Evaluation of Multiple Impacts of Furfural Acetone on Nematodes In Vitro and Control Efficiency against Root-Knot Nematodes in Pots and Fields


Abstract
:1. Introduction
2. Results
2.1. Effect of FAc on the Egg Hatching of M. incognita
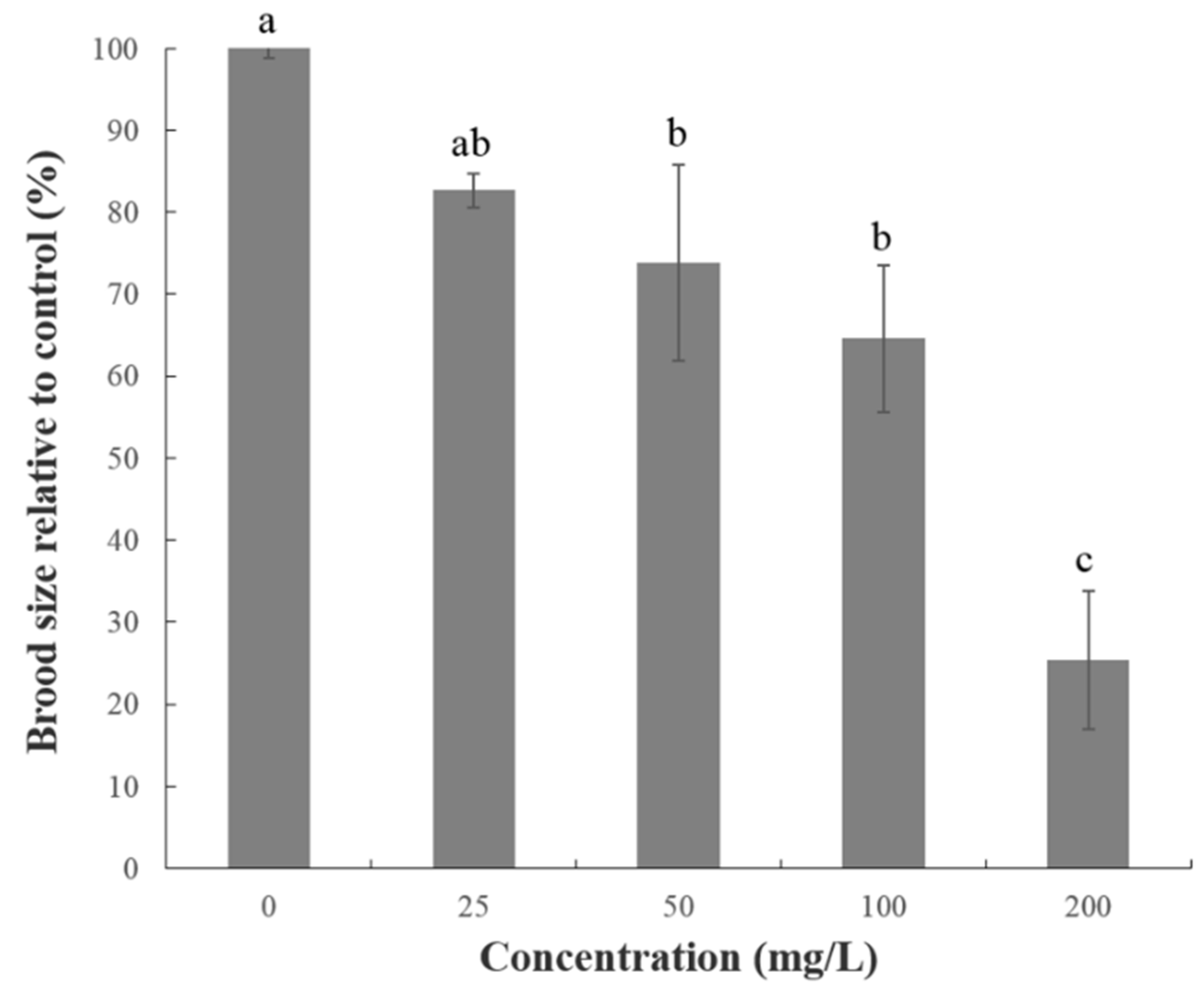
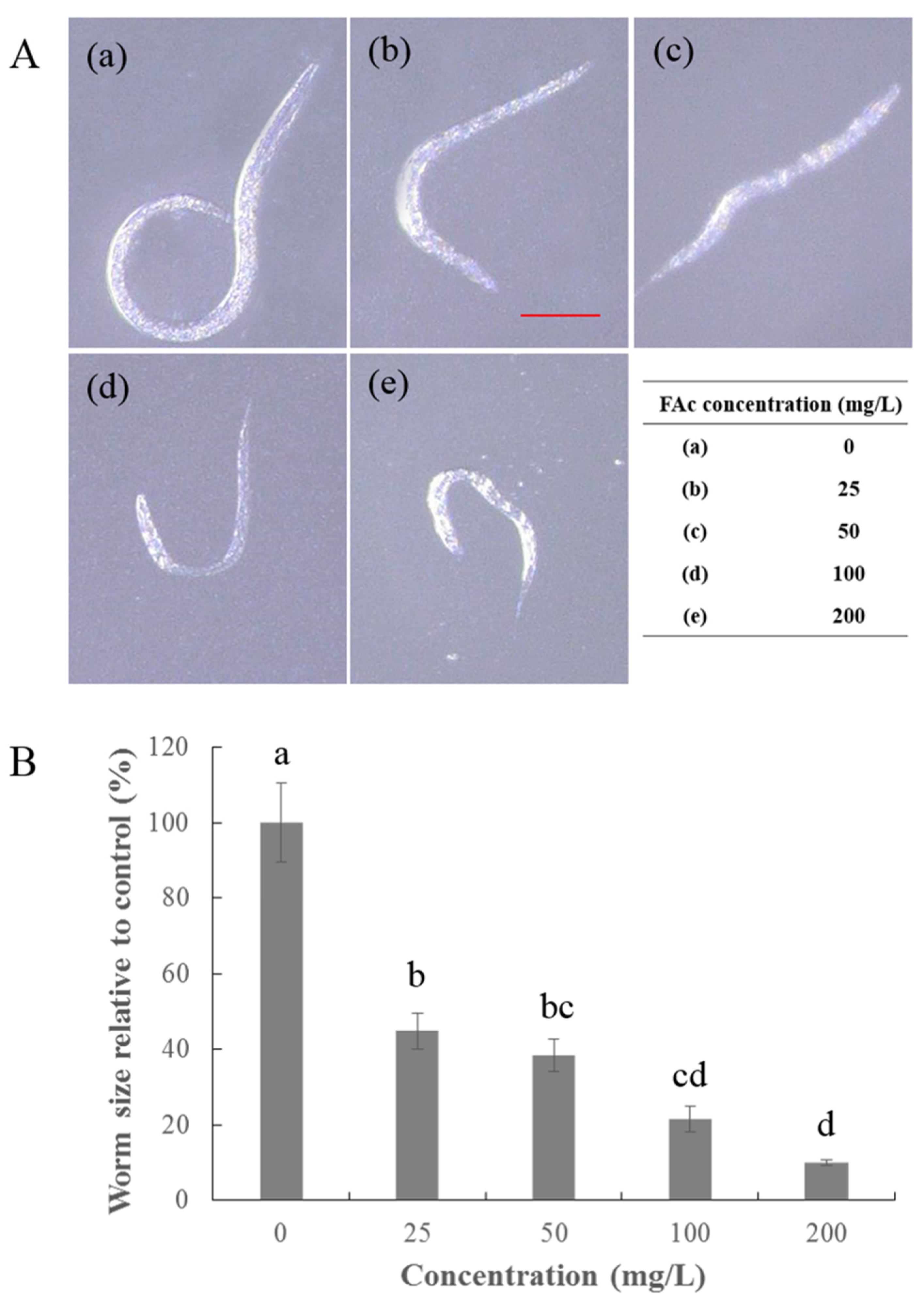
2.2. Effect of FAc on the Reproduction, Feeding, and Growth of C. elegans
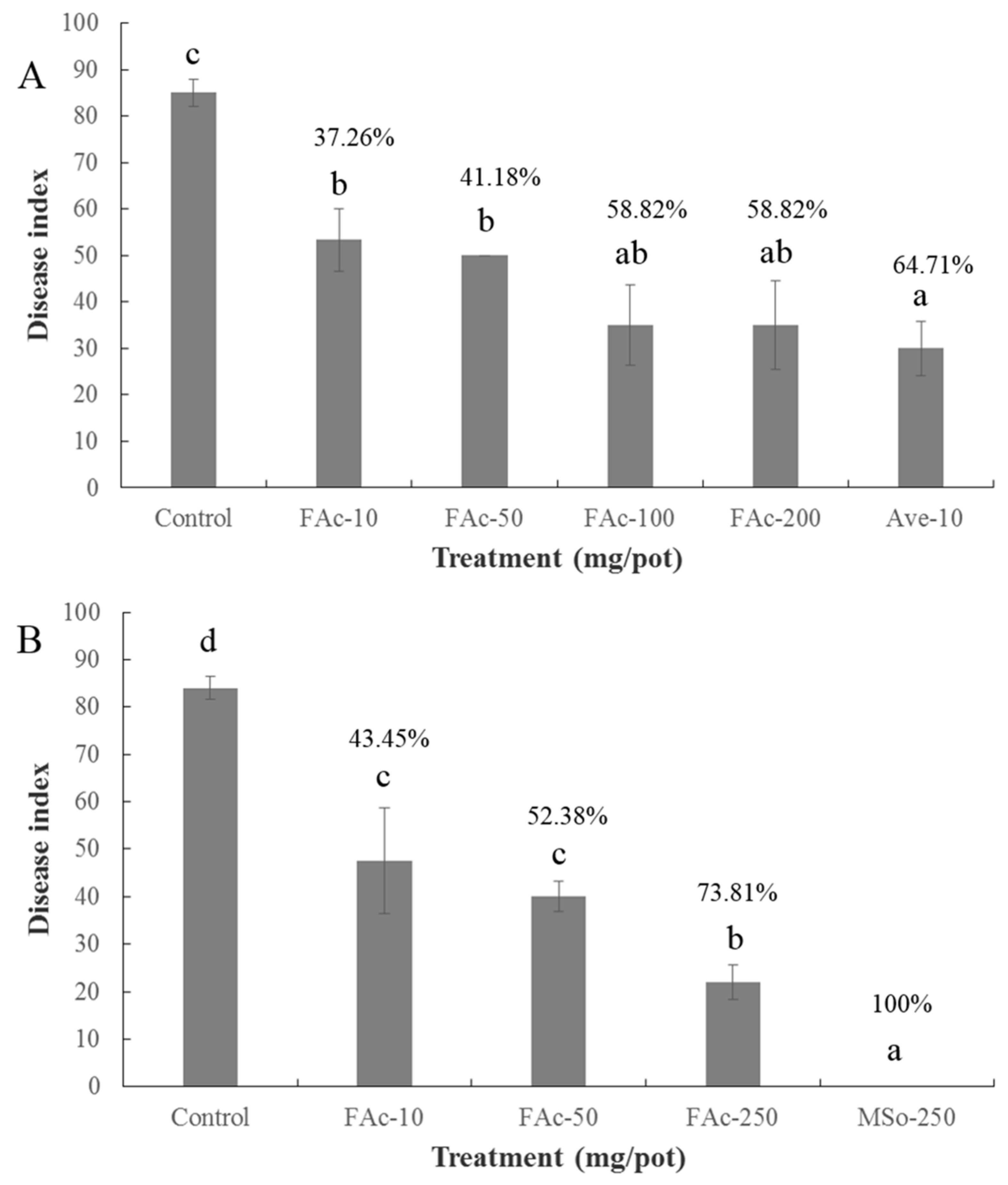
2.3. Efficiency of FAc against RKNs in Pots
2.4. Efficiency of FAc against RKNs in the Field
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Nematodes
4.3. Effect of FAc on the Egg Hatching of M. incognita
4.4. Effect of FAc on the Reproduction of C. elegans
4.5. Effect of FAc on Pharyngeal Pumping of C. elegans
4.6. Effect of FAc on the Growth of C. elegans
4.7. Control Efficacy of FAc against RKNs in Pots
4.8. Control Efficacy of FAc against RKNs in Field
4.9. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Wei, L.H.; Shao, Y.; Wan, J.W.; Feng, H.; Zhu, H.; Huang, H.W.; Zhou, Y.J. Isolation and characterization of a rhizobacterial antagonist of root-knot nematodes. PLoS ONE 2014, 9, e85988. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.W.; Gan, Y.T.; Ji, W.H.; Xu, B.L.; Hou, B.H.; Liu, J. Mechanisms and characterization of Trichoderma longibrachiatum T6 in suppressing nematodes (Heterodera avenae) in wheat. Front. Plant Sci. 2017, 8, 1491. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zou, C.G.; Xu, J.P.; Ji, X.L.; Niu, X.M.; Yang, J.K.; Huang, X.W.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Jang, J.Y.; Jeon, S.J.; Lee, H.W.; Bae, C.H.; Yeo, J.H.; Lee, H.B.; Kim, I.S.; Park, H.W.; Kim, J.C. Nematicidal activity of kojic acid produced by Aspergillus oryzae against Meloidogyne incognita. J. Microbiol. Biotechnol. 2016, 26, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Abdelnabby, H.; Wang, Y.H.; Xiao, X.Q.; Wang, G.F.; Yang, F.; Xiao, Y.N. Impact of direct and indirect application of rising furfural concentrations on viability, infectivity and reproduction of the root knot nematode, Meloidogyne incognita in Pisum sativum. Microb. Pathog. 2016, 96, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Win, P.P.; Kyi, P.P.; Maung, Z.T.Z.; Myint, Y.Y.; Cabasan, M.T.N.; De Waele, D. Crop rotation sequencing to minimize yield losses of summer-irrigated lowland rice in Myanmar caused by the rice root-knot nematode Meloidogyne graminicola. Int. J. Pest Manag. 2020, 66, 319–331. [Google Scholar] [CrossRef]
- Xie, G.H.; Cui, H.D.; Dong, Y.; Wang, X.Q.; Li, X.F.; Deng, R.K.; Wang, Y.; Xie, Y. Crop rotation and intercropping with marigold are effective for root-knot nematode (Meloidogyne sp.) control in angelica (Angelica sinensis) cultivation. Can. J. Plant Sci. 2017, 97, 26–31. [Google Scholar] [CrossRef]
- Jordan, S. Yield to the resistance: The impact of nematode resistant varieties on alfalfa yield. Nat. Resour. Model. 2018, 31, e12150. [Google Scholar] [CrossRef] [Green Version]
- Giannakou, I.O.; Anastasiadis, I. Evaluation of chemical strategies as alternatives to methyl bromide for the control of root-knot nematodes in greenhouse cultivated crops. Crop Prot. 2005, 24, 499–506. [Google Scholar] [CrossRef]
- Desaeger, J.A.; Watson, T.T. Evaluation of new chemical and biological nematicides for managing Meloidogyne javanica in tomato production and associated double-crops in Florida. Pest Manag. Sci. 2019, 75, 3363–3370. [Google Scholar] [CrossRef]
- Coyne, D.L.; Cortada, L.; Dalzell, J.J.; Claudius-cole, A.O.; Haukeland, S.; Luambano, N.; Talwana, H. Plant-parasitic nematodes and food security in Sub-Saharan Africa. Annu. Rev. Phytopathol. 2018, 56, 381–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.J.; Gao, Y.; Li, X.; Chen, S.L.; Yan, S.Z.; Tian, X.J. Identification and nematicidal characterization of proteases secreted by endophytic bacteria Bacillus cereus BCM2. Phytopathology 2020, 110, 336–344. [Google Scholar] [CrossRef]
- Li, L.L.; Tan, J.J.; Chen, F.M.; Hao, D.J. Colonization of Bacillus cereus NJSZ-13, a species with nematicidal activity in Masson pine (Pinus massoniana Lamb.). J. For. Res. 2020, 3, 1025–1033. [Google Scholar] [CrossRef]
- Park, E.J.; Jang, H.J.; Park, C.S.; Lee, S.J.; Lee, S.; Kim, K.H.; Yun, B.S.; Lee, S.W.; Rho, M.C. Evaluation of nematicidal activity of Streptomyces yatensis KRA-28 against Meloidogyne incognita. J. Microbiol. Biotechnol. 2020, 5, 700–707. [Google Scholar] [CrossRef]
- Jayasinghe, U.L.B.; Kumarihamy, B.M.M.; Bandara, A.G.D.; Vasquez, E.A.; Kraus, W. Nematicidal activity of some Sri Lankan plants. Nat. Prod. Res. 2003, 4, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.F.; Gonzalez-Coloma, A.; Munoz, R.; De la Pena, F.; Julio, L.F.; Burillo, J. Nematicidal potential of hydrolates from the semi industrial vapor-pressure extraction of Spanish aromatic plants. Environ. Sci. Pollut. Res. 2018, 25, 29834–29840. [Google Scholar] [CrossRef]
- Zhai, Y.L.; Shao, Z.Z.; Cai, M.M.; Zheng, L.Y.; Li, G.Y.; Yu, Z.N.; Zhang, J.B. Cyclo (l-Pro–l-Leu) of Pseudomonas putida MCCC 1A00316 isolated from Antarctic soil: Identification and characterization of activity against Meloidogyne incognita. Molecules 2019, 24, 768. [Google Scholar] [CrossRef] [Green Version]
- Ntalli, N.G.; Vargiu, S.; Menkissoglu-Spiroudi, U.; Caboni, P. Nematicidal carboxylic acids and aldehydes from Melia azedarach fruits. J. Agric. Food Chem. 2010, 58, 11390–11394. [Google Scholar] [CrossRef]
- Zhai, Y.L.; Shao, Z.Z.; Cai, M.M.; Zheng, L.Y.; Li, G.Y.; Huang, D.; Cheng, W.L.; Thomashow, L.S.; Weller, D.M.; Yu, Z.N.; et al. Multiple modes of nematode control by volatiles of Pseudomonas putida 1A00316 from Antarctic soil against Meloidogyne incognita. Front. Microbiol. 2018, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Yu, C.; Shao, Z.Z.; Cai, M.M.; Li, G.Y.; Zheng, L.Y.; Yu, Z.N.; Zhang, J.B. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [Green Version]
- Fourie, H.; Van Aardt, W.J.; Venter, C.; Tiedt, L.R. The effects of CropGuard (R) on the motility, ultrastructure, and respiration of two Meloidogyne spacies. Nematropica 2014, 44, 93–100. [Google Scholar]
- Rodriguez-Kabana, R.; Kloepper, J.W.; Weaver, C.F.; Robertson, D.G. Control of Plant-parasitic nematodes with furfural-A naturally occurring fumigant. Nematropica 1993, 23, 63–73. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives. Evaluation of certain food additives: Eighty-sixth report of the joint FAO/WHO expert committee on food additives. WHO Tech. Rep. Ser. 2019, 1014, 1–170. [Google Scholar]
- Cheng, W.L.; Yang, J.Y.; Nie, Q.Y.; Huang, D.; Yu, C.; Zheng, L.Y.; Cai, M.M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.N.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Recillas, A.; Arroyo-Herrera, A.L.; Araujo-Leon, J.A.; Nunez, E.; Andrade, R.O. Spasmolytic and antibacterial activity of two Citrus sinensis Osbeck varieties cultivated in Mexico. Evid. Based Complement. Altern. Med. 2017, 2017, 3960837. [Google Scholar] [CrossRef] [PubMed]
- Kurz, C.L.; Ewbank, J.J. Caenorhabditis elegans for the study of host-pathogen interactions. Trends Microbiol. 2020, 8, 142–144. [Google Scholar] [CrossRef]
- Costa, J.C.; Lilley, C.J.; Urwin, P.E. Caenorhabditis elegans as a model for plant-parasitic nematodes. Nematology 2007, 9, 3–16. [Google Scholar]
- Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C. Caenorhabditis elegans as a model organism to evaluate the antioxidant effects of phytochemicals. Molecules 2020, 25, 3194. [Google Scholar]
- Huang, W.K.; Sun, J.H.; Cui, J.K.; Wang, G.F.; Kong, L.A.; Peng, H.; Chen, S.L.; Peng, D.L. Efficacy evaluation of fungus Syncephalastrum racemosum and nematicide avermectin against the root-knot nematode meloidogyne incognita on cucumber. PLoS ONE 2014, 2, e89717. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, C.M.; McGiffen, M.E.; Ohr, H.D.; Sims, J.J.; Becker, J.O. Evaluation of methyl iodide as a soil fumigant for root-knot nematode control in carrot production. Plant Dis. 1999, 1, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.G.; Jiang, H.Y.; Zhang, L.; Zhang, Y.N.; Sial, M.U.; Yu, H.T.; Cao, A.C. Replacing methyl bromide with a combination of 1,3-dichloropropene and metam sodium for cucumber production in China. PLoS ONE 2017, 11, e0188137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.; Umeda, M.; Ishizu, S.; Sato, M. Cytotoxicity of methyl bromide: Effect of methyl bromide on cultured mammalian cells. J. Toxicol. Sci. 1980, 4, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.J.; Zhang, Z.W.; Wang, X.S.; Xu, S.W.; Li, M.; Li, S. Effects of avermectin on microsomal cytochrome P450 enzymes in the liver and kidneys of pigeons. Environ. Toxicol. Pharmacol. 2014, 2, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Gonzalez, M.; Solis-Fuentes, J.A.; Duran-Dominguez-de-Bazua, M.D. Metam Sodium (MS) in water and cane juice at different processing conditions according to the industrial case. part 1. effects of matrix, pH, temperature, processing time, and photolysis. Water Air Soil Pollut. 2017, 12, 456. [Google Scholar] [CrossRef]
- Zhu, J.W.; Dhammi, A.; van Kretschmar, J.B.; Vargo, E.L.; Apperson, C.S.; Roe, R.M. Novel use of aliphatic n-methyl ketones as a fumigant and alternative to methyl bromide for insect control. Pest Manag. Sci. 2018, 3, 648–657. [Google Scholar] [CrossRef]
- Fakhfakh, N.; Cognet, P.; Cabassud, M.; Lucchese, Y.; Rios, M.D.D.L. Stoichio-kinetic modeling and optimization of chemical synthesis: Application to the aldolic condensation of furfural on acetone. Chem. Eng. Process. 2008, 47, 349–362. [Google Scholar] [CrossRef] [Green Version]
- Wierckx, N.; Koopman, F.; Ruijssenaars, H.J.; de Winde, J.H. Microbial degradation of furanic compounds: Biochemistry, genetics, and impact. Appl. Microbiol. Biot. 2011, 6, 1095–1105. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, L.A.; Campos, V.P.; Pedroso, M.P.; Barros, A.F.; Freire, E.S.; Resende, F.M.P. Volatile organic compounds produced by castor bean cake incorporated into the soil exhibit toxic activity against Meloidogyne incognita. Pest Manag. Sci. 2019, 2, 476–483. [Google Scholar] [CrossRef]
- Bui, H.X.; Hadi, B.A.R.; Oliva, R.; Schroeder, N.E. Beneficial bacterial volatile compounds for the control of root-knot nematode and bacterial leaf blight on rice. Crop Prot. 2020, 135, 104792. [Google Scholar] [CrossRef]
- Raza, W.; Wang, J.N.; Jousset, A.; Friman, V.P.; Mei, X.L.; Wang, S.M.; Wei, Z.; Shen, Q.R. Bacterial community richness shifts the balance between volatile organic compound-mediated microbe–pathogen and microbe–plant interactions. Proc. R. Soc. B Biol. Sci. 2020, 1925, 20200403. [Google Scholar] [CrossRef] [Green Version]
- Manan, A.; Bazai, Z.A.; Fan, J.; Yu, H.F.; Li, L. The Nif3-family protein YqfO03 from Pseudomonas syringae MB03 has multiple nematicidal activities against Caenorhabditis elegans and Meloidogyne incognita. Int. J. Mol. Sci. 2018, 19, 3915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudelska, M.M.; Lewis, A.; Ng, C.T.; Doyle, D.A.; Holden-Dye, L.; O’Connor, V.M.; Walker, R.J. Investigation of feeding behaviour in C. elegans reveals distinct pharmacological and antibacterial effects of nicotine. Invertebr. Neurosci. 2018, 18, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridge, J.; Page, S.L.J. Estimation of root-knot nematode infestation levels on roots using a rating chart. Int. J. Pest Manag. 1980, 26, 296–298. [Google Scholar] [CrossRef]
- Zhou, L.H.; Yuen, G.; Wang, Y.; Wei, L.F.; Ji, G.H. Evaluation of bacterial biological control agents for control of root-knot nematode disease on tomato. Crop Prot. 2016, 84, 8–13. [Google Scholar] [CrossRef]
- Cheng, X.K.; Liu, X.M.; Wang, H.Y.; Ji, X.X.; Wang, K.Y.; Wei, M.; Qiao, K. Effect of emamectin benzoate on root-knot nematodes and tomato yield. PLoS ONE 2015, 10, e0141235. [Google Scholar] [CrossRef]
- Freckman, D.H.; Virginia, R.A. Extraction of nematodes from dry valley antarctic soils. Polar Biol. 1993, 13, 483–487. [Google Scholar] [CrossRef]
- Seesao, Y.; Gay, M.; Merlin, S.; Viscogliosi, E.; Aliouat-Denis, C.M.; Audebert, C. A review of methods for nematode identification. J. Microbiol. Methods 2017, 138, 37–49. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/L) | 2 Days | 4 Days | ||
|---|---|---|---|---|
| Hatched Worms/Egg Mass | Inhibition Rate (%) | Hatched Worms/Egg Mass | Inhibition Rate (%) | |
| 0 | 47.6 ± 5.8 a 1 | (0.0) | 74.3 ± 9.3 a | (0.0) |
| 50 | 40.1 ± 4.9 a | 15.8 | 67.8 ± 10.1 a | 8.8 |
| 100 | 16.9 ± 3.3 b | 64.6 | 19.7 ± 5.3 b | 73.5 |
| 200 | 4.0 ± 1.5 c | 91.7 | 4.6 ± 1.5 b | 93.8 |
| 500 | 2.4 ± 0.6 c | 94.9 | 2.7 ± 0.7 b | 96.4 |
| Concentrations (mg/L) | Pumping 1 (Hz) | |
|---|---|---|
| 24 h | 48 h | |
| 0 | 183.7 ± 2.6 a 2 | 175.1 ± 3.9 a |
| 25 | 163.7 ± 8.8 ab | 155.4 ± 2.7 b |
| 50 | 164.3 ± 7.8 ab | 147.9 ± 4.5 b |
| 100 | 148.1 ± 7.8 b | 135.5 ± 4.5 c |
| 200 | 117.8 ± 2.9 c | 109.1 ± 3.5 d |
| Treatment | Disease Index | Control Effect (%) | Number of RKNs/30 g Soil | Reduction Rate of RKNs (%) |
|---|---|---|---|---|
| Irrigation 1 | ||||
| Control 2 | 82.4 ± 2.2 b 7 | (0.0) | 432 ± 93 a | (0.0) |
| Fac 3 10 mg/plant | 57.5 ± 2.8 a | 30.2 | 246 ± 62 a | 43.1 |
| FAc 50 mg/plant | 52.3 ± 2.9 a | 36.9 | 225 ± 97 a | 47.9 |
| Ave 4 10 mg/plant | 58.1 ± 3.0 a | 29.5 | 401 ± 31 a | 7.2 |
| Fumigant 5 | ||||
| Control | 73.6 ± 2.6 b | (0.0) | 1617 ± 1323 a | (0.0) |
| FAc 375 mg/m2 | 61.1 ± 3.8 a | 17.1 | 759 ± 259 a | 53.1 |
| FAc 750 mg/m2 | 59.0 ± 2.6 a | 19.9 | 400 ± 51 a | 75.3 |
| MSo 6 750 mg/m2 | 65.4 ± 3.2 a | 11.2 | 726 ± 314 a | 55.1 |
| Treatment | Fresh Weight on the Ground (g) | Plant Height (cm) | Stem Thickness (cm) |
|---|---|---|---|
| Irrigation | |||
| Control | 618 ± 78 a 1 | 89.5 ± 3.1 a | 3.64 ± 0.13 a |
| FAc 10 mg/plant | 737 ± 109 a | 96.5 ± 3.7 a | 3.76 ± 0.11 a |
| FAc 50 mg/plant | 593 ± 72 a | 85.3 ± 4.4 a | 3.59 ± 0.14 a |
| Ave 10 mg/plant | 655 ± 16 a | 94.7 ± 2.8 a | 3.72 ± 0.10 a |
| Fumigant | |||
| Control | 667 ± 52 a | 101.0 ± 3.0 a | 3.46 ± 0.09 a |
| FAc 375 mg/m2 | 724 ± 77 a | 98.7 ± 3.4 a | 3.70 ± 0.15 a |
| FAc 750 mg/m2 | 749 ± 44 a | 105.4 ± 2.9 a | 3.71 ± 0.11 a |
| MSo 750 mg/m2 | 821 ± 42 a | 106.1 ± 2.7 a | 3.63 ± 0.08 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, W.; Yang, X.; Zeng, L.; Huang, D.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Evaluation of Multiple Impacts of Furfural Acetone on Nematodes In Vitro and Control Efficiency against Root-Knot Nematodes in Pots and Fields. Antibiotics 2020, 9, 605. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090605
Cheng W, Yang X, Zeng L, Huang D, Cai M, Zheng L, Yu Z, Zhang J. Evaluation of Multiple Impacts of Furfural Acetone on Nematodes In Vitro and Control Efficiency against Root-Knot Nematodes in Pots and Fields. Antibiotics. 2020; 9(9):605. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090605
Chicago/Turabian StyleCheng, Wanli, Xue Yang, Li Zeng, Dian Huang, Minmin Cai, Longyu Zheng, Ziniu Yu, and Jibin Zhang. 2020. "Evaluation of Multiple Impacts of Furfural Acetone on Nematodes In Vitro and Control Efficiency against Root-Knot Nematodes in Pots and Fields" Antibiotics 9, no. 9: 605. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090605