Virulence Characteristics and Antibiotic Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Diverse Sources


Department of Food Science and Biotechnology, College of Agriculture and Life Sciences, Kangwon National University, Chuncheon 200-701, Korea
*
Author to whom correspondence should be addressed.
Antibiotics 2020, 9(9), 587; https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090587
Submission received: 17 July 2020
/
Revised: 2 September 2020
/
Accepted: 7 September 2020
/
Published: 8 September 2020
(This article belongs to the Special Issue Discovery and Functional Evaluation of Antimicrobials)
Abstract
:Shiga toxin-producing Escherichia coli (STEC) is an enteric pathogen that causes several gastrointestinal ailments in humans across the world. STEC’s ability to cause ailment is attributed to the presence of a broad range of known and putative virulence factors (VFs) including those that encode Shiga toxins. A total of 51 E. coli strains belonging to serogroups O26, O45, O103, O104, O113, O121, O145, and O157 were tested for the presence of nine VFs via PCR and for their susceptibility to 17 frequently used antibiotics using the disc diffusion method. The isolates belonged to eight different serotypes, including eight O serogroups and 12 H types. The frequency of the presence of key VFs were stx1 (76.47%), stx2 (86.27%), eae (100%), ehxA (98.03%), nleA (100%), ureC (94.11%), iha (96.07%), subA (9.80%), and saa (94.11%) in the E. coli strains. All E. coli strains carried seven or more distinct VFs and, among these, four isolates harbored all tested VFs. In addition, all E. coli strains had a high degree of antibiotic resistance and were multidrug resistant (MDR). These results show a high incidence frequency of VFs and heterogeneity of VFs and MDR profiles of E. coli strains. Moreover, half of the E. coli isolates (74.5%) were resistant to > 9 classes of antibiotics (more than 50% of the tested antibiotics). Thus, our findings highlight the importance of appropriate epidemiological and microbiological surveillance and control measures to prevent STEC disease in humans worldwide.
1. Introduction
Escherichia coli is ubiquitous in nature, often found in soil, water, food, human, and animal intestinal tracts. However, E. coli can also act as a pathogen in a wide range of conditions, from enteric diseases to extraintestinal infections. E. coli strains which cause enteric diseases or diarrhea are known as diarrheagenic E. coli (DEC) and are divided into six distinct pathotypes based on clinical, epidemiological, and molecular criteria [1]. Among them, enteropathogenic E. coli (EPEC) is the predominant cause of diarrhea in developing countries [1,2,3,4,5] and enterohemorrhagic E. coli (EHEC) is attributed to the foodborne outbreaks in developed countries and can cause bloody diarrhea due to the production of Shiga toxins (stxs) and is known as Shiga toxin-producing E. coli (STEC). Among the DEC, STEC includes the most dangerous strains and more than 400 serotypes that produce stxs have been identified, and this term was created since E. coli species possess the toxin, which is more or less identical to that produced by Shigella dysentery type I [6]. STEC possesses a broad range of virulence factors (VFs), which are encoded by chromosomal genes, and they are often located in pathogenicity islands (PAIs) or plasmids, with the production of stxs being the most crucial resulting in endothelial cell damage and possible hemolytic uremic syndrome (HUS) [7,8]. In addition, STEC is responsible for 2.8 million illnesses, and 3890 cases per year of enteric disease in humans, globally [8]. Shiga toxin is classified into two types, Shiga toxin type 1 (stx1) and Shiga toxin type 2 (stx2) and subtypes which dominate the pathogenicity of STEC, and it is known to be an important factor in differentiating the severity of illness [2,9,10], but cannot be solely responsible for full pathogenicity, as a vast arsenal of VFs are essential for STEC pathogenicity. Thus, STEC possesses other VFs necessary for infection, such as intimin (eae), which is an outer membrane encoded by the locus for enterocyte effacement (LEE) and is essential for the intimate adherence of eae-positive STEC strains to the host’s intestine and, eventually, for the attaching and effacing (A/E) lesions frequently perceived in STEC infections. In addition, several STEC strains contain a number of plasmid-encoded VFs, including enterohemolysin (pO157; ehxA). The toxin ehxA is a heat-labile pore-forming toxin that causes hemolysis of host red blood cells and the possession of ehxA by a STEC strain has been attributed to HUS [11]. Other plasmid-encoded (pO113) VFs include a subtilase cytotoxin gene (subA; triggers apoptosis in human cells) and an autoagglutinating adhesin gene (saa; associated with the absence of LEE) [12] also recognized as key plasmid-encoded VFs. The gene subA suppresses the host’s immune system and allows STEC to adhere to enterocytes. Growing evidence suggests that differences in virulence between pathogenic and nonpathogenic bacterial strains can be attributed to VFs in pathogenicity islands [13]. Several PAIs of STEC, including genes encoding OI-43/48, OI-57, and OI-71, are absent in nonpathogenic E. coli and are viewed as STEC VFs. These genes have been used in molecular risk assessment research to classify STEC serotypes into various seropathotypes depending on whether a specific serotype in humans has been involved in mild, severe ailment, or no disease at all. Furthermore, numerous genes located on OI-43/48, including IrgA homolog adhesin (encodes an adhesin; iha) and ureC (encode urease resistance), and on OI-71, including nleA (effector; disrupts protein secretion) are considered appropriate virulence markers in STEC serotypes involved in severe human ailments and outbreaks but their individual role in the pathogenesis of human infection is still poorly understood [2]. In addition, these genes are mainly found in high risk (HUS) STEC strains and are associated with colonization and survival in the host, and may interfere with signaling pathways during inflammation [14].
In the last decade, the STEC has gained substantial attention as a global public health concern for various sporadic infections and major outbreaks due to the emergence of resistance to multiple antibiotics as there are fewer, or often no, effective antibiotics available for infections caused by such bacteria. Furthermore, multidrug resistance (MDR) in E. coli strains has increased in recent years, causing serious problems in healthcare settings worldwide [1,15,16]. MDR represents one of the notable global health challenges of this century and the global increase in its spread is a major issue, particularly in developing countries where there is limited control over the quality, distribution, and use of antibiotics in human medicine, veterinary medicine, and food animal farming [17]. MDR bacterial infections have increased at an alarming rate due to the tremendous dissemination of antibiotic resistance determinants [18,19,20] and MDR pathogens and infections thereof are expected to cause 10 million deaths per year by 2050 [21,22]. Even though the role of commensal bacteria in providing antibiotic resistance has long been recognized, however, these bacteria have not been extensively studied [18]. E. coli comprises only a small fraction of the human gastrointestinal tract’s bacterial flora, but it is not a significant reservoir. Resistance in commensal E. coli from healthy patients was first demonstrated over 50 years ago, and several recent studies have demonstrated the high or increased incidence of MDR in commensal E. coli from healthy children and adults from various countries [23,24,25]. Currently, the use of antibiotics to treat STEC infection in humans is controversial and not recommended in many countries according to the current clinical guidelines, as some antibiotics can induce the production of Shiga toxin [8,26,27]. However, antibiotic resistance is a matter of growing concern due to the wide spread of E. coli resistance to all antibiotics used in human therapy, and the dispersion of resistance via mobile genetic elements. Additionally, STEC bacteriophages may carry antibiotic resistance-encoding genes that can be transferred to naïve E. coli which are then transformed into antibiotic-resistant strains [8,21].
Hence, bacteria with VFs and antibiotic resistance should be carefully monitored. Currently, there is limited information available on the prevalence of antibiotic resistance in E. coli isolates from humans and food samples. Understanding the prevalence of resistance to antibiotics, especially for critically important antibiotics, in E. coli isolates from human and food samples, will provide the useful information for developing risk management options to mitigate the spread of resistance. This research was performed to investigate the diversity and distribution of the major virulence-associated factors (stx1, stx2, iha, ureC, eae, uidA, nleA, ehxA, subA, and saa) in previously isolated strains from diverse sources including foods, animal carcasses and feces, and humans by using PCR. The second objective was to determine the strains’ potential as human pathogens by employing 17 frequently used antibiotics in clinical practices. This study emphasizes on the importance of preventing the spread of E. coli isolates that harbor both antibiotic resistance and virulence genes and the overall objective of this research was to contribute to STEC surveillance and gain insight into STEC strains’ molecular epidemiology in human diseases.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
A total of 51 E. coli strains were investigated, obtained from the US Food Fermentation Laboratory Culture Collection (USDA ARS, Raleigh, N.C., USA) and the USDA ARS Eastern Regional Research Center (Wyndmoor, Pa., USA). The strains were derived from humans (n = 40), domestic animals (n = 9), and food (n = 2). The E. coli and non-targeted reference strains that were used in this study are described in detail in Table 1. All strains were routinely propagated at 37 °C in Luria–Bertani (LB) broth (Difco, Becton, Dicknison, MD, USA) under aerobic conditions.
2.2. DNA Extraction
The genomic DNA was extracted using a GeneAll® ExgeneTM Cell SV genomic DNA purification kit (GeneAllTM, Seoul, Republic of Korea) according to the manufacturer’s instructions from 1 mL overnight culture of STEC strains grown at 37 °C. The concentrations of extracted DNA were determined using Eppendorf Biospectrometer® fluorescence (Eppendorf, Hamburg, Germany) at 260 nm. The genomic DNA was stored at −20 °C until use.
2.3. Detection of Virulence Genes
All 51 isolates were screened for genetic markers of virulence associated with STEC by conventional polymerase chain reaction (PCR; Mygenie32 Thermal Block, Bioneer Co., Daejeon, Korea) using primers and conditions described previously for targeting bacteriophage-encoded Shiga toxin genes (stx1, stx2), an attaching and effacing gene (eae), putative adhesin genes (saa, iha), a toxicity gene (subA), and a plasmid-encoded virulence factor (ehxA). The strains were also characterized by the use of O71 (MRA; based on the presence of various O-island nle genes) associated with pathogenicity molecular risk assessment. The primers for the detection of targeted genes have been described previously and are listed in Table 2. The uidA marker was use to confirm that all were E. coli. The E. coli strain ATCC 35150 served as a positive control for STEC strains and K-12 (ATCC 700926; MG1655) served as a negative control for the nine VFs investigated in this work. The PCR assay was carried out using Accupower Taq PCR PreMix (Bioneer Co., Daejeon, Korea) in a total volume of 20 µL comprising 2 µL of DNA template. Amplified PCR products were loaded into single wells of 2% agarose gel containing SafeViewTM (ABM, Richmond, BC, Canada). After subsequent electrophoresis using an electrophoresis system (Mupid-exU, Mupid, Tokyo, Japan) at 100 V for 30 min in 1× Tris-acetate-EDTA (TAE) buffer, the gel was visualized using a UV transilluminator (Gel Doc 2000; Bio-Rad, Hercules, CA, USA), where a 100-bp DNA ladder (ThermoFisher Scientific, Seoul, Korea) was used in each agarose run and used as a molecular weight marker.
2.4. Antibiotic Susceptibility Testing
The antibiotic susceptibility of the E. coli strains as per the guidelines of the Clinical and Laboratory Standards Institute (CLSI) were examined using a disc diffusion method [35]. The following 17 antibiotics were tested: ampicillin, methicillin, penicillin, vancomycin, erythromycin, clindamycin, ciprofloxacin, chloramphenicol, gentamicin, imipenem, meropenem, streptomycin, tetracycline, kanamycin, nalidixic acid, novobiocin, and tigecycline. E. coli ATCC 25922, which is sensitive to all the drugs, was used as the control strain [6,36,37,38,39]. The results were used to classify the strains as resistant or susceptible to a specific antibiotic using standard reference values recommended by the CLSI National Committee [35]. The strains were classified as MDR when they presented resistance to ≥ 1 antibiotic in 3 ≥ antibiotic categories [40].
3. Results
3.1. Virulence Gene Profile of STEC Strains from Diverse Resources
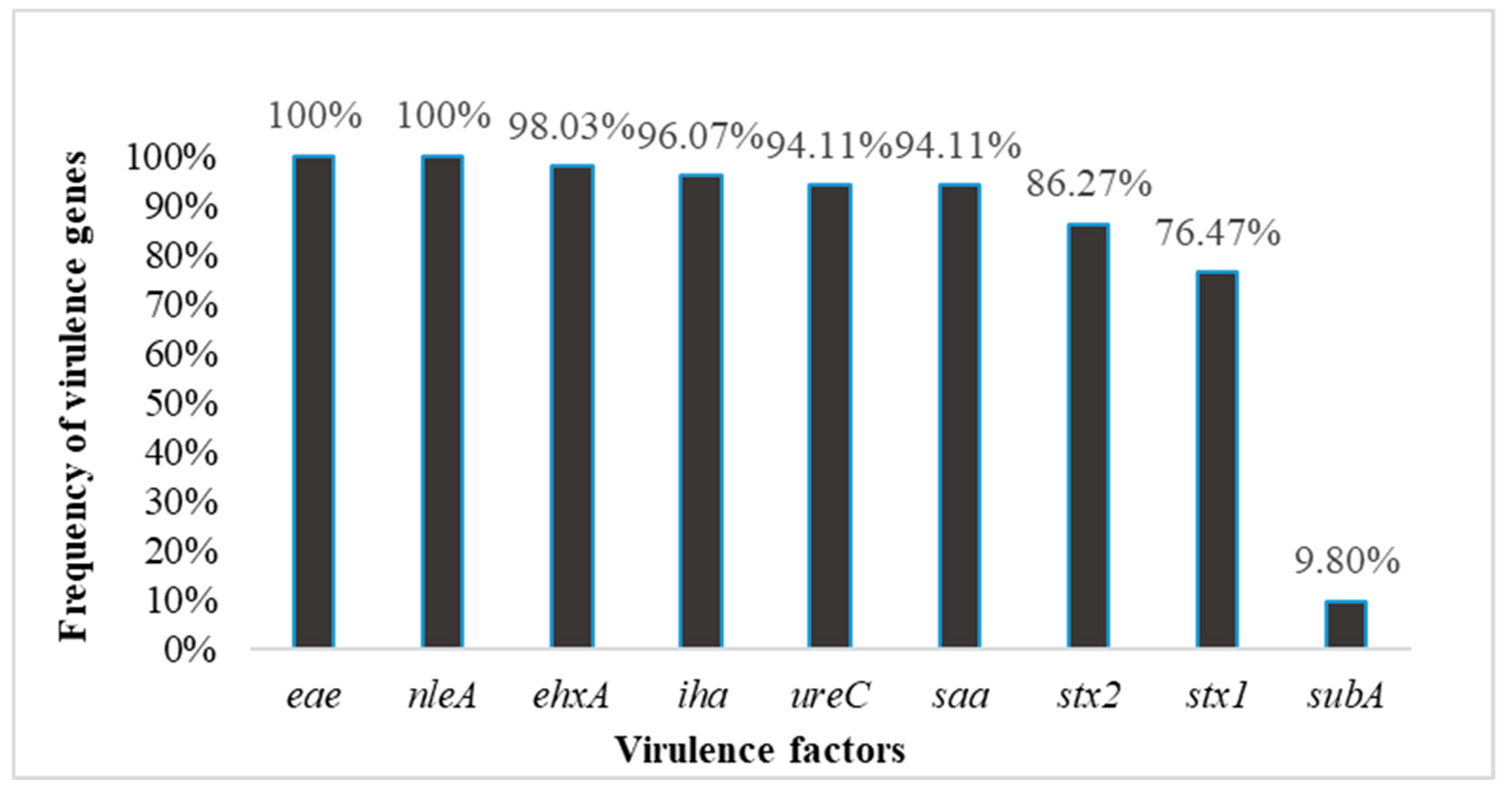
The isolated strains belonged to eight different serotypes, including eight O serogroups and 12 H types: 13.7% belonged to each serotype except O121 (12%) and O157 (5.88%) (Table 1). The strains expressed 11 different H antigens: H- (3.92%), H2 (15.6%), H4 (5.88%), H6 (1.96%), H7 (7.84%), H8 (5.88%), H11 (17.6%), H12 (3.92%), H19 (3.92%), H21 (1.92%), H25 (3.92%), and H-nonmotile (NM; 27.4%). The E. coli strains analyzed in this study were isolated from various sources, with the majority isolated from humans (Table 1). The presence of nine toxin-encoding genes was investigated using PCR in all E. coli strains. PCR revealed that all strains tested positive for eae, nleA, and uidA genes. The detection rate of stx1 and stx2 was 76.47 and 86.27%, respectively. The prevalences of the plasmid-encoded genes were as follows: iha (96.07%), saa (94.11%), ehxA (98.03%), subA (9.80%), and ureC (94.11%) (Figure 1). Our findings revealed that most of the E. coli isolates contained multiple and heterogeneous VFs and different gene combinations were observed in our investigation, as recorded in Table 3.
3.2. Antibiotic Resistance of STEC Strains
The resistance profiles of the E. coli strains against the tested antibiotics are described in Table 4. All antibiotic susceptibility results were interpreted using the breakpoints of the CLSI guidelines. Among the 17 tested antibiotics, resistance was most frequent for gentamicin (50/51, 98.03%) and novobiocin (49/51, 96.07%), followed by kanamycin (39/51, 76.47%), streptomycin (42/51, 82.35%), ampicillin (37/51, 72.54%), and tetracycline (22/51, 43.13%). Meanwhile, all the strains were resistant against methicillin, penicillin, vancomycin, and erythromycin. Additionally, all E. coli strains were susceptible to imipenem and meropenem. The high prevalence of antibiotic susceptibility was detected for nalidixic acid (50/51, 98.0%), followed by chloramphenicol (43/51, 84.3%), tetracycline (19/51, 37.25%), ciprofloxacin (17/51, 33.33%), clindamycin (15/51, 29.41%), and tigecycline (13/51, 25.49%). Moreover, half of the E. coli isolated (38/51, 74.5%) were resistant to > 9 classes of antibiotics (more than 50% of the tested antibiotics) (Table 5).
3.3. Frequency of Virulence Factor Occurrence in Isolated E. coli Strains Exhibiting Antibiotic Resistance
The frequencies of VF occurrence in isolated E. coli strains exhibiting antibiotic resistance are detailed in Table 6. The frequencies for eae, ehxA, and nleA among the resistant E. coli isolates were nearly > 98%, whereas those of ureC and saa were > 95%. Moreover, the frequencies of iha, stx2, stx1, and subA in the resistant isolates were higher than 90, 86, 80, and 9%, respectively.
4. Discussion
The concept of molecular risk assessment [41] has been used effectively to classify STEC strains into those that are attributed to outbreaks and life-threatening diseases in humans and those causing less severe disease or that are not involved in human disease. E. coli strains have been characterized by serotype and the presence and subtype of VFs, as well as other toxins and plasmid-associated adherence and virulence factors. Thus, in this study, we investigated the presence of VFs in E. coli strains to broaden the knowledge of the properties of E. coli strains isolated from diverse sources. Recent epidemiological studies have revealed that the STEC serotypes O26, O103, O111, O145, and O157 are highly related to human infections (they may account for up to 80% of human STEC infections) [8] and our results are in agreement with reports from other countries describing the high pathogenic potential of strains associated with these serogroups [2]. The presence of non-LEE effector genes encoded by O-island O1-71 is highly attributed to strains that were often involved in outbreaks and serious disease in humans [41,42,43]. We observed that stx2-harboring isolates were more frequent than stx1-harboring isolates. The dominant combinations of VFs present in the strains studied were stx1, stx2, and eae (76.4% of strains). The distribution pattern of VFs was similar to the STEC strains isolated from domestic animals in Mexico [44]. In France, the eminent toxin genotype was that of stx2-carrying STEC strains. From these findings, it seems that STEC strains carrying the stx1 gene are more often confronted than those carrying stx2. Possession of OI-43/48, OI-71, and non-LEE effectors genes together with stx2, eae and a whole plasmid is the hallmark of highly virulent STEC strains that are frequently associated with outbreaks and serious diseases such as hemorrhagic colitis (HC) and HUS [8]. Since the carriage of combinations of stx genes has been correlated with severity of the disease, the stx gene profile provides us an overview of the pathogenic potential of these STEC strains from diverse sources. STEC strains that carry both stx2 and the eae genes were more often associated with severe disease. Interestingly, all of the isolates were harboring the eae gene, and it is reported that a significant majority of human STEC isolates obtained from HC or HUS patients contained eae [45]. The presence of subA in the E. coli strains was similar to that observed in some STEC strains isolated from human infections in the USA and Australia [46]. It was previously reported that the amount of stx2 production is capable of determining the severity of diseases caused by STEC strains. Results of this characterization have identified that E. coli strains, defined by the presence of eae, subA, and the nleA genes, may be considered a significant food safety threat. Plasmid-encoded VFs enhance pathogenesis and contribute to the survival of STEC in humans [47], however, the pathogenic mechanisms of STEC infection are only partially understood. The varying prevalence of various VFs indicates that STEC strains are heterogeneous and it has been hypothesized that the combination of these genes may complement the Shiga toxin effect and enhance its virulence among STEC strains. The majority of strains in our research were positive for key VFs. Most of the strains carried the full complement of OI-43/48 VFs and all non-LEE-encoding effector genes. There is no specific pattern of virulence markers capable of interfering with the pathogenic potential of a given STEC isolate, and the search for a broad set of VFs has become the best strategy for measuring the microbiological and clinical risks that these pathogens may pose.
The 17 most frequently used antibiotics in clinical practice were employed to assess the actual frequency of antibiotic resistance in 51 E. coli strains. Generally, antibiotics are divided mainly into three categories on the basis of their functions (Table 4). While MDR was seen, however, there was no common pattern of resistance. It is important to note that all strains were resistant to at least three different classes of antibiotic agents and were considered as MDR. The majority of the E. coli strains of our research showed a high prevalence of resistance against first-line antibiotics (commonly prescribed oral antibiotics) such as ampicillin, penicillin, methicillin, gentamicin, vancomycin, novobiocin, streptomycin, kanamycin, and erythromycin (Table 4). The similar results of antibiotic resistance of E. coli strains were reported in developing countries such as Brazil, Turkey, China, and Ghana [1,4,5,45]. Our data exhibit a high resistance rate in E. coli strains that is comparable to those reported in previous studies. These results illustrate the growing extent of the misuse of antibiotics in clinical practices. In particular, E. coli strain resistance to methicillin, penicillin, vancomycin, and novobiocin reached 100%. The resistance rates of these E. coli strains were higher than reported in developing countries [3,48]. These kinds of MDR STEC strains pose a serious threat to human health by affecting treatment against them. It was previously reported that patients infected with STEC should not be treated with antibiotics due to the risk of developing HUS [26,49]. E. coli forms part of the human gut’s commensal flora and has been identified as the predominant reservoir of genes for antibiotic resistance [20]. These resistance genes are stable once acquired and are easily transferable to pathogenic bacteria [50]. These transfers have effectively changed the etiological and pathogenic character of bacterial species [51]. The majority of the STEC serotypes found in this study have also been reported in other countries. Depending on comparison by serotype and sequence type with human strains and the prevalence of VFs, the STEC strains could have a higher potential to cause human disease. Documented data showed that STEC strains isolated from human and food samples show a high prevalence of resistance to various types of antibiotics, including aminoglycosides, tetracycline, penicillin, and chloramphenicol. Molecular epidemiological studies have shown that the presence of certain antibiotic resistance genes, including the genes that encode resistance against tetracycline (tetA and tetB), ampicillin (CITM), gentamicin (aac (3)-IV), chloramphenicol (cat1 and cmlA), and aminoglycosides (aadA1), is the key cause of antibiotic resistance in STEC [52,53]. With regard to macrolide antibiotics (erythromycin), E. coli is an enteric bacterium, which are often non-susceptible due to the presence of chromosomal efflux pumps (mel) or cellular impermeability [54,55]. A number of different mechanisms have been reported for the macrolide resistance of Gram-negative bacteria. These mechanisms include the presence of a number of genes, such as two ester genes (ere(A) and ere(B)) [56,57,58], phosphorylase genes (mph(A), mph(B), and mph(D)) [59], and one rRNA methylase gene (erm(B)) [56,60,61]. In addition, if they acquire macrolide resistance genes, such as mef(A) and mef(B), that may increase their resistance levels further [49]. We found that the pattern of phenotypic resistance of STEC strains was supported by the genotypic resistance of STEC strains isolated from various samples followed by a high prevalence of antibiotic resistance genes [52]. Several studies have recorded that the prevalence of antibiotic-resistant E. coli has increased since 1950 [62]. An alarming increase in the prevalence of MDR E. coli strains all over the world has been reported and this is a result of the spread of plasmids and other genetic elements. This has made antibiotic resistance a major public health issue globally [62]. E. coli is an important food safety and public health concern because of its pathogenicity and potential for MDR.
Lastly, but most importantly, we found that E. coli strains harbor a high level of VFs in addition to high MDR (Table 3 and Table 4). These results explain how E. coli strains can effectively invade the human body and evade antibiotic treatment. The results of this research demonstrate that MDR E. coli strains harbor a high frequency of VFs and their VF profiles are highly heterogeneous. These results suggest that use of antibiotics needs to be monitored by the private, public, and agricultural sectors as certain antibiotics can induce the production of stx and thus encourage the onset of severe disease symptoms in humans. The improper use of antibiotics has become a public health problem worldwide in healthcare settings. Given the importance of E. coli in food safety and public health, our findings on the prevalence of antibiotic resistance and VFs provide valuable information for risk management strategies to protect public health. The monitoring of the antibiotic resistance of STEC is pivotal due to the likelihood of the horizontal transfer of resistance genes from notorious STEC strains to other pathogens. In addition, the monitoring process will help in developing new treatment approaches and help in establishing effective control strategies that assist in stopping the spread of resistance. Hence, the molecular typing and contentious monitoring of antibiotic resistance could be helpful in developing efficacious control strategies against STEC and in formulating new antibiotics with reduced tendency for antibiotic resistance. The diversity in the prevalence of stx genes, enterotoxin genes, and other virulence-related genes in this study and the other studies can be attributed to the geographical origin of samples, the sample size, the handling of the collected samples, the number of strains examined, the type of the examined VFs, and the role of the examined VFs in the pathogenesis of the disease. Surveillance data suggest that resistance in E. coli is consistently highest for antibiotics that have been in use in human and veterinary medicine for the longest period of time [62]. The past two decades have witnessed major increases in the emergence and spread of MDR bacteria and increasing resistance to newer compounds, such as fluoroquinolones and certain cephalosporins [62]. Thus, surveillance and control measures need to be intensified to prevent further spread of these strains in the world.
In summary, E. coli is a significant cause of diarrheal and foodborne outbreaks, resulting in severe economic losses. This research demonstrates a high prevalence and heterogeneity of VF profiles among human MDR E. coli strains. The virulotyping revealed that the majority of E. coli strains were positive for stx1, stx2, eae, ehxA, ureC, nleA, and iha but subA was observed in a very small number of isolates. The serotype O26 strains possess the highest number of virulence-associated factors. The majority of isolates were resistant to two or more antibiotics that are commonly used in clinical medicine for the treatment of various bacterial diseases. However, all the 51 isolates were sensitive to imipenem and meropenem and, therefore, these drugs could be the drugs of choice in the treatment of STEC infections. We conclude that appropriate efforts should be focused on surveillance and that control measures to prevent/reduce further the spread of such microorganisms are crucial. However, further research using whole genome sequences would therefore be required to better understand the prevalence of VFs and antibiotic resistance in E. coli strains that may arise in this important human pathogen. The continuous monitoring and screening for MDR foodborne pathogens should be performed. Taken together, this knowledge will provide a better understanding of the risks associated with STEC and will aid in the development of appropriate and tailored intervention strategies.
Author Contributions
Conceptualization, M.R and D.-H.O.; methodology, M.R.; software, M.R.; validation, M.R., and D.-H.O.; formal analysis, M.R.; investigation, M.R.; resources, D.-H.O.; data curation, M.R.; writing—original draft preparation, M.R.; writing—review and editing, M.R.; visualization M.R.; supervision, D.-H.O.; project administration, D.-H.O.; funding acquisition, D.-H.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research work was supported by a grant from the Brain Korea (BK) 21 Plus Project (Grant No. 22A20153713433) funded by the Korean Government, Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| STEC | Shiga toxin-producing Escherichia coli |
| VFs | Virulence factors |
| PCR | Polymerase chain reaction |
| MDR | Multidrug resistance |
| DEC | Diarrheagenic E. coli |
| EPEC | Enteropathogenic E. coli |
| EHEC | Enterohemorrhagic E. coli |
| HUS | Hemolytic uremic syndrome |
| LEE | Locus for enterocyte effacement |
| CLSI | Clinical and Laboratory Standards Institute |
References
- Spano, L.C.; Da Cunha, K.F.; Monfardini, M.V.; Fonseca, R.D.C.B.; Scaletsky, I.C.A. High prevalence of diarrheagenic Escherichia coli carrying toxin-encoding genes isolated from children and adults in southeastern Brazil. BMC Infect. Dis. 2017, 17, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, A.M.F.; Hernandes, R.T.; Takagi, E.H.; Guth, B.E.C.; Ori, E.D.L.; Pinheiro, S.R.S.; De Andrade, T.S.; Oliveira, S.L.; Cergole-Novella, M.C.; Francisco, G.R.; et al. Virulence profiling and molecular typing of shiga toxin-producing E. coli (STEC) from human sources in Brazil. Microorganisms 2020, 8, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, A.; Sengupta, A.; Kumar, A.; Singh, U.K.; Jaiswal, A.K.; Das, P.; Das, S. Molecular Epidemiology of Extended-Spectrum β-Lactamase–producing Escherichia coli pathotypes in diarrheal children from low socioeconomic status communities in Bihar, India: Emergence of the CTX-M type. Infect. Dis. Res. Treat. 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozcal, E.; Eldem, V.; Aydemir, S.; Skurnik, M. The relationship between phylogenetic classification, virulence and antibiotic resistance of extraintestinal pathogenic Escherichia coli in İzmir province, Turkey. PeerJ 2018, 6, e5470. [Google Scholar] [CrossRef] [Green Version]
- Forson, A.O.; Tsidi, W.B.; Nana-Adjei, D.; Quarchie, M.N.; Obeng-Nkrumah, N.; Obeng-Nkrumah, N. Escherichia coli bacteriuria in pregnant women in Ghana: Antibiotic resistance patterns and virulence factors. BMC Res. Notes 2018, 11, 901. [Google Scholar] [CrossRef]
- Amézquita-López, B.A.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Quiñones, B. Isolation, genotyping and antimicrobial resistance of Shiga toxin-producing Escherichia coli. J. Microbiol. Immunol. Infect. 2018, 51, 425–434. [Google Scholar] [CrossRef]
- Bergan, J.; Lingelem, A.B.D.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A.; Goga, B.T.C.; Ashram, S.E. Virulence characteristics and antimicrobial resistance profiles of shiga toxin-producing Escherichia coli isolates from humans in South Africa: 2006–2013. Toxins 2019, 11, 424. [Google Scholar] [CrossRef] [Green Version]
- Orth-Höller, D.; Grif, K.; Khan, A.B.; Naim, A.; Dierich, M.P.; Würzner, R. The Shiga toxin genotype rather than the amount of Shiga toxin or the cytotoxicity of Shiga toxin in vitro correlates with the appearance of the hemolytic uremic syndrome. Diagn. Microbiol. Infect. Dis. 2007, 59, 235–242. [Google Scholar] [CrossRef]
- Bai, X.; Zhao, A.; Lan, R.; Xin, Y.; Xie, H.; Meng, Q.; Jin, D.; Yu, B.; Sun, H.; Lu, S.; et al. Shiga toxin-producing Escherichia coli in Yaks (Bos grunniens) from the Qinghai-Tibetan Plateau, China. PLoS ONE 2013, 8, e65537. [Google Scholar] [CrossRef]
- Jiang, C.; An, T.; Wang, S.; Wang, G.; Si, W.; Tu, Y.; Liu, Y.; Wu, J.; Liu, S.; Cai, X. Role of the ehxA gene from Escherichia coli serotype O82 in hemolysis, biofilm formation, and in vivo virulence. Can. J. Microbiol. 2015, 61, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Srimanote, P.; Woodrow, M.C.; Paton, J.C. Characterization of saa, a novel autoagglutinating adhesin produced by locus of enterocyte effacement-negative shiga-toxigenic Escherichia coli strains that are virulent for humans. Infect. Immun. 2001, 69, 6999–7009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, W.; Shen, J.; Toro, M.; Zhao, S.; Meng, J. Distribution of pathogenicity islands OI-122, OI-43/48, and OI-57 and a high-pathogenicity island in shiga toxin-producing Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 3406–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombes, B.K.; Gilmour, M.W.; Goodman, C.D. The evolution of virulence in Non-O157 shiga toxin-producing Escherichia Coli. Front. Microbiol. 2011, 2, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, A.; Chandran, S.P.; Mahadik, K.; Macaden, R.; Lundborg, C.S. Frequency and factors associated with carriage of multi-drug resistant commensal Escherichia coli among women attending antenatal clinics in Central India. BMC Infect. Dis. 2013, 13, 199. [Google Scholar] [CrossRef]
- Zheng, S.-F.; Yu, F.; Chen, X.; Cui, D.; Cheng, Y.; Xie, G.; Yang, X.; Han, D.; Wang, Y.; Zhang, W.; et al. Enteropathogens in children less than 5 years of age with acute diarrhea: A 5-year surveillance study in the Southeast Coast of China. BMC Infect. Dis. 2016, 16, 434. [Google Scholar] [CrossRef] [Green Version]
- Adamu, M.S.; Ugochukwu, I.C.I.; Idoko, S.I.; Kwabugge, Y.A.; Abubakar, N.S.; Ameh, J.A. Virulent gene profile and antibiotic susceptibility pattern of Shiga toxin-producing Escherichia coli (STEC) from cattle and camels in Maiduguri, North-Eastern Nigeria. Trop. Anim. Health Prod. 2018, 50, 1327–1341. [Google Scholar] [CrossRef]
- Alekshun, M.; Levy, S. Commensals upon us. Biochem. Pharmacol. 2006, 71, 893–900. [Google Scholar] [CrossRef]
- Marshall, B.M.; Ochieng, D.J.; Levy, S.B. Commensals: Underappreciated reservoir of antibiotic resistance. Microbe Mag. 2009, 4, 231–238. [Google Scholar] [CrossRef]
- Bailey, J.; Pinyon, J.L.; Anantham, S.; Hall, R.M. Commensal Escherichia coli of healthy humans: A reservoir for antibiotic-resistance determinants. J. Med. Microbiol. 2010, 59, 1331–1339. [Google Scholar] [CrossRef]
- Mir, R.A.; Kudva, I.T. Antibiotic-resistant Shiga toxin-producing Escherichia coli: An overview of prevalence and intervention strategies. Zoonoses Public Health 2018, 66, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallcross, L.; Howard, S.J.; Fowler, T.; Davies, S.C. Tackling the threat of antimicrobial resistance: From policy to sustainable action. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140082. [Google Scholar] [CrossRef] [PubMed]
- Guinée, P.; Ugueto, N.; Van Leeuwen, N. Escherichia coli with resistance factors in vegetarians, babies, and nonvegetarians. Appl. Microbiol. 1970, 20, 531–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moorhouse, E.C. Transferable drug resistance in enterobacteria isolated from urban infants. BMJ 1969, 2, 405–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallecchi, L.; Lucchetti, C.; Bartoloni, A.; Bartalesi, F.; Mantella, A.; Gamboa, H.; Carattoli, A.; Paradisi, F.; Rossolini, G.M. Population structure and resistance genes in antibiotic-resistant bacteria from a remote community with minimal antibiotic exposure. Antimicrob. Agents Chemother. 2006, 51, 1179–1184. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.E.; Wilker, P.R.; Reiter, P.L.; Hedican, E.; Bender, J.B.; Hedberg, C.W. Antibiotic treatment of Escherichia coli O157 infection and the risk of hemolytic uremic syndrome, minnesota. Pediatr. Infect. Dis. J. 2012, 31, 37–41. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R.; O’Brien, A.D. New therapeutic developments against shiga toxin-producing Escherichia coli. In Environmental Microbial Forensics; American Society for Microbiology: Washington, DC, USA, 2015; pp. 341–358. [Google Scholar]
- Chou, T.C.; Chiu, H.C.; Kuo, C.J.; Wu, C.M.; Syu, W.J.; Chiu, W.T.; Chen, C.S. Enterohaemorrhagic Escherichia coli O157:H7 shiga-like toxin 1 is required for full pathogenicity and activation of the p38 mitogen-activated protein kinase pathway in C aenorhabditis elegans. Cell. Microbiol. 2013, 15, 82–97. [Google Scholar] [CrossRef]
- Anklam, K.S.; Kanankege, K.S.T.; Gonzales, T.K.; Kaspar, C.W.; Döpfer, D. Rapid and reliable detection of shiga toxin–producing Escherichia coli by real-time multiplex PCR. J. Food Prot. 2012, 75, 643–650. [Google Scholar] [CrossRef]
- Paton, A.W.; Paton, J.C. Multiplex PCR for direct detection of shiga toxigenic Escherichia coli strains producing the novel subtilase cytotoxin. J. Clin. Microbiol. 2005, 43, 2944–2947. [Google Scholar] [CrossRef] [Green Version]
- Paton, A.W.; Paton, J.C. Direct detection and characterization of shiga toxigenic Escherichia coli by multiplex PCR for stx1, stx2, eae, ehxA, and saa. J. Clin. Microbiol. 2002, 40, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Coombes, B.K.; Wickham, M.E.; Mascarenhas, M.; Gruenheid, S.; Finlay, B.B.; Karmali, M.A. Molecular analysis as an aid to assess the public health risk of non-O157 shiga toxin-producing Escherichia coli strains. Appl. Environ. Microbiol. 2008, 74, 2153–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, M.; Iida, T.; Ohnishi, M.; Kurokawa, K.; Takahashi, A.; Tsukamoto, T.; Yasunaga, T.; Hayashi, T.; Honda, T. Association of the urease gene with enterohemorrhagic Escherichia coli strains irrespective of their serogroups. J. Clin. Microbiol. 2001, 39, 4541–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janka, A.; Becker, G.; Sonntag, A.-K.; Bielaszewska, M.; Dobrindt, U.; Karch, H. Presence and characterization of a mosaic genomic island which distinguishes sorbitol-fermenting enterohemorrhagic Escherichia coli O157:H− from E. coli O157:H7. Appl. Environ. Microbiol. 2005, 71, 4875–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, R.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; Hardy, D.; Zimmer, B.; et al. CLSI methods development and standardization working group best practices for evaluation of antimicrobial susceptibility tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-L.; Hsueh, J.-Y.; Yip, B.-S.; Chih, Y.-H.; Peng, K.-L.; Cheng, J. Antimicrobial peptides display strong synergy with vancomycin against vancomycin-resistant E. faecium, S. aureus, and wild-type E. coli. Int. J. Mol. Sci. 2020, 21, 4578. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, A.; Langley, T.; Skinner, K.; Trimby, E.; Birchall, S.; Dorali, A.; Sandiford, S.; Williams, J.; Warn, P.; et al. Potentiation of antibiotic activity by a novel cationic peptide: Potency and spectrum of activity of SPR741. Antimicrob. Agents Chemother. 2017, 61, 200–217. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wu, D.; Liu, K.; Suolang, S.; He, T.; Liu, X.; Wu, C.; Wang, Y.; Lin, D. Investigation of antimicrobial resistance in Escherichia coli and enterococci isolated from tibetan pigs. PLoS ONE 2014, 9, e95623. [Google Scholar] [CrossRef] [Green Version]
- Wayne, P.A. Clinical and laboratory standards institute. In Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Seventh Informational Supplement: 2016; Clinical Laboratory Standards Institute: Wayne, PA, USA.
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.E.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Bugarel, M.; Beutin, L.; Fach, P. Low-density macroarray targeting non-locus of enterocyte effacement effectors (nle Genes) and major virulence factors of shiga toxin-producing Escherichia coli (STEC): A new approach for molecular risk assessment of STEC isolates. Appl. Environ. Microbiol. 2009, 76, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Bugarel, M.; Beutin, L.; Martin, A.; Gill, A.; Fach, P. Micro-array for the identification of Shiga toxin-producing Escherichia coli (STEC) seropathotypes associated with Hemorrhagic Colitis and Hemolytic Uremic Syndrome in humans. Int. J. Food Microbiol. 2010, 142, 318–329. [Google Scholar] [CrossRef]
- Amézquita-López, B.A.; Quiñones, B.; Lee, B.G.; Chaidez, C. Virulence profiling of Shiga toxin-producing Escherichia coli recovered from domestic farm animals in Northwestern Mexico. Front. Microbiol. 2014, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, Q.; Yu, X.; Zhang, Y.; Cao, X.; Li, D. Diversity of virulence genes in multidrug resistant Escherichia coli from a hospital in Western China [Retraction]. Infect. Drug Resist. 2020, 13, 901–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, H.J.; Sloan, J.; Bulach, D.M.; Seemann, T.; Allison, C.C.; Tauschek, M.; Robins-Browne, R.M.; Paton, J.C.; Whittam, T.S.; Paton, A.W.; et al. Shiga toxin–producing Escherichia coli strains negative for locus of enterocyte effacement. Emerg. Infect. Dis. 2009, 15, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Karama, M.; Mainga, A.O.; Cenci-Goga, B.T.; Malahlela, M.; El-Ashram, S.; Kalake, A. Molecular profiling and antimicrobial resistance of Shiga toxin-producing Escherichia coli O26, O45, O103, O121, O145 and O157 isolates from cattle on cow-calf operations in South Africa. Sci. Rep. 2019, 9, 11930. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, T.J.; Molina, M.; Lanata, C.F.; Ecker, L.; Ruiz, J.; Hall, E.; I Gil, A.; Cleary, T.G.; Del Valle, L.J.; Barletta, F.; et al. High frequency of antimicrobial drug resistance of diarrheagenic Escherichia coli in Infants in Peru. Am. J. Trop. Med. Hyg. 2009, 81, 296–301. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.S.; Mooney, J.C.; Brandt, J.R.; Staples, A.O.; Jelacic, S.; Boster, D.R.; Watkins, S.L.; Tarr, P.I. Risk factors for the hemolytic uremic syndrome in children infected with Escherichia coli O157:H7: A multivariable analysis. Clin. Infect. Dis. 2012, 55, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Bouzari, S.; Farhang, E.; Hosseini, S.M.; Alikhani, M.Y. Prevalence and antimicrobial resistance of shiga toxin-producing Escherichia coli and enteropathogenic Escherichia coli isolated from patients with acute diarrhea. Iran. J. Microbiol. 2018, 10, 151–157. [Google Scholar]
- Oporto, B.; Ocejo, M.; Alkorta, M.; Marimón, J.M.; Montes, M.; Hurtado, A. Zoonotic approach to Shiga toxin-producing Escherichia coli: Integrated analysis of virulence and antimicrobial resistance in ruminants and humans. Epidemiol. Infect. 2019, 147, e164. [Google Scholar] [CrossRef] [Green Version]
- Lutz, E.A.; Mccarty, M.J.; Mollenkopf, D.F.; Funk, J.A.; Gebreyes, W.A.; Wittum, T.E. Ceftiofur use in finishing swine barns and the recovery of fecal Escherichia coli or Salmonella spp. resistant to ceftriaxone. Foodborne Pathog. Dis. 2011, 8, 1229–1234. [Google Scholar] [CrossRef]
- Liu, J.; Keelan, P.; Bennett, P.M.; Enne, V.I. Characterization of a novel macrolide efflux gene, mef(B), found linked to sul3 in porcine Escherichia coli. J. Antimicrob. Chemother. 2009, 63, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Graves, A.K. Genetic diversity and antimicrobial resistance of Escherichia coli from human and animal sources uncovers multiple resistances from human sources. PLoS ONE 2011, 6, e20819. [Google Scholar] [CrossRef] [Green Version]
- Ojo, K.K.; Ulep, C.; Van Kirk, N.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M.C. The mef(A) gene predominates among seven macrolide resistance genes identified in gram-negative strains representing 13 genera, isolated from healthy portuguese children. Antimicrob. Agents Chemother. 2004, 48, 3451–3456. [Google Scholar] [CrossRef] [Green Version]
- Ounissi, H.; Courvalin, P. Nucleotide sequence of the gene ereA encoding the erythromycin esterase in Escherichia coli. Gene 1985, 35, 271–278. [Google Scholar] [CrossRef]
- Arthur, M.; Autissier, D.; Courvalin, P. Analysis of the nucleotide sequence of theereBgene encoding the erythromydn esterase type II. Nucleic Acids Res. 1986, 14, 4987–4999. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, N.; Katayama, J.; O’Hara, K. Cloning and nucleotide sequence of themphBgene for macrolide 2′-phosphotransferase II inEscherichia coli. FEMS Microbiol. Lett. 1996, 144, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob. Agents Chemother. 2011, 55, 4224–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappell, A.D.; Denies, M.S.; Ahuja, N.H.; Ledeboer, N.A.; Newton, R.J.; Hristova, K.R. Detection of multi-drug resistant Escherichia coli in the urban waterways of Milwaukee, WI. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, D.A.; Zhao, S.; Tong, E.; Ayers, S.; Singh, A.; Bartholomew, M.J.; McDermott, P.F. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. Emerg. Infect. Dis. 2012, 18, 741–749. [Google Scholar] [CrossRef]
- Adamu, M.S.; Ugochukwu, I.C.I.; Idoko, S.I.; Kwabugge, Y.A.; Abubakar, N.S.; Ameh, J.A. Virulent gene profile and antimicrobial susceptibility pattern of Shiga toxin-producing Escherichia coli (STEC) from humans in Maiduguri, Borno State, North-Eastern Nigeria. Comp. Haematol. Int. 2017, 27, 341–351. [Google Scholar] [CrossRef]
Figure 1.
Virulence-associated factor profiles of STEC isolates from diverse sources.


{kind=link}
Table 1.
List of Shiga toxin-producing E. coli isolates analyzed in this study.
| ID No. | Serotype | Source |
|---|---|---|
| B444 | O26:H11 | H |
| B445 | O26:H11 | H |
| B446 | O26:H11 | H |
| B447 | O26:H11 | H |
| B448 | O26:H11 | H |
| B449 | O26:H11 | H |
| B450 | O26:H11 | H |
| B451 | O45:NM | Co (calf) |
| B452 | O45:H2 | D |
| B453 | O45:H2 | H |
| B454 | O45:H2 | Go |
| B455 | O45:H2 | R |
| B456 | O45:H12 | Co |
| B457 | O45:H2 | Co (calf) |
| B458 | O103:H2 | H |
| B459 | O103:H25 | H |
| B460 | O103:H25 | H |
| B461 | O103:H2 | H |
| B462 | O103:H11 | H |
| B463 | O103:H6 | H |
| B464 | O103:H11 | H |
| B465 | O104:H4 | H |
| B466 | O104:H4 | H |
| B467 | O104:H21 | H |
| B468 | O104:H7 | Ca |
| B469 | O104:H4 | H |
| B470 | O104:H2 | Co (feces) |
| B471 | O104:H12 | Co |
| B472 | O111:H- | H |
| B473 | O111:NM | H |
| B474 | O111:NM | H |
| B475 | O111:NM | H |
| B476 | O111:H8 | H |
| B477 | O111:H8 | H |
| B478 | O111:H8 | H |
| B479 | O121:NM | H |
| B480 | O121:H19 | H |
| B481 | O121:NM | H |
| B482 | O121:H19 | H |
| B483 | O121:NM | H |
| B484 | O121:NM | H |
| B485 | O145:NM | H |
| B486 | O145:NM | H |
| B487 | O145:H- | H |
| B488 | O145:NM | H |
| B489 | O145:NM | Co |
| B490 | O145:NM | H |
| B491 | O145:NM | H |
| B492 | O157:H7 | H |
| B493 | O157:H7 | H |
| B494 | O157:H7 | Gb |
Table 2.
List of DNA oligonucleotides used in this study for PCR amplification.
| Target Gene | Primer | Nucleotide Sequence (5′—3′) | PCR Conditions | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| stx1 | stx1-F | GAAAGCGATGCAGCTATTA | 95 °C for 15 min, 95 °C for 30 s, 60 °C for 40 s, 72 °C for 30 s, 40 cycles, 72 °C for 3 min | 789 | [28] |
| stx1-R | GGATAATTTGTTTGCAGTTG | ||||
| stx2 | stx2-F | TATTATTTAAATGGGTACTGTGC | 1073 | ||
| stx2-R | ATGTGTCATCCTCATTATACTTG | ||||
| eae | eae-F | CATTGATCAGGATTTTTCTGGTGATA | 102 | [29] | |
| eae-R | CTCATGCGGAAATAGCCGTTA | ||||
| ehxA | ehxA-F | CGTTAAGGAACAGGAGGTGTCAGTA | 142 | ||
| ehxA-R | ATCATGTTTTCCGCCAATGAG | ||||
| subA | subA-F | TATGGCTTCCCTCATTGCC | 94 °C for 5 min, 94 °C for 45 s, 50 °C for 30 s, 72 °C for 30 s, 30 cycles, 72 °C for 3 min | 556 | [30] |
| subA-R | TATAGCTGTTGCTTCTGACG | ||||
| Saa | saa-F | CGTGATGAACAGGCTATTGC | 94 °C for 5 min, 94 °C for 45 s, 56 °C for 30 s, 72 °C for 30 s, 30 cycles, 72 °C for 7 min | 119 | [31] |
| saa-R | ATGGACATGCCTGTGGCAAC | ||||
| nleA | nleA-F | ATGAACATTCAACCGACCATAC | 94 °C for 5 min, 94 °C for 30 s, 55 °C for 60 s, 35 cycles, 72 °C for 2.5 min | 1296 | [32] |
| nleA-R | GACTCTTGTTTCTTGGATTATATCAAA | ||||
| ureC | ureC-F | TCTAACGCCACAACCTGTAC | 94 °C for 3 min, 94 °C for 60 s, 60 °C for 60 s, 72 °C for 60 s, 35 cycles, 72 °C for 2.5 min | 397 | [33] |
| ureC-R | GAGGAAGGCAGAATATTGGG | ||||
| iha | iha-F | CAGTTCAGTTTCGCATTCACC | 95 °C for 15 min, 94 °C for 30 s, 55 °C for 60 s, 72 °C for 60 s, 30 cycles, 72 °C for 5 min | 1305 | [34] |
| iha-R | GTATGGCTCTGATGCGATG |
Table 3.
Pattern of distribution of essential chromosomal and plasmid-encoded virulence-associated factors in STEC isolates from diverse sources.
Table 3.
Pattern of distribution of essential chromosomal and plasmid-encoded virulence-associated factors in STEC isolates from diverse sources.
| No. of Virulence Genes | Virulence Gene Profile | No. (%) of Bacterial Strains | Total No. (%) |
|---|---|---|---|
| 6 genes | stx1, stx2, eae, ehxA, saa, nleA | 2 (3.92) | 4 (7.84) |
| stx1, stx2, eae, nleA, iha, ureC | 1 (1.96) | ||
| eae, nleA, ehxA, saa, iha, ureC | 1 (1.96) | ||
| 7 genes | stx1, stx2, eae, ehxA, saa, iha, nleA | 1 (1.96) | 19 (37.25) |
| stx1, eae, ehxA, saa, iha, nleA, ureC | 6 (11.76) | ||
| stx2, eae, ehxA, saa, iha, nleA, ureC | 10 (19.60) | ||
| stx1, stx2, eae, ehxA, iha, nleA, ureC | 2 (3.92) | ||
| 8 genes | stx1, stx2, eae, ehxA, saa, iha, nleA, ureC | 23 (45.09) | 24 (47.05) |
| stx2, eae, ehxA, saa, iha, nleA, ureC, subA | 1 (1.96) | ||
| 9 genes | stx1, stx2, eae, ehxA, iha, nleA, ureC, subA | 4 (7.84) | 4 (7.84) |
Table 4.
Multidrug resistance patterns of STEC isolates from diverse sources.
| Antibiotic | Resistant n (%) | Intermediate n (%) | Susceptible n (%) |
|---|---|---|---|
| Bacterial protein synthesis | |||
| Gentamicin | 50 (98.03) | 0 (0) | 1 (1.96) |
| Kanamycin | 39 (76.47) | 5 (9.8) | 7 (13.72) |
| Streptomycin | 42 (82.35) | 7 (13.72) | 2 (3.92) |
| Erythromycin | 51 (100) | 0 (0) | 0 (0) |
| Tetracycline | 22 (43.13) | 10 (19.6) | 19 (37.25) |
| Clindamycin | 17 (33.33) | 19 (37.25) | 15 (29.41) |
| Tigecycline | 7 (13.72) | 31 (60.78) | 13 (25.49) |
| Chloramphenicol | 6 (11.76) | 2 (3.92) | 43 (84.31) |
| Cell wall synthesis | |||
| Ampicillin | 37 (72.54) | 12 (23.52) | 2 (3.92) |
| Penicillin | 51 (100) | 0 (0) | 0 (0) |
| Methicillin | 51 (100) | 0 (0) | 0 (0) |
| Vancomycin | 51 (100) | 0 (0) | 0 (0) |
| Imipenem | 0 (0) | 0 (0) | 51 (100) |
| Meropenem | 0 (0) | 0 (0) | 51 (100) |
| Nucleic acid targeting | |||
| Novobiocin | 49 (96.07) | 2 (3.92) | 0 (0) |
| Inhibit DNA synthesis | |||
| Ciprofloxacin | 17 (33.33) | 17 (33.33) | 17 (33.33) |
| Nalidixic acid | 1 (1.96) | 0 (0) | 50 (98.03) |
Table 5.
Number of STEC isolates resistant to different classes of antibiotics.
| Different Classes of Antibiotics | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
|---|---|---|---|---|---|---|---|---|
| Isolates | N | 2 | 1 | 10 | 11 | 10 | 9 | 7 |
| % | 3.92 | 1.96 | 19.6 | 21.5 | 19.6 | 17.6 | 13.7 | |
Table 6.
Frequency of virulence factors among antibiotic-resistant STEC isolates from diverse sources.
Table 6.
Frequency of virulence factors among antibiotic-resistant STEC isolates from diverse sources.
| Antibiotics (n) | Virulence Factors, n (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| stx1 | stx2 | eae | ehxA | nleA | iha | saa | ureC | subA | |
| Gentamicin (50) | 38 (76.00) | 43 (86.00) | 49 (98.00) | 49 (98.00) | 49 (98.00) | 49 (98.00) | 49 (98.00) | 49 (98.00) | 5 (10.00) |
| Kanamycin (39) | 31 (79.48) | 33 (84.61) | 39 (100) | 39 (100) | 39 (100) | 38 (97.43) | 38 (97.43) | 37 (94.87) | 5 (12.82) |
| Streptomycin (42) | 33 (78.57) | 35 (83.33) | 42 (100) | 42 (100) | 42 (100) | 40 (95.23) | 41 (97.61) | 39 (92.85) | 5 (11.90) |
| Erythromycin (51) | 39 (76.47) | 44 (86.27) | 51 (100) | 50 (98.03) | 51 (100) | 49 (96.07) | 48 (94.11) | 48 (94.11) | 5 (9.80) |
| Tetracycline (22) | 18 (81.81) | 17 (77.27) | 22 (100) | 22 (100) | 22 (100) | 22 (100) | 22 (100) | 22 (100) | 3 (1.36) |
| Clindamycin (17) | 15 (88.23) | 15 (88.23) | 17 (100) | 17 (100) | 17 (100) | 17 (100) | 16 (94.11) | 17 (100) | 2 (11.76) |
| Tigecycline (7) | 6 (85.71) | 6 (85.71) | 7 (100) | 7 (100) | 7 (100) | 7 (100) | 7 (100) | 7 (100) | 1 (14.28) |
| Ampicillin (37) | 27 (72.97) | 34 (91.89) | 37 (100) | 36 (97.29) | 37 (100) | 36 (97.29) | 34 (91.89) | 35 (94.59) | 2 (5.40) |
| Penicillin (51) | 39 (76.47) | 44 (86.27) | 51 (100) | 50 (98.03) | 51 (100) | 49 (96.07) | 48 (94.11) | 48 (94.11) | 5 (9.80) |
| Methicillin (51) | 39 (76.47) | 44 (86.27) | 51 (100) | 50 (98.03) | 51 (100) | 49 (96.07) | 48 (94.11) | 48 (94.11) | 5 (9.80) |
| Vancomycin (51) | 39 (76.47) | 44 (86.27) | 51 (100) | 50 (98.03) | 51 (100) | 49 (96.07) | 48 (94.11) | 48 (94.11) | 5 (9.80) |
| Novobiocin (49) | 38 (77.55) | 43 (87.75) | 49 (100) | 48 (97.95) | 49 (100) | 47 (95.91) | 46 (93.87) | 46 (93.87) | 5 (10.20) |
| Chloramphenicol (6) | 5 (83.33) | 6 (100) | 6 (100) | 6 (100) | 6 (100) | 6 (100) | 6 (100) | 6 (100) | 1 (16.66) |
| Ciprofloxacin (17) | 13 (76.47) | 14 (82.35) | 17 (100) | 16 (94.11) | 17 (100) | 16 (94.11) | 15 (88.23) | 16 (94.11) | 2 (11.76) |
| Nalidixic acid (1) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 0 (0) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rubab, M.; Oh, D.-H. Virulence Characteristics and Antibiotic Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Diverse Sources. Antibiotics 2020, 9, 587. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090587
AMA Style
Rubab M, Oh D-H. Virulence Characteristics and Antibiotic Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Diverse Sources. Antibiotics. 2020; 9(9):587. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090587
Chicago/Turabian StyleRubab, Momna, and Deog-Hwan Oh. 2020. "Virulence Characteristics and Antibiotic Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Diverse Sources" Antibiotics 9, no. 9: 587. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090587
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.