. 2016, 121, 278–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galati, G.; O’Brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Nemeikaite-Čeniene, A.; Imbrasaite, A.; Sergediene, E.; Čenas, N. Quantitative structure-activity relationships in prooxidant cytotoxicity of polyphenols: Role of potential of phenoxyl radical/phenol redox couple. Arch. Biochem. Biophys. 2005, 441, 182–190. [Google Scholar] [CrossRef]
- Campo, M.M.; Nute, G.R.; Hughes, S.I.; Enser, M.; Wood, J.D.; Richardson, R.I. Flavour perception of oxidation in beef. Meat Sci. 2006, 72, 303–311. [Google Scholar] [CrossRef]
- Miller, J.K.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Aurousseau, B. Oxygen radicals in farm animals. Physiological effects and consequences on animal products. INRA Prod. Anim. 2002, 15, 67–82. [Google Scholar]
- Bhatt, N. Herbs and Herbal Supplements, a Novel Nutritional Approach in Animal Nutrition. Iran. J. Appl. Anim. Sci. 2015, 5, 497–516. [Google Scholar]
- Fasseas, M.K.; Mountzouris, K.C.; Tarantilis, P.A.; Polissiou, M.; Zervas, G. Antioxidant activity in meat treated with oregano and sage essential oils. Food Chem. 2008, 106, 1188–1194. [Google Scholar] [CrossRef]
- Rice-evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The Relative Antioxidant Activities of Plant-Derived Polyphenolic Flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Decker, E.A.; Park, Y. Healthier meat products as functional foods. Meat Sci. 2010, 86, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Govaris, A.; Botsoglou, N.; Papageorgiou, G.; Botsoglou, E.; Ambrosiadis, I. Dietary versus post-mortem use of oregano oil and/or α-tocopherol in turkeys to inhibit development of lipid oxidation in meat during refrigerated storage. Int. J. Food Sci. Nutr. 2004, 55, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Simitzis, P.E. Enrichment of Animal Diets with Essential Oils—A Great Perspective on Improving Animal Performance and Quality Characteristics of the Derived Products. Medicines 2017, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemipour, H.; Khaksar, V.; Rubio, L.A.; Veldkamp, T.; van Krimpen, M.M. Effect of feed supplementation with a thymol plus carvacrol mixture, in combination or not with an NSP-degrading enzyme, on productive and physiological parameters of broilers fed on wheat-based diets. Anim. Feed Sci. Technol. 2016, 211, 117–131. [Google Scholar] [CrossRef]
- Chaves, A.V.; Stanford, K.; Gibson, L.L.; McAllister, T.A.; Benchaar, C. Effects of carvacrol and cinnamaldehyde on intake, rumen fermentation, growth performance, and carcass characteristics of growing lambs. Anim. Feed Sci. Technol. 2008, 145, 396–408. [Google Scholar] [CrossRef]
- Cardozo, P.W.; Calsamiglia, S.; Ferret, A.; Kamel, C. Screening for the effects of natural plant extracts at different pH on in vitro rumen microbial fermentation of a high-concentrate diet for beef cattle1. J. Anim. Sci. 2005, 83, 2572–2579. [Google Scholar] [CrossRef]
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.R.; McAllister, T.A.; Beauchemin, K.A. A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed Sci. Technol. 2008, 145, 209–228. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Grigoropoulou, S.H.; Botsoglou, E.; Govaris, A.; Papageorgiou, G. The effects of dietary oregano essential oil and α-tocopheryl acetate on lipid oxidation in raw and cooked Turkey during refrigerated storage. Meat Sci. 2003, 65, 1193–1200. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Christaki, E.; Fletouris, D.J.; Florou-Paneri, P.; Spais, A.B. The effect of dietary oregano essential oil on lipid oxidation in raw and cooked chicken during refrigerated storage. Meat Sci. 2002, 62, 259–265. [Google Scholar] [CrossRef]
- Fernandes, R.P.P.; Trindade, M.A.; Tonin, F.G.; Pugine, S.M.P.; Lima, C.G.; Lorenzo, J.M.; de Melo, M.P. Evaluation of oxidative stability of lamb burger with Origanum vulgare extract. Food Chem. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Watkins, P.J.; Frank, D.; Singh, T.K.; Young, O.A.; Warner, R.D. Sheepmeat Flavor and the Effect of Different Feeding Systems: A Review. J. Agric. Food Chem. 2013, 61, 3561–3579. [Google Scholar] [CrossRef]
- Wheeler, T.L.; Shackelford, S.D.; Koohmaraie, M. Variation in proteolysis, sarcomere length, collagen content, and tenderness among major pork muscles. J. Anim. Sci. 2000, 78, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Castillejos, L.; Calsamiglia, S.; Ferret, A. Effect of essential oil active compounds on rumen microbial fermentation and nutrient flow in in vitro systems. J. Dairy Sci. 2006, 89, 2649–2658. [Google Scholar] [CrossRef]
- Smith, G.C.; Dutson, T.R.; Hostetler, R.L.; Carpenter, Z.L. Fatness, rate of chilling and tenderness of lamb. J. Food Sci. 1976, 41, 748–756. [Google Scholar] [CrossRef]
- Simitzis, P.E.; Deligeorgis, S.G.; Bizelis, J.A.; Dardamani, A.; Theodosiou, I.; Fegeros, K. Effect of dietary oregano oil supplementation on lamb meat characteristics. Meat Sci. 2008, 79, 217–223. [Google Scholar] [CrossRef]
- Demirel, G.; Ekiz, B.; Unal, A.; Yalcintan, H.; Kocabagli, N.; Yilmaz, A. Effects of dietary oregano essential oil on carcass and meat quality of kivircik lambs. J. Anim. Vet. Adv. 2013, 12, 991–995. [Google Scholar]
- Cheng, C.; Liu, Z.; Zhou, Y.; Wei, H.; Zhang, X.; Xia, M.; Deng, Z.; Zou, Y.; Jiang, S.; Peng, J. Effect of oregano essential oil supplementation to a reduced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of Longissimus thoracis muscle in growing-finishing pigs. Meat Sci. 2017, 133, 103–109. [Google Scholar] [CrossRef]
- Forte, C.; Ranucci, D.; Beghelli, D.; Branciari, R.; Acuti, G.; Todini, L.; Cavallucci, C.; Trabalza-Marinucci, M. Dietary integration with oregano (Origanum vulgare L.) essential oil improves growth rate and oxidative status in outdoor-reared, but not indoor-reared, pigs. J. Anim. Physiol. Anim. Nutr. 2017, 101, e352–e361. [Google Scholar] [CrossRef]
- Ranucci, D.; Beghelli, D.; Trabalza-Marinucci, M.; Branciari, R.; Forte, C.; Olivieri, O.; Badillo Pazmay, G.V.; Cavallucci, C.; Acuti, G. Dietary effects of a mix derived from oregano (Origanum vulgare L.) essential oil and sweet chestnut (Castanea sativa Mill.) wood extract on pig performance, oxidative status and pork quality traits. Meat Sci. 2015, 100, 319–326. [Google Scholar] [CrossRef]
- Rossi, R.; Pastorelli, G.; Cannata, S.; Tavaniello, S.; Maiorano, G.; Corino, C. Effect of long term dietary supplementation with plant extract on carcass characteristics meat quality and oxidative stability in pork. Meat Sci. 2013, 95, 542–548. [Google Scholar] [CrossRef]
- Nieto, G.; Jongberg, S.; Andersen, M.L.; Skibsted, L.H. Thiol oxidation and protein cross-link formation during chill storage of pork patties added essential oil of oregano, rosemary, or garlic. Meat Sci. 2013, 95, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Zhang, K.; Li, C.; Wu, J.; Davis, D.; Casper, D.; Jiang, H.; Jiao, T.; Wang, X.; Wang, J. Dietary supplementation with Essential-oils-cobalt for improving growth performance, meat quality and skin cell capacity of goats. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Velasco, V.; Orellana, C.; Williams, P.; Campos, J.; Melín, P. Meat Quality Characteristics of Goats Fed with Dietary Dry Oregano (Origanum vulgare L.). Chil. J. Agric. Anim. Sci. 2011, 27, 95–104. [Google Scholar]
- Payne, C.E.; Pannier, L.; Anderson, F.; Pethick, D.W.; Gardner, G.E. Lamb age has little impact on eating quality. Foods 2020, 9, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbut, S. Principles of meat processing. In The Science of Poultry and Meat Processing; Barbut, S., Ed.; University of Guelp: Guelp, ON, Canada, 2015. [Google Scholar]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.; O’Grady, M.N.; O’Callaghan, Y.C.; O’Brien, N.M.; Kerry, J.P. Evaluation of the antioxidant potential of grape seed and bearberry extracts in raw and cooked pork. Meat Sci. 2007, 76, 604–610. [Google Scholar] [CrossRef]
- Teixeira, A.; Batista, S.; Delfa, R.; Cadavez, V. Lamb meat quality of two breeds with protected origin designation. Influence of breed, sex and live weight. Meat Sci. 2005, 71, 530–536. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Fogarty, N.M. Diverse lamb genotypes -2. Meat pH, colour and tenderness. Meat Sci. 1998, 49, 477–488. [Google Scholar] [CrossRef]
- Ramos, F.A.; Martínez, A.P.; Montes, E.S.; Gaytán, C.N.; Selene, A.; Cázarez, H.; Tovar, J.C.; Sánchez, J.G.; Rodríguez, C. Oregano oil use in broiler diet increases accumulation of carvacrol and thymol in breast meat Broiler production. Acta Univ. 2017, 27, 34–39. [Google Scholar]
- Boroski, M.; Giroux, H.J.; Sabik, H.; Petit, H.V.; Visentainer, J.V.; Matumoto-Pintro, P.T.; Britten, M. Use of oregano extract and oregano essential oil as antioxidants in functional dairy beverage formulations. Food Sci. Technol. 2012, 47, 167–174. [Google Scholar] [CrossRef]
- Hernández-Hernández, E.; Ponce-Alquicira, E.; Jaramillo-Flores, M.E.; Guerrero Legarreta, I. Antioxidant effect rosemary (Rosmarinus officinalis L.) and oregano (Origanum vulgare L.) extracts on TBARS and colour of model raw pork batters. Meat Sci. 2009, 81, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Effect of thymol and carvacrol feed supplementation on performance, antioxidant enzyme activities, fatty acid composition, digestive enzyme activities, and immune response in broiler chickens. Poult. Sci. 2013, 8, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Petron, M.J.; Raes, K.; Claeys, E.; Lourenco, M.; Fremaut, D.; De Smet, S. Effect of grazing pastures of different botanical composition on antioxidant enzyme activities and oxidative stability of lamb meat. Meat Sci. 2007, 75, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Moura, L.V.; Oliveira, E.R.; Fernandes, A.R.M.; Gabriel, A.M.A.; Silva, L.H.X.; Takiya, C.S.; Cônsolo, N.R.B.; Rodrigues, G.C.G.; Lemos, T.; Gandra, J.R. Feed efficiency and carcass traits of feedlot lambs supplemented either monensin or increasing doses of copaiba (Copaifera spp.) essential oil. Anim. Feed Sci. Technol. 2017, 232, 110–118. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Invited Review: Essential Oils as Modifiers of Rumen Microbial Fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef] [Green Version]
- Ogunade, I.; Schweickart, H.; Andries, K.; Lay, J.; Adeyemi, J. Monensin Alters the Functional and Metabolomic Profile of Rumen Microbiota in Beef Cattle. Animals 2018, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, S.C.; Young, D.G.; Oberg, C.J. Screening for Inhibitory Activity of Essential Oils on Selected Bacteria, Fungi and Viruses. J. Essent. Oil Res. 2000, 12, 639–649. [Google Scholar] [CrossRef]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. Determining the antimicrobial actions of tea tree oil. Molecules 2001, 6, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Castro-Montoya, J.M.; Makkar, H.P.S.; Becker, K. Effects of monensin on the chemical composition of the liquid associated microbial fraction in an in vitro rumen fermentation system. Livest. Sci. 2012, 150, 414–418. [Google Scholar] [CrossRef]
- Boucqué, C.V.; Fiems, L.O.; Cottyn, B.G.; Casteels, M.; Buysse, F.X. Monensin-sodium as a performance-promoting additive for fattening bulls and its impact on carcass and meat quality characteristics. Anim. Feed Sci. Technol. 1982, 7, 401–410. [Google Scholar] [CrossRef]
- Smith, S.B.; Kawachi, H.; Choi, C.B.; Choi, C.W.; Wu, G.; Sawyer, J.E. Cellular regulation of bovine intramuscular adipose tissue development and composition. J. Anim. Sci. 2009, 87, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.B.; Strobel, H.J. Effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Isabel Osorio-Teran, A.; David Mendoza-Martínez, G.; Alberto Miranda-Romero, L.; Martínez-Gomez, D.; Abel Hernández-García, P.; Antonio Martínez-García, J.; Brasileira de Zootecnia, R. Effect of calcium propionate and monensin on in vitro digestibility and gas production. Brazilian J. Anim. Sci. 2017, 46, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Wood, J.D.; Nute, G.R.; Richardson, R.I.; Whittington, F.M.; Southwood, O.; Plastow, G.; Mansbridge, R.; Da Costa, N.; Chang, K.C. Effects of breed, diet and muscle on fat deposition and eating quality in pigs. Meat Sci. 2004, 67, 651–667. [Google Scholar] [CrossRef]
- Ladeira, M.M.; Santarosa, L.C.; Chizzotti, M.L.; Ramos, E.M.; Machado Neto, O.R.; Oliveira, D.M.; Carvalho, J.R.R.; Lopes, L.S.; Ribeiro, J.S. Fatty acid profile, color and lipid oxidation of meat from young bulls fed ground soybean or rumen protected fat with or without monensin. Meat Sci. 2014, 96, 597–605. [Google Scholar] [CrossRef]
- Daley, C.; Abbott, A.; Doyle, P.; Nader, G.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.C.; Santos, G.T.; Branco, A.F.; Damasceno, J.C.; Kazama, R.; Matsushita, M.; Horst, J.A.; Santos, W.B.R.; Petit, H. V Production performance and milk composition of dairy cows fed whole or ground flaxseed with or without monensin. J. Dairy Sci. 2007, 90, 2928–2936. [Google Scholar] [CrossRef]
- Parvar, R.; Ghoorchi, T.; Kashfi, H.; Parvar, K. Effect of Ferulago angulata (Chavil) essential oil supplementation on lamb growth performance and meat quality characteristics. Small Rumin. Res. 2018, 167, 48–54. [Google Scholar] [CrossRef]
- Dávila-Ramírez, J.L.; Avendaño-Reyes, L.; MacÍas-Cruz, U.; Peña-Ramos, E.A.; Islava-Lagarda, T.Y.; Zamorano-García, L.; Valenzuela-Melendres, M.; Camou, J.P.; González-Ríos, H. Fatty acid composition and physicochemical and sensory characteristics of meat from ewe lambs supplemented with zilpaterol hydrochloride and soybean oil. Anim. Prod. Sci. 2017, 57, 767–777. [Google Scholar] [CrossRef]
- Committee on the Nutrient Requirements of Small Ruminants, National Research Council. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Tarladgis, B.; Watts, B.; Younathan, M.; Dugan, L. A distillation method for the quantitative determination of malonaldehyde in rancid foods. J. Am. Oil Chem. Soc. 1960, 37, 44–48. [Google Scholar] [CrossRef]
- American Meat Science Association. Research Guidelines for Cookery, Sensory Evaluation, and Instrumental Tenderness Measurements of Meat; AMSC: Champaign, IL, USA, 2016. [Google Scholar]
- Segars, R.A.; Hamel, R.G.; Kapsalis, J.G.; Klutter, R.A. A punch and die test cell for determining the textural qualities of meat. J. Texture Stud. 1975, 6, 211–225. [Google Scholar] [CrossRef]
- Jones, R.C.; Dransfield, E.; Robinson, J.M.; Crosland, A.R. Correlation of mechanical properties, composition and perceived texture of beefburgers. J. Texture Stud. 1985, 16, 241–262. [Google Scholar] [CrossRef]
- Roufs, J.A.J. Light as a True Visual Quantity: Principles of Measurement; Commission Internationale de l’Éclairage: Paris, France, 1978. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.K. Improved Bligh and Dyer extraction procedure. Lipid Technol. 2008, 20, 280–281. [Google Scholar] [CrossRef]
- AOAC Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1995.
- R Team Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Treatments | |||||
|---|---|---|---|---|---|---|
| CON | SM | HO | MO | LO | p-Value | |
| C14:0 | 1.95 | 1.68 | 1.99 | 2.00 | 1.78 | 0.60 |
| C14:1 | 0.11 | 0.10 | 0.09 | 0.10 | 0.10 | 0.99 |
| C16:0 | 25.00 | 23.69 | 23.26 | 25.64 | 25.07 | 0.49 |
| C16:1n7 | 1.00ab | 1.35a | 0.87b | 0.91ab | 1.06ab | 0.04 |
| C18:0 | 14.69 | 15.07 | 15.04 | 15.87 | 14.90 | 0.94 |
| C18:1n9t | 2.31 | 2.22 | 1.63 | 2.84 | 2.20 | 0.19 |
| C18:1n9c | 44.17 | 46.28 | 45.83 | 44.89 | 46.24 | 0.52 |
| C18:2n6t | 0.25 | 0.24 | 0.24 | 0.22 | 0.21 | 0.97 |
| C18:2n6c | 3.10 | 3.32 | 3.28 | 2.69 | 2.85 | 0.60 |
| C18:3n6 | 0.09 | 0.06 | 0.07 | 0.05 | 0.03 | 0.78 |
| C18:3n3 | 0.06 | 0.04 | 0.04 | 0.04 | 0.04 | 0.17 |
| C20:0 | 0.38 | 0.27 | 0.18 | 0.25 | 0.32 | 0.10 |
| C20:1n9 | 0.72ab | 0.62b | 0.69ab | 0.82a | 0.58b | 0.01 |
| C20:3n3 | 0.05 | 0.05 | 0.08 | 0.04 | 0.04 | 0.35 |
| C20:4n6 | 0.14 | 0.13 | 0.08 | 0.05 | 0.08 | 0.46 |
| C20:5n3 | 1.68 | 1.48 | 1.81 | 1.25 | 1.12 | 0.13 |
| C22:5n3 | 0.86 | 0.39 | 0.53 | 0.33 | 0.46 | 0.27 |
| C22:6n3 | 0.30ab | 0.23ab | 0.86a | 0.21b | 0.23ab | 0.03 |
| Fatty Acid | Treatments | |||||
|---|---|---|---|---|---|---|
| CON | SM | HO | MO | LO | p-Value | |
| Saturated | ||||||
| C14:0 | 13.99b | 14.47ab | 9.67b | 23.50a | 13.18b | 0.042 |
| C16:0 | 172.7b | 200.1ab | 120.88b | 289.77a | 183.74ab | 0.031 |
| C18:0 | 94.93b | 121.7ab | 71.94b | 164.17a | 103.58ab | 0.043 |
| Mono-unsaturated | ||||||
| C16:1n7 | 6.60ab | 10.68a | 4.31b | 9.51a | 7.14ab | 0.024 |
| C18:1n9c | 287.41b | 369.6ab | 221.58b | 472.97a | 321.59ab | 0.043 |
| C18:1n9t | 14.67 | 16.82 | 14.39 | 8.12 | 30.19 | 0.007 |
| C20:1n9 | 2.90b | 3.54b | 4.21b | 7.88a | 4.46b | 0.001 |
| PUFA’s | ||||||
| C18:2n6t | 18.74ab | 24.88ab | 14.04b | 27.10a | 18.66ab | 0.043 |
| Treatment | |||||
|---|---|---|---|---|---|
| Ingredients (%) | Control | Monensin | Low Oil | Medium Oil | High Oil |
| Rolled sorghum | 36.32 | 36.32 | 36.32 | 36.32 | 36.32 |
| Soybean meal | 34.79 | 34.79 | 34.79 | 34.79 | 34.79 |
| Alfalfa hay, full bloom | 20 | 20 | 20 | 20 | 20 |
| Molasses cane | 5 | 5 | 5 | 5 | 5 |
| Corn gluten (60% CP) | 2 | 2 | 2 | 2 | 2 |
| Calcium carbonate | 0.883 | 0.883 | 0.883 | 0.883 | 0.883 |
| Salt | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Mineral premix 1 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Essential oil (Carvacrol, g/Kg MS) | 0 | 0 | 0.2 | 0.3 | 0.4 |
| Monensin (pmm) | 0 | 33 | 0 | 0 | 0 |
| Calculated chemical composition (% DM basis) | |||||
| CP | 23.68 | 23.68 | 23.68 | 23.68 | 23.68 |
| ME (Mcal/kg) | 2.739 | 2.739 | 2.739 | 2.739 | 2.739 |
| Ca | 0.791 | 0.791 | 0.791 | 0.791 | 0.791 |
| P | 0.453 | 0.453 | 0.453 | 0.453 | 0.453 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Galicia, I.A.; Arras-Acosta, J.A.; Huerta-Jimenez, M.; Rentería-Monterrubio, A.L.; Loya-Olguin, J.L.; Carrillo-Lopez, L.M.; Tirado-Gallegos, J.M.; Alarcon-Rojo, A.D. Natural Oregano Essential Oil May Replace Antibiotics in Lamb Diets: Effects on Meat Quality. Antibiotics 2020, 9, 248. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9050248
Garcia-Galicia IA, Arras-Acosta JA, Huerta-Jimenez M, Rentería-Monterrubio AL, Loya-Olguin JL, Carrillo-Lopez LM, Tirado-Gallegos JM, Alarcon-Rojo AD. Natural Oregano Essential Oil May Replace Antibiotics in Lamb Diets: Effects on Meat Quality. Antibiotics. 2020; 9(5):248. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9050248
Chicago/Turabian StyleGarcia-Galicia, Ivan A., Jose A. Arras-Acosta, Mariana Huerta-Jimenez, Ana L. Rentería-Monterrubio, Jose L. Loya-Olguin, Luis M. Carrillo-Lopez, Juan M. Tirado-Gallegos, and Alma D. Alarcon-Rojo. 2020. "Natural Oregano Essential Oil May Replace Antibiotics in Lamb Diets: Effects on Meat Quality" Antibiotics 9, no. 5: 248. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9050248