Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes

 , and
, and 
Abstract
:
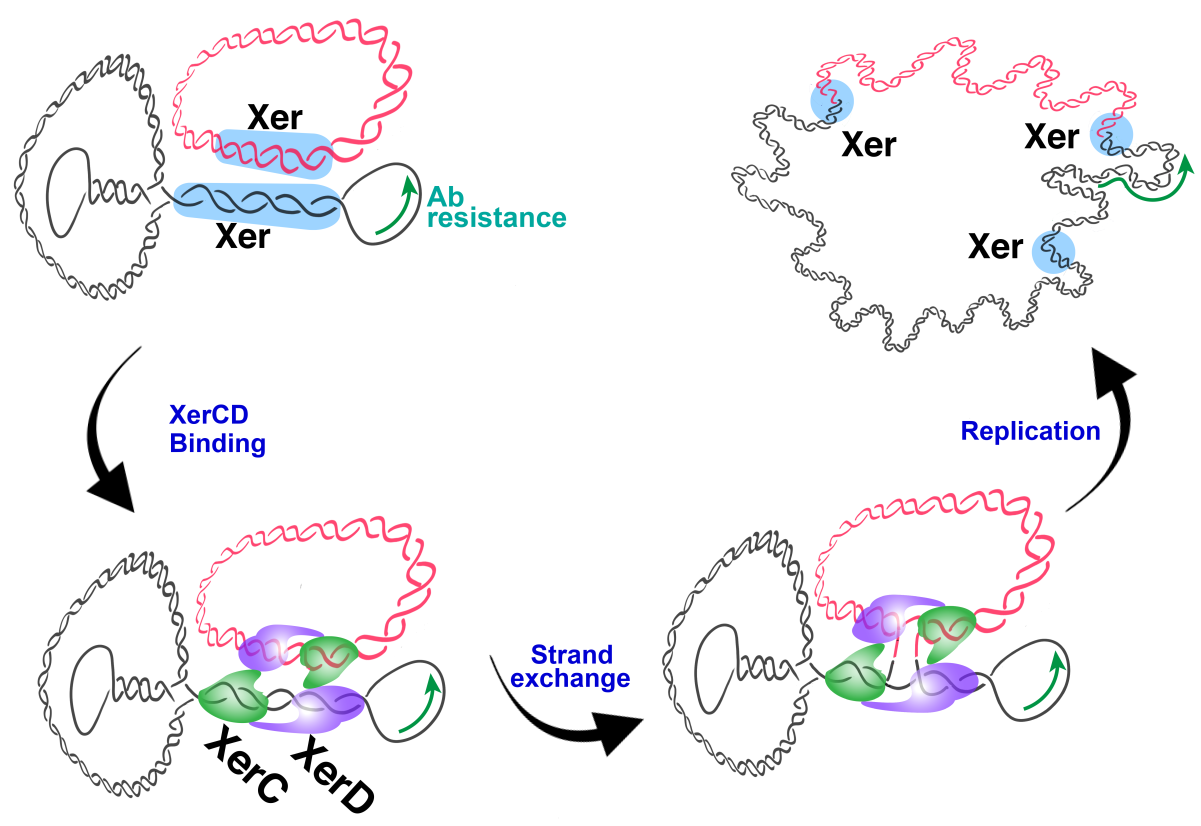
1. Introduction
2. Results
2.1. Cloning and Complementation Analysis of A. baumannii Xer Recombinases
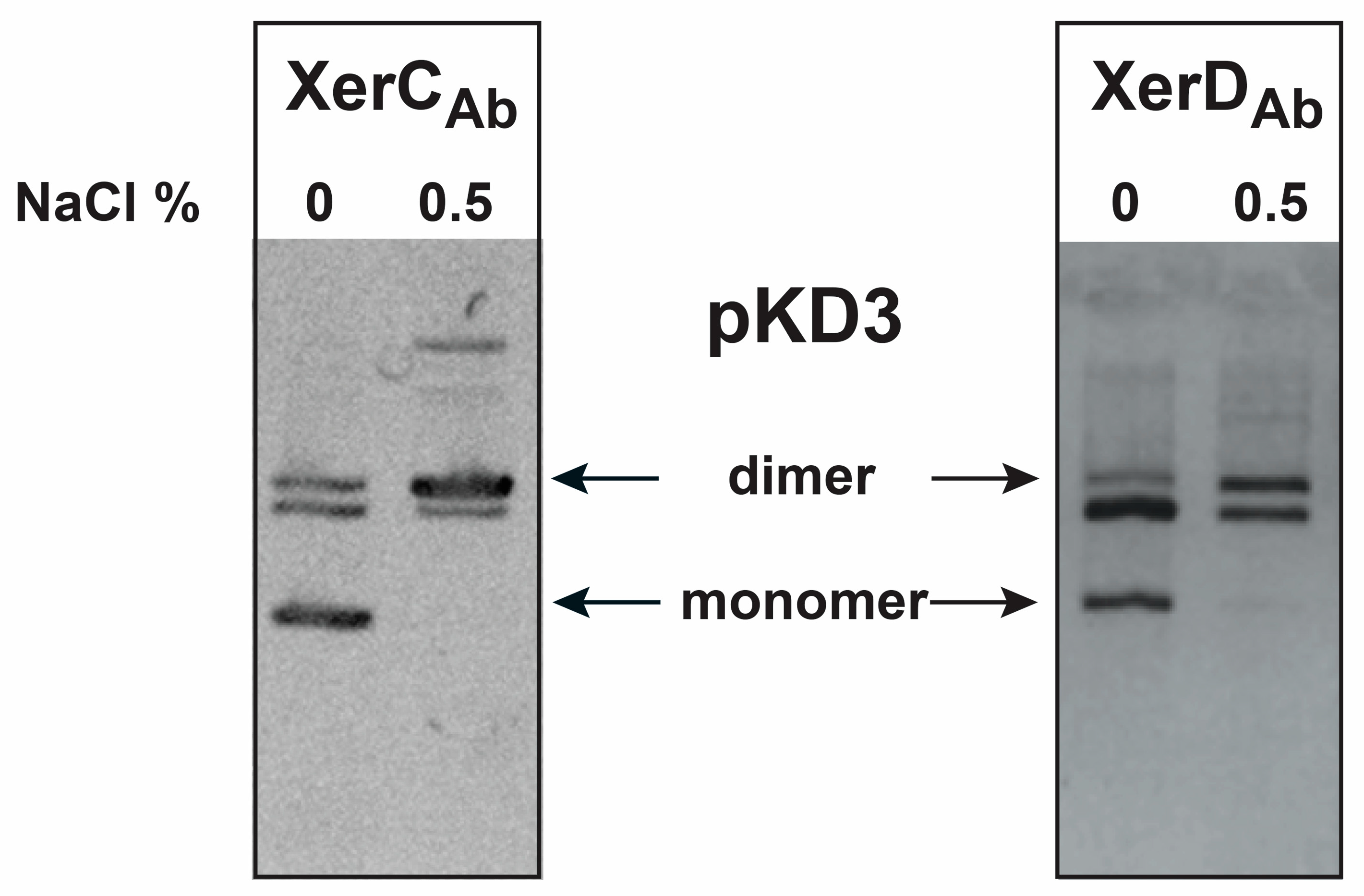
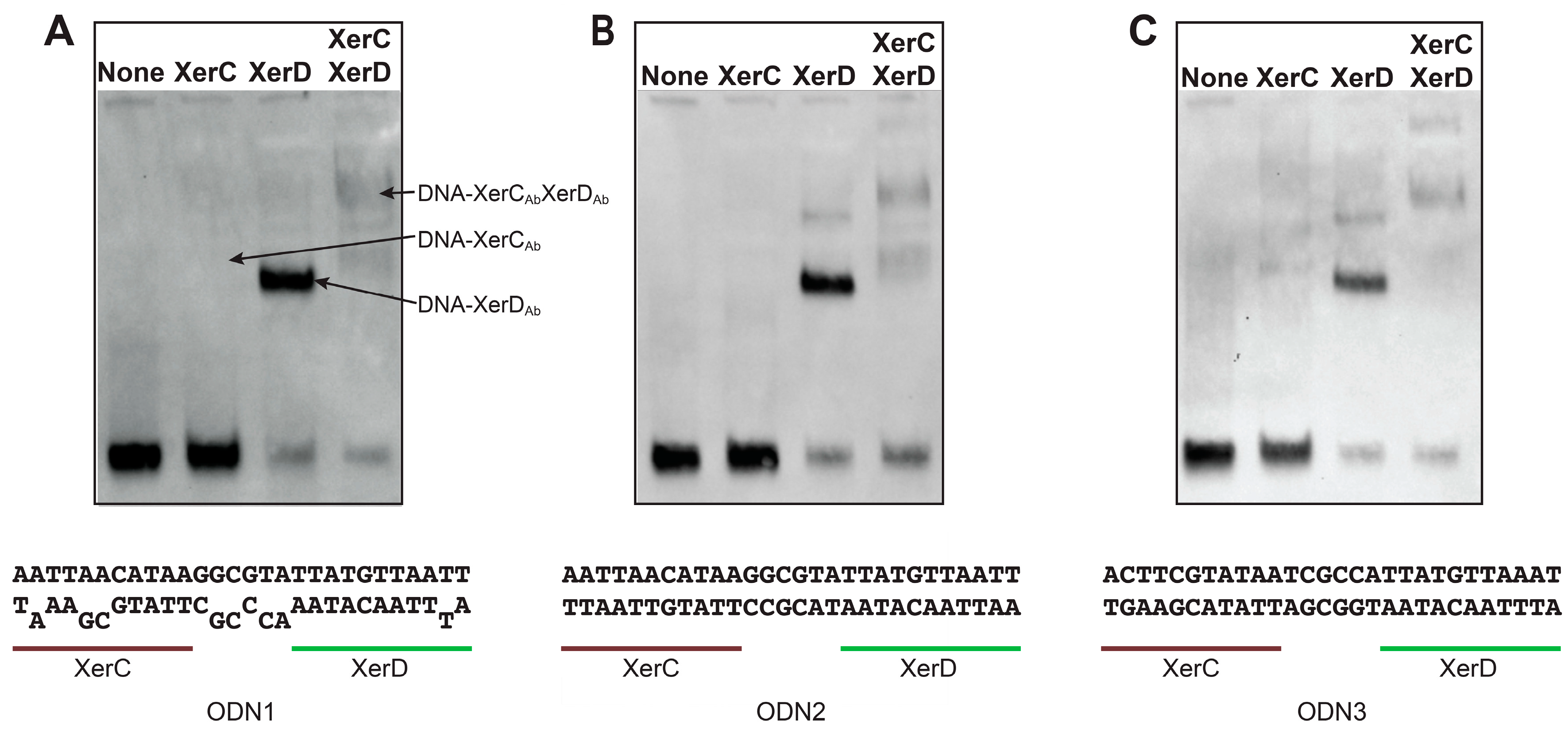
2.2. Binding of A. baumannii Xer Recombinases to XerC/D Binding Sites
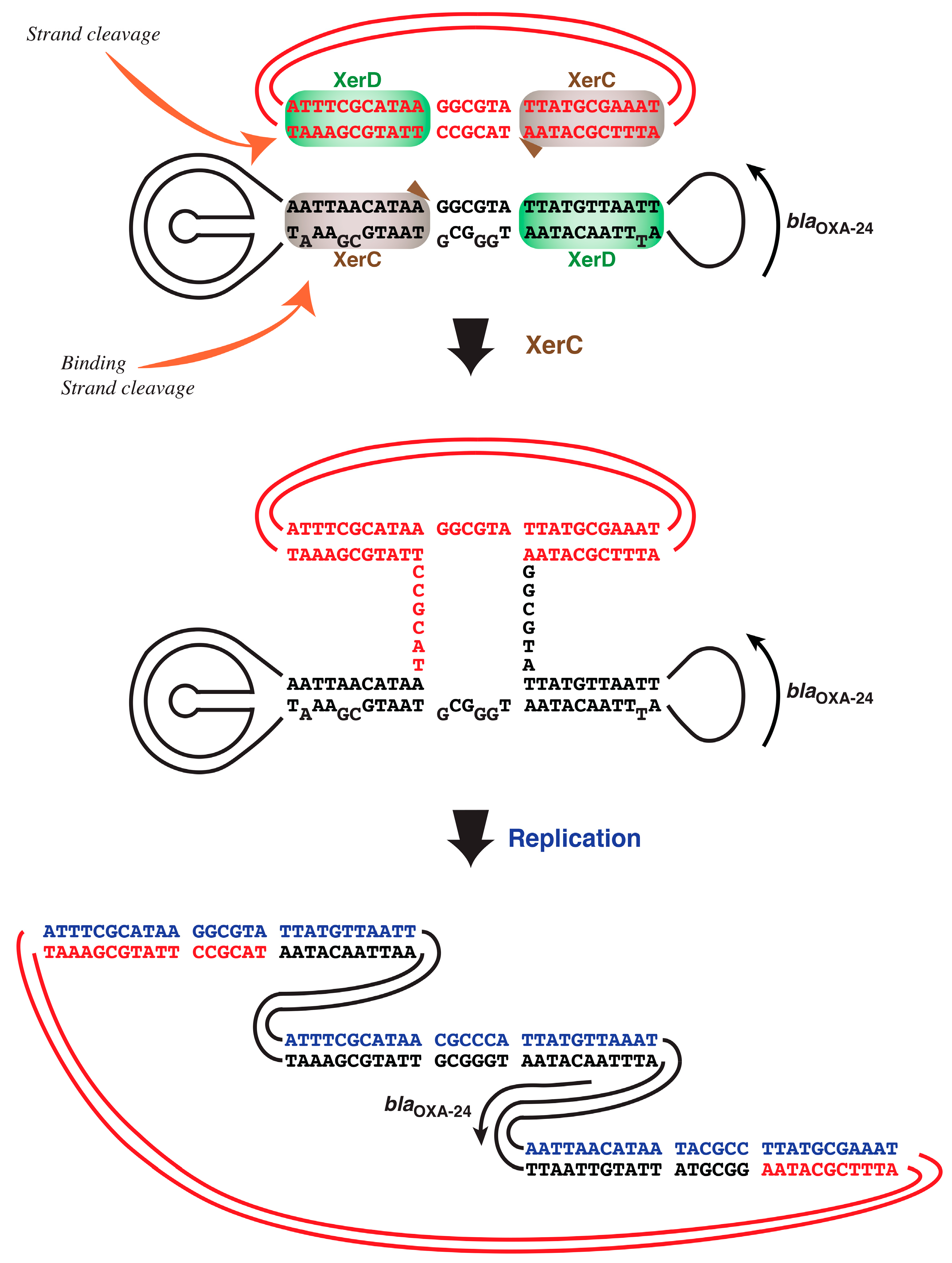
2.3. A. baumannii Xer Recombinases-Mediated Strand Exchange
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. General DNA Procedures
4.3. Protein Purification
4.4. In Vivo Resolution Assay
4.5. DNA-Binding Assay
4.6. In Vitro Xer-Mediated DNA Cleavage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain or Plasmid | Relevant Characteristics, Genotype, or Phenotype a | Source or Reference |
|---|---|---|
| E. coli strains | ||
| DS941 | AB1157 recF143 lacIq lacZΔM15 | [65] |
| DS981 | DS941 xerC (Kanr) | [56] |
| DS9028 | DS941 xerD (Tmpr) | [57] |
| DS981XerCAb | DS981 (pMSR1) (Kanr Tetr) | This work |
| DS9028XerDAb | DS9028 (pMSR2) (Tmpr Tetr) | This work |
| DS9040 | DS941 xerC xerD (Kanr Genr) | [34] |
| JC8679 | DS945 recBC sbcA (hyperrecombinogenic) | [39] |
| A. baumannii strain | ||
| A118 | Human clinical isolate | [58] |
| Plasmids | ||
| pMSR1 | xerCAb cloned into the pACYC184 EcoRI site (Tetr) | This work |
| pMSR2 | xerDAb cloned into the pACYC184 EcoRI site (Tetr) | This work |
| pBAD102xerCAb | xerCAb cloned into pBAD102 (Ampr) | This work |
| pBAD102xerDAb | xerDAb cloned into pBAD102 (Ampr) | This work |
| pKD3 | EcoRI-SacI fragment containing the pJHCMW1 mwr site with substitution C to T at the ArgR binding site cloned in pUC18 (Ampr) | [8] |
| pUC18 | Cloning vector (Ampr) | [60] |
| pCR2.1 | Cloning vector (Ampr, Kanr) | ThermoFisher |
| pACYC184 | Cloning vector, p15A replicon (Chlr Tetr) | [61] |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aussel, L.; Barre, F.X.; Aroyo, M.; Stasiak, A.; Stasiak, A.Z.; Sherratt, D. FtsK Is a DNA motor protein that activates chromosome dimer resolution by switching the catalytic state of the XerC and XerD recombinases. Cell 2002, 108, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Sherratt, D.J.; Soballe, B.; Barre, F.X.; Filipe, S.; Lau, I.; Massey, T.; Yates, J. Recombination and chromosome segregation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 61–69. [Google Scholar] [CrossRef]
- Zawadzki, P.; May, P.F.; Baker, R.A.; Pinkney, J.N.; Kapanidis, A.N.; Sherratt, D.J.; Arciszewska, L.K. Conformational transitions during FtsK translocase activation of individual XerCD-dif recombination complexes. Proc. Natl. Acad. Sci. USA 2013, 110, 17302–17307. [Google Scholar] [CrossRef] [Green Version]
- Summers, D.K.; Beton, C.W.; Withers, H.L. Multicopy plasmid instability: The dimer catastrophe hypothesis. Mol. Microbiol. 1993, 8, 1031–1038. [Google Scholar] [CrossRef]
- Colloms, S.D.; Alen, C.; Sherratt, D.J. The ArcA/ArcB two-component regulatory system of Escherichia coli is essential for Xer site-specific recombination at psi. Mol. Microbiol. 1998, 28, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Colloms, S.D. The topology of plasmid-monomerizing Xer site-specific recombination. Biochem. Soc. Trans. 2013, 41, 589–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, D.; Ramiscal, J.; Trigueros, S.; Newmark, J.S.; Do, A.; Sherratt, D.J.; Tolmasky, M.E. Differences in resolution of mwr-containing plasmid dimers mediated by the Klebsiella pneumoniae and Escherichia coli XerC recombinases: Potential implications in dissemination of antibiotic resistance genes. J. Bacteriol. 2006, 188, 2812–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, H.; Dery, K.J.; Sherratt, D.J.; Tolmasky, M.E. Osmoregulation of dimer resolution at the plasmid pJHCMW1 mwr locus by Escherichia coli XerCD recombination. J. Bacteriol. 2002, 184, 1607–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolmasky, M.E.; Colloms, S.; Blakely, G.; Sherratt, D.J. Stability by multimer resolution of pJHCMW1 is due to the Tn1331 resolvase and not to the Escherichia coli Xer system. Microbiology 2000, 146, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trigueros, S.; Tran, T.; Sorto, N.; Newmark, J.; Colloms, S.D.; Sherratt, D.J.; Tolmasky, M.E. mwr Xer site-specific recombination is hypersensitive to DNA supercoiling. Nucleic Acids Res. 2009, 37, 3580–3587. [Google Scholar] [CrossRef]
- Das, B.; Martinez, E.; Midonet, C.; Barre, F.X. Integrative mobile elements exploiting Xer recombination. Trends Microbiol. 2013, 21, 23–30. [Google Scholar] [CrossRef]
- Midonet, C.; Barre, F.X. Xer site-specific recombination: Promoting vertical and horizontal transmission of genetic information. Microbiol Spectr 2014, 2. MDNA3-0056-2014. [Google Scholar]
- Val, M.E.; Bouvier, M.; Campos, J.; Sherratt, D.; Cornet, F.; Mazel, D.; Barre, F.X. The single-stranded genome of phage CTX is the form used for integration into the genome of Vibrio cholerae. Mol. Cell 2005, 19, 559–566. [Google Scholar] [CrossRef]
- Campos, J.; Martinez, E.; Suzarte, E.; Rodriguez, B.L.; Marrero, K.; Silva, Y.; Ledon, T.; del Sol, R.; Fando, R. VGJ phi, a novel filamentous phage of Vibrio cholerae, integrates into the same chromosomal site as CTX phi. J. Bacteriol. 2003, 185, 5685–5696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, F.; Kamruzzaman, M.; Mekalanos, J.J.; Faruque, S.M. Satellite phage TLCphi enables toxigenic conversion by CTX phage through dif site alteration. Nature 2010, 467, 982–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharova, M.V.; Beletskaya, I.V.; Denjmukhametov, M.M.; Yurkova, T.V.; Semenova, L.M.; Shlyapnikov, M.G.; Solonin, A.S. Characterization of pECL18 and pKPN2: A proposed pathway for the evolution of two plasmids that carry identical genes for a Type II restriction-modification system. Mol. Genet. Genomics 2002, 267, 171–178. [Google Scholar] [CrossRef]
- Tran, T.; Andres, P.; Petroni, A.; Soler-Bistue, A.; Albornoz, E.; Zorreguieta, A.; Reyes-Lamothe, R.; Sherratt, D.J.; Corso, A.; Tolmasky, M.E. Small plasmids harboring qnrB19: A model for plasmid evolution mediated by site-specific recombination at oriT and Xer sites. Antimicrob. Agents Chemother. 2012, 56, 1821–1827. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Traglia, G.M.; Lin, D.L.; Tran, T.; Tolmasky, M.E. Plasmid-mediated antibiotic resistance and virulence in gram-negatives: The Klebsiella pneumoniae paradigm. Microbiol. Spectr. 2014, 2. PLAS-0016-2013. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, G.A.; Hall, R.M. The tet39 determinant and the msrE-mphE genes in Acinetobacter plasmids are each part of discrete modules flanked by inversely oriented pdif (XerC-XerD) sites. Antimicrob. Agents Chemother. 2017, 61, e00717–e00780. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Nordmann, P. Genetic structures at the origin of acquisition and expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-58 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2006, 50, 1442–1448. [Google Scholar] [CrossRef] [Green Version]
- Merino, M.; Acosta, J.; Poza, M.; Sanz, F.; Beceiro, A.; Chaves, F.; Bou, G. OXA-24 carbapenemase gene flanked by XerC/XerD-like recombination sites in different plasmids from different Acinetobacter species isolated during a nosocomial outbreak. Antimicrob. Agents Chemother. 2010, 54, 2427–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, F.; Quinteira, S.; Poirel, L.; Novais, A.; Peixe, L. Role of common blaOXA-24/OXA-40-carrying platforms and plasmids in the spread of OXA-24/OXA-40 among Acinetobacter species clinical isolates. Antimicrob. Agents Chemother. 2012, 56, 3969–3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, M.; Giani, T.; D’Arezzo, S.; Capone, A.; Petrosillo, N.; Visca, P.; Luzzaro, F.; Rossolini, G.M. Characterization of pABVA01, a plasmid encoding the OXA-24 carbapenemase from italian isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3528–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Povilonis, J.; Seputiene, V.; Krasauskas, R.; Juskaite, R.; Miskinyte, M.; Suziedelis, K.; Suziedeliene, E. Spread of carbapenem-resistant Acinetobacter baumannii carrying a plasmid with two genes encoding OXA-72 carbapenemase in Lithuanian hospitals. J. Antimicrob. Chemother. 2013, 68, 1000–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, G.B.; Adams-Haduch, J.M.; Bogdanovich, T.; Pasculle, A.W.; Quinn, J.P.; Wang, H.N.; Doi, Y. Identification of diverse OXA-40 group carbapenemases, including a novel variant, OXA-160, from Acinetobacter baumannii in Pennsylvania. Antimicrob. Agents Chemother. 2011, 55, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girlich, D.; Bonnin, R.A.; Bogaerts, P.; De Laveleye, M.; Huang, D.T.; Dortet, L.; Glaser, P.; Glupczynski, Y.; Naas, T. Chromosomal amplification of the blaOXA-58 carbapenemase gene in a Proteus mirabilis clinical isolate. Antimicrob. Agents Chemother. 2017, 61, e01616–e01697. [Google Scholar] [CrossRef] [Green Version]
- Boucher, H.W.; Talbot, G.H.; Benjamin, D.K.; Bradley, J.; Guidos, R.J.; Jones, R.N.; Murray, B.E.; Bonomo, R.A.; Gilbert, D. 10 × ‘20 Progress—Development of new drugs active against gram-negative bacilli: An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef]
- Mindlin, S.; Petrenko, A.; Petrova, M. Chromium resistance genetic element flanked by XerC/XerD recombination sites and its distribution in environmental and clinical Acinetobacter strains. FEMS Microbiol. Lett. 2018, 365, fny047. [Google Scholar] [CrossRef]
- Mindlin, S.; Beletsky, A.; Mardanov, A.; Petrova, M. Adaptive dif modules in permafrost strains of Acinetobacter iwoffii and their distribution and abundance among present day Acinetobacter strains. Front. Microbiol. 2019, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Girlich, D.; Damaceno, Q.S.; Oliveira, A.C.; Nordmann, P. OXA-253, a variant of the carbapenem-hydrolyzing class D beta-lactamase OXA-143 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 2976–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnin, R.A.; Girlich, D.; Jousset, A.B.; Gauthier, L.; Cuzon, G.; Bogaerts, P.; Haenni, M.; Madec, J.Y.; Couve-Deacon, E.; Barraud, O.; et al. A single Proteus mirabilis lineage from human and animal sources: A hidden reservoir of OXA-23 or OXA-58 carbapenemases in Enterobacterales. Sci. Rep. 2020, 10, 9160. [Google Scholar] [CrossRef] [PubMed]
- Cameranesi, M.M.; Moran-Barrio, J.; Limansky, A.S.; Repizo, G.D.; Viale, A.M. Site-Specific recombination at XerC/D sites mediates the formation and resolution of plasmid co-integrates carrying a blaOXA-58- and TnaphA6-Resistance module in Acinetobacter baumannii. Front. Microbiol. 2018, 9, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, H.; Butler-Cole, B.; Burgin, A.; Baker, R.; Sherratt, D.J.; Arciszewska, L.K. Functional analysis of the C-terminal domains of the site-specific recombinases XerC and XerD. J. Mol. Biol. 2003, 330, 15–27. [Google Scholar] [CrossRef]
- Subramanya, H.S.; Arciszewska, L.K.; Baker, R.A.; Bird, L.E.; Sherratt, D.J.; Wigley, D.B. Crystal structure of the site-specific recombinase, XerD. EMBO J. 1997, 16, 5178–5187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallet, B.; Arciszewska, L.K.; Sherratt, D. Reciprocal control of catalysis by the tyrosine recombinases XerC and XerD: An enzymatic switch in site-specific recombination. Mol. Cell 1999, 4, 949–959. [Google Scholar] [CrossRef]
- Spiers, A.J.; Sherratt, D.J. C-terminal interactions between the XerC and XerD site-specific recombinases. Mol. Microbiol. 1999, 32, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.; Sherratt, D.; Arciszewska, L. Switching catalytic activity in the XerCD site-specific recombination machine. J. Mol. Biol. 2001, 312, 45–57. [Google Scholar] [CrossRef]
- Summers, D.K.; Sherratt, D.J. Multimerization of high copy number plasmids causes instability: CoIE1 encodes a determinant essential for plasmid monomerization and stability. Cell 1984, 36, 1097–1103. [Google Scholar] [CrossRef]
- Sarno, R.; McGillivary, G.; Sherratt, D.J.; Actis, L.A.; Tolmasky, M.E. Complete nucleotide sequence of Klebsiella pneumoniae multiresistance plasmid pJHCMW1. Antimicrob. Agents Chemother. 2002, 46, 3422–3427. [Google Scholar] [CrossRef] [Green Version]
- Arciszewska, L.K.; Baker, R.A.; Hallet, B.; Sherratt, D.J. Coordinated control of XerC and XerD catalytic activities during Holliday junction resolution. J. Mol. Biol. 2000, 299, 391–403. [Google Scholar] [CrossRef]
- Colloms, S.D.; McCulloch, R.; Grant, K.; Neilson, L.; Sherratt, D.J. Xer-mediated site-specific recombination in vitro. EMBO J. 1996, 15, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.; Abe, T.; Arnaud, M.B.; Berlyn, M.K.; Blattner, F.R.; Chaudhuri, R.R.; Glasner, J.D.; Horiuchi, T.; Keseler, I.M.; Kosuge, T.; et al. Escherichia coli K-12: A cooperatively developed annotation snapshot—2005. Nucleic Acids Res. 2006, 34, 1–9. [Google Scholar] [CrossRef]
- Martinez, J.; Fernandez, J.S.; Liu, C.; Hoard, A.; Mendoza, A.; Nakanouchi, J.; Rodman, N.; Courville, R.; Tuttobene, M.R.; Lopez, C.; et al. Human pleural fluid triggers global changes in the transcriptional landscape of Acinetobacter baumannii as an adaptive response to stress. Sci. Rep. 2019, 9, 17251. [Google Scholar] [CrossRef]
- Tran, T.; Sherratt, D.J.; Tolmasky, M.E. fpr, a deficient Xer recombination site from a Salmonella plasmid, fails to confer stability by dimer resolution: Comparative studies with the pJHCMW1 mwr site. J. Bacteriol. 2010, 192, 883–887. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.; Merino, M.; Viedma, E.; Poza, M.; Sanz, F.; Otero, J.R.; Chaves, F.; Bou, G. Multidrug-resistant Acinetobacter baumannii Harboring OXA-24 carbapenemase, Spain. Emerg. Infect. Dis. 2011, 17, 1064–1067. [Google Scholar] [CrossRef] [PubMed]
- Larouche, A.; Roy, P.H. Effect of attC structure on cassette excision by integron integrases. Mob DNA 2011, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, J.A.; Loot, C.; Nivina, A.; Mazel, D. The Integron: Adaptation on demand. Microbiol. Spectr. 2015, 3. MDNA3-0019-2014. [Google Scholar]
- Vallenet, D.; Nordmann, P.; Barbe, V.; Poirel, L.; Mangenot, S.; Bataille, E.; Dossat, C.; Gas, S.; Kreimeyer, A.; Lenoble, P.; et al. Comparative analysis of Acinetobacters: Three genomes for three lifestyles. PLoS ONE 2008, 3, e1805. [Google Scholar] [CrossRef]
- Blakely, G.; Sherratt, D. Determinants of selectivity in Xer site-specific recombination. Genes Dev. 1996, 10, 762–773. [Google Scholar] [CrossRef] [Green Version]
- Grainge, I.; Bregu, M.; Vazquez, M.; Sivanathan, V.; Ip, S.C.; Sherratt, D.J. Unlinking chromosome catenanes in vivo by site-specific recombination. EMBO J. 2007, 26, 4228–4238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimokawa, K.; Ishihara, K.; Grainge, I.; Sherratt, D.J.; Vazquez, M. FtsK-dependent XerCD-dif recombination unlinks replication catenanes in a stepwise manner. Proc. Natl. Acad. Sci. USA 2013, 110, 20906–20911. [Google Scholar] [CrossRef] [Green Version]
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New treatment options against carbapenem-resistant Acinetobacter baumannii infections. Antimicrob. Agents Chemother. 2019, 63, e01110–e01118. [Google Scholar] [PubMed] [Green Version]
- Hartstein, A.I.; Rashad, A.L.; Liebler, J.M.; Actis, L.A.; Freeman, J.; Rourke, J.W., Jr.; Stibolt, T.B.; Tolmasky, M.E.; Ellis, G.R.; Crosa, J.H. Multiple intensive care unit outbreak of Acinetobacter calcoaceticus subspecies anitratus respiratory infection and colonization associated with contaminated, reusable ventilator circuits and resuscitation bags. Am. J. Med. 1988, 85, 624–631. [Google Scholar] [CrossRef]
- Summers, D.K.; Sherratt, D.J. Resolution of ColE1 dimers requires a DNA sequence implicated in the three-dimensional organization of the cer site. EMBO J. 1988, 7, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Cornet, F.; Mortier, I.; Patte, J.; Louarn, J.M. Plasmid pSC101 harbors a recombination site, psi, which is able to resolve plasmid multimers and to substitute for the analogous chromosomal Escherichia coli site dif. J. Bacteriol. 1994, 176, 3188–3195. [Google Scholar] [CrossRef] [Green Version]
- Spiers, A.J.; Sherratt, D.J. Relating primary structure to function in the Escherichia coli XerD site-specific recombinase. Mol. Microbiol. 1997, 24, 1071–1082. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistue, A.J.; Zorreguieta, A.; Centron, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Traglia, G.M.; Chua, K.; Centron, D.; Tolmasky, M.E.; Ramirez, M.S. Whole-genome sequence analysis of the naturally competent Acinetobacter baumannii clinical isolate A118. Genome Biol. Evol. 2014, 6, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef]
- Chang, A.C.; Cohen, S.N. Construction and characterization of amplifiable multicopy DNA cloning vehicles derived from the P15A cryptic miniplasmid. J. Bacteriol. 1978, 134, 1141–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.N.; Chang, A.C.; Hsu, L. Nonchromosomal antibiotic resistance in bacteria: Genetic transformation of Escherichia coli by R-factor DNA. Proc. Natl. Acad. Sci. USA 1972, 69, 2110–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakely, G.; May, G.; McCulloch, R.; Arciszewska, L.K.; Burke, M.; Lovett, S.T.; Sherratt, D.J. Two related recombinases are required for site-specific recombination at dif and cer in E. coli K12. Cell 1993, 75, 351–361. [Google Scholar] [CrossRef]
- Nunes-Duby, S.E.; Radman-Livaja, M.; Kuimelis, R.G.; Pearline, R.V.; McLaughlin, L.W.; Landy, A. Gamma integrase complementation at the level of DNA binding and complex formation. J. Bacteriol. 2002, 184, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Summers, D. Timing, self-control and a sense of direction are the secrets of multicopy plasmid stability. Mol. Microbiol. 1998, 29, 1137–1145. [Google Scholar] [CrossRef] [PubMed]





| Name | Sequence |
|---|---|
| ODN1 | A(A/A)TT(A/G)(A/C)CATAAG(G/G)(C/C)G(T/C)(A/A)TTATGTTAATT |
| ODN2 | AATTAACATAAGGCGTATTATGTTAATT |
| ODN3 | ACTTCGTATAATCGCCATTATGTTAAAT |
| ODN4 | ATTTCGCATAAGGCGTATTATGCGAAAT |
| ODN5 | AATTAACATAAGGCGTATTATGTTAATT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, D.L.; Traglia, G.M.; Baker, R.; Sherratt, D.J.; Ramirez, M.S.; Tolmasky, M.E. Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes. Antibiotics 2020, 9, 405. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070405
Lin DL, Traglia GM, Baker R, Sherratt DJ, Ramirez MS, Tolmasky ME. Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes. Antibiotics. 2020; 9(7):405. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070405
Chicago/Turabian StyleLin, David L., German M. Traglia, Rachel Baker, David J. Sherratt, Maria Soledad Ramirez, and Marcelo E. Tolmasky. 2020. "Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes" Antibiotics 9, no. 7: 405. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070405