Novel Approaches to Combat Medical Device-Associated BioFilms



Department of Infectious Diseases, First Hospital of Jilin University, Changchun 130021, China
*
Author to whom correspondence should be addressed.
Coatings 2021, 11(3), 294; https://0-doi-org.brum.beds.ac.uk/10.3390/coatings11030294
Submission received: 24 January 2021
/
Revised: 25 February 2021
/
Accepted: 27 February 2021
/
Published: 4 March 2021
(This article belongs to the Special Issue Novel Approaches to Combat Biofilms Formed on Medical Devices)
Abstract
:Biofilms are aggregates formed as a protective survival state by microorganisms to adapt to the environment and can be resistant to antimicrobial agents and host immune responses due to chemical or physical diffusion barriers, modified nutrient environments, suppression of the growth rate within biofilms, and the genetic adaptation of cells within biofilms. With the widespread use of medical devices, medical device-associated biofilms continue to pose a serious threat to human health, and these biofilms have become the most important source of nosocomial infections. However, traditional antimicrobial agents cannot completely eliminate medical device-associated biofilms. New strategies for the treatment of these biofilms and targeting biofilm infections are urgently required. Several novel approaches have been developed and identified as effective and promising treatments. In this review, we briefly summarize the challenges associated with the treatment of medical device-associated biofilm infections and highlight the latest promising approaches aimed at preventing or eradicating these biofilms.
1. Introduction
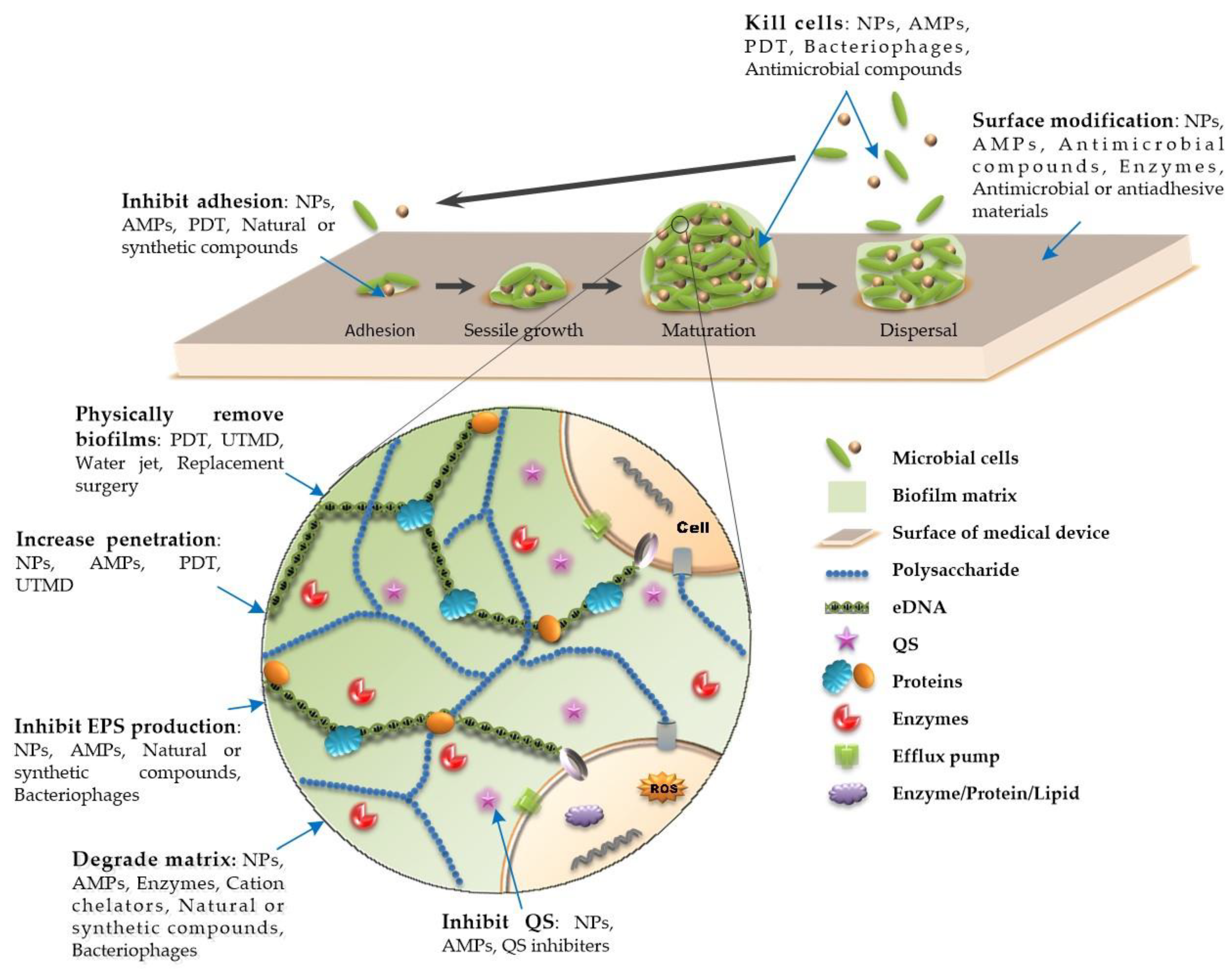
A biofilm is a special existence state with a three-dimensional tower structure formed by a microbial community attached to a biological or nonbiological surface and embedded in the matrix of an extracellular polymeric substance (EPS). An EPS is secreted by attached cells and is mainly composed of polysaccharides, extracellular DNA (eDNA), proteins, and other secreted substances (Figure 1) [1,2,3]. These substances act as a shield that often makes microorganisms resistant to physical removal, host immune responses, and antimicrobial treatments [1,2,3]. The development of biofilms is linked to quorum sensing
(QS), which is a cell-to-cell communication mechanism that regulates a variety of physiological and biochemical functions [4,5]. It can influence the biological cycle of biofilms by targeting matrix compounds and regulate the production of multiple virulence factors to resist phagocytosis [4,6,7]. Biofilms have their own properties that change dynamically, and their biological cycle involves four stages, namely, adhesion, sessile growth, maturation, and dispersal [1,8]. The adhesion and sessile growth stages are reversible, wherein the cells aggregate loosely, but can also become detached and return to a free-floating state [3,8,9]. The maturation of biofilms is related to the production of EPS, which encapsulates cells in a thick and stable layer of complex biomolecules. This stage is irreversible [3,8,10,11]. Cells in the dispersion stage can secrete destruction factors to promote detachment of the biofilm and the colonization of surrounding sites [3,8,10,12]. Biofilms exhibit spatial heterogeneity [10,13]. The concentrations of various chemical substances in the biofilm, such as metabolic products, signal molecules, and nutrients, change stepwise from the surface to the inside, which makes the physiological activities of bacteria different in different parts of the biofilm [10,13]. The outer-layer cells remain metabolically active, while the inner-layer cells are persister bacteria, which are usually dormant [14]. Because of the uneven metabolism of bacteria in biofilms, regardless of the metabolic link that the antibacterial factors act on, some bacteria that are not in this metabolic state can survive, leading to difficulties in the elimination of infection or repeated outbreaks of infection.
Up to 80% of microbial infections in the human body involve biofilm formation, especially in hospital settings, which greatly promotes the incidence rate and mortality [15,16]. With the high standards for the quality of life of patients, the improvement of medical standards, and the widespread application of medical devices, medical device-associated biofilms pose a serious threat to human health, and these biofilms are the most important source of nosocomial infections, despite the comprehensive prevention and control measures taken, such as disinfection and hygiene monitoring. According to statistics, about 2 million nosocomial infection cases occurred in the United States each year at the beginning of the 21st century, of which 50%–70% of nosocomial infections were related to indwelling medical devices [17,18,19]. Given the challenges associated with medical device-associated biofilm control, new strategies are urgently needed for the treatment of these biofilm-specific infections. Several novel approaches have been developed and identified as effective and promising treatment strategies. Although previous reviews have already summarized novel approaches to combat medical device-associated biofilms [20,21,22,23], we briefly summarize the challenges associated with the treatment of medical device-associated biofilm infections and provide a more comprehensive summary of strategies that can be employed to combat medical device-associated biofilms. In particular, we discuss the progress and limitations of the latest studies on antimicrobial peptides (AMPs) and nanotechnology in detail. Moreover, bacteriophage therapy is included in our review, though it is often ignored by most other review articles.
2. Medical Device-Related Biofilm Infection and the Challenges of Treatment
Implantable medical devices have become an integral part of treatments in clinical work. However, in the body, external implantation is often accompanied by infection complications. Approximately 60%–70% of nosocomial infections are associated with biomaterials or implants [24]. Medical device-related biofilm infections are mainly caused by the colonization of microorganisms on abiotic surfaces, such as indwelling medical devices. All implantable medical devices (e.g., cardiac pacemakers, artificial heart valves, hemodialyzers, artificial joints, needleless connectors, center venous catheters, urinary catheters, endotracheal intubation, artificial breasts, contact lenses, and orthodontal prosthetics) carry the risk of biofilm-related infections, which severely affect the quality of life and even endanger life. We have tried to summarize the rates of infection for some of the most common medical devices (Table 1). Catheter-related urinary tract infections are very common, and microorganisms in biofilms on the inner surface of the catheter in patients with long-term catheterization can resist antibiotic treatment and cause chronic infections [25]. Biofilms formed by microorganisms in the endotracheal tube are related to ventilator-associated pneumonia, which is one of the most common infections in the intensive care unit (ICU) and the main cause of death [26]. Long-term medical implants can cause inflammation and tissue destruction around implants, and sometimes, these infections are life threatening [1]. Pathogens contaminate indwelling medical devices through a variety of exogenous pathways. Foreign microorganisms can be transferred to the surface of medical devices, either through cross-infection of the surgical devices, direct inoculation during device placement, or migration of the surrounding skin mucosal sheath [1]. Host-related bacteria can colonize the implanted device after the operation, such as bacteria from the respiratory tract, urinary tract, and gastrointestinal tract [25]. In addition, microorganisms can form a medical device-associated biofilm through a bloodstream infection. Irrespective of whether the microbes that cause infection are present inside or outside the body, medical device-related infections often begin with the colonization of bacteria on the surface of the device and go through a series of complex changes leading to the formation of biofilms. Persistent biofilms are the main cause of chronic wound infection and infection recurrence. In a meta-analysis study, the prevalence of biofilms in chronic wound infections was 78.2% [27]. Biofilms formed on chronic peritoneal dialysis catheters are considered to be an important cause of peritonitis recurrence or persistence [28]. Biofilm-associated infections are often exacerbated by the host’s strong proinflammatory response to persistent microbial stimulation, which may greatly promote tissue damage [10]. Microbial cells may separate from the biofilm and spread to surrounding tissues or blood, thereby further worsening the clinical outcome of the infection [29]. Various pathogens have been implicated in device-related infections. Some pathogens that form biofilms originate from the skin and contaminate the device during implantation or insertion. Staphylococcus epidermidis, Staphylococcus aureus, and Candida albicans are common microbes on the skin; therefore, they are the most important pathogenic cause of implant biofilm infections [1,30,31,32]. However, the pathogens that form biofilms are different at different sites of infection and are related to many factors. For example, the most common pathogen in biofilms formed on indwelling catheters is Escherichia coli, and the sex, catheterization period, and catheter type have also been shown to be significantly correlated with catheter biofilms [33]. Several microorganisms of the normal flora cause infection when the immune system is compromised, including E. coli [34], Pseudomonas aeruginosa [35], Klebsiella pneumonia [36], Acinetobacter baumannii [37], and Enterococcus faecalis [38]. We have listed the main microorganisms involved in the formation of biofilms at the level of various medical implants in Table 1. Biofilms can be formed by a single bacterial strain. However, biofilms are usually formed by a variety of microorganisms in nature, and such mixed-species biofilms represent the actual lifestyles of microbes, including bacteria, fungi, and/or protozoa [39]. Bacterial species extensively interact with each other and these interactions (such as adhesin-mediated coaggregation, bacterial surface pili, and flagella, and their mediated motilities, competition, and cooperation) determine the structure development and functional dynamics of multi-species biofilms [40]. The oral biofilm formation process is initiated by an early colonizer, such as S. gordonii, S. sanguinis, and S. oligofermentans, which provides further attachment sites for later colonizers, such as S. mutans. The succession of biofilm development involves the coaggregation and coadherence of oral bacteria, and if undisturbed, develops into a stratified, complex biofilm [41].
The resistance of bacteria in biofilms to antibiotics can be 10–1000 times that of the corresponding planktonic cells [88]. The mechanisms of biofilm resistance to conditional antimicrobial agents have been studied carefully and can be summarized as follows. The presence of biofilms enhances the drug resistance of bacteria, which is different from the mechanism of drug resistance of planktonic cells. As mentioned above, the large amounts of EPSs, such as exopolysaccharides and eDNA, in biofilms form molecular and charge barriers to prevent and delay drug penetration [89,90]. Extracellular DNA can also induce the expression of drug resistance genes by chelating cations to increase the drug resistance [91]. In addition, some high-concentration hydrolases are present in biofilms; these hydrolases inactivate antibacterial drugs that enter the biofilm. The biofilm matrix provides shelter and physical and chemical barriers for cells (Figure 1). Due to the heterogeneity of biofilms, such as those in cell metabolism, nutrients, and oxygen, bacterial cells in low-metabolism and slow-growth states are not sensitive to most antimicrobial agents. Cells in biofilms also acquire a resistance phenotype by altering their DNA, which has been associated with increased oxidative stress in biofilms [92,93], leading to, for example, increased β-lactamase expression, the activation of multidrug efflux pumps, or the downregulation of outer membrane proteins [94,95,96,97,98]. Biofilms are considered ideal reaction chambers for horizontal gene transfer. Cells can acquire antibiotic resistance through the horizontal transfer of antibiotic resistance genes due to the high cell density of bacteria in biofilms [99]. Horizontal gene transfer promotes biofilm formation and heightens pathogens’ host direct fitness. Furthermore, it can enforce pathogens to maintain stable social interactions [100]. Cells communicate through signal transduction pathways (e.g., QS or two-component systems), leading to global changes in gene expression by conjugation, transformation, and transduction, enhancing the virulence and expediting the acquisition of antibiotic resistance [101]. Among them, conjugation is certainly the most commonly studied. Victoria J et al. have demonstrated that the biofilm growth of S. aureus dramatically increases the horizontal transfer of plasmid-borne antibiotic resistance determinants by conjugation/mobilization [102]. It has also been reported that the horizontal transfer of antibiotic resistance genes between cells occurs efficiently in early biofilms and multiresistant gene transfer is heavily suppressed in dense biofilms [103]. Researchers have paid more attention to the risk of the development and dissemination of horizontal gene transfer in medical device-related biofilms in recent years and some pathogens used in horizontal gene transfer studies have been directly isolated from biofilms located in implanted medical devices [104]. Many horizontal transfer genes have been identified, such as pSK41 and pT181, which were identified as the most common staphylococcal horizontal transfer and relaxase genes, as well as aac6-aph2a, ermC, and tetK genes, which were the most common antibiotic resistance genes in Staphylococcus aureus and Staphylococcus epidermidis biofilms [104].
Each of these mechanisms can only partially explain the increase in antimicrobial resistance observed in biofilms. However, in the face of the synergistic effects of these mechanisms, even the most aggressive antibiotic treatment options or combination schemes involving different kinds of antibiotics cannot completely eliminate biofilm infections because of the limited dose of antimicrobial agents and their side effects [105]. In addition, the removal of medical devices can effectively prevent the recurrence of biofilm infections. Although many studies have emphasized the importance of implant removal to avoid the recurrence of infection at the surgical site, not all patients need or can undergo surgical revision surgery for local infections [32]. Implant exchange surgery, that is, the replacement of infected implants with new implants, may not be the best choice for the treatment of biofilm infection due to the high degree of trauma and high infection rate [106], which not only cause great physical and mental damage to patients, but also increase the economic burden on patients and the medical and healthcare system [32,106]. A study on the treatment of infection with surgical site implants revealed that the average number of surgeries required for infection treatment was 4 (range 1–16), resulting in an average cost of treatment ranging from two hundred and fifty thousand to one million dollars per patient [107]. Approximately 2 million healthcare-related infections occur in the United States each year, resulting in more than $5 billion in additional medical expenses each year. Among them, 50%–70% of nosocomial infections are related to indwelling medical devices [17,18,19]. Additionally, the mortality from nosocomial infections associated with medical devices is largely dependent on the type of devices, ranging from <5% for dental implants and foley catheters to >25% for mechanical heart valves [108]. In the intensive care unit, the incidence of catheter-related bloodstream infections is as high as 2.5 ~ 6.9/1000 catheter days, depending on the type of catheter and the site of catheter insertion [17,109]. The additional economic cost of treatment for each catheter-related infection may be as high as $30,000, with the potential for increased mechanical ventilation, an additional 1-week stay in the ICU, and an additional 2–3-week hospital stay [110,111].
Therefore, the constant improvement of prevention strategies and the development of novel therapeutic approaches are needed to overcome these weaknesses. Currently, we still need to emphasize the basic role of antimicrobial therapy in combatting medical device-associated biofilms, while the combination of novel promising approaches and antimicrobial therapy may help us eradicate existing biofilms.
3. Novel Approaches to Combat Medical Device-Associated Biofilms
3.1. Physical Therapy
The removal of medical devices from the body and continuous irrigation and debridement are the most successful treatment options for eradicating biofilms when standardized antibiotic treatment fails to effectively eliminate medical device-related biofilm infections [32]. Because of technical difficulties and a high morbidity, the removal of implants infected with biofilms, such as prosthetic joints and pacemakers, may not be the best option.
Photodynamic therapy (PDT) is an effective physical method for removing biofilms. It is composed of three main components: Light; a chemical molecule called a photosensitizer; and oxygen [112]. Light at a specific wavelength has an antibacterial effect. Antibacterial PDT generates highly cytotoxic reactive oxygen species (ROS) or excitable singlet oxygen to oxidize biological molecules on or within the cell membrane, such as proteins, nucleic acids, and lipids, causing cell DNA damage, cell membrane and organelle destruction, and even cell death [113] (Figure 1). A recent study has shown that PDT can not only inhibit the formation of E. faecalis biofilms formed on root canals by producing ROS, but also fight against biofilm infections by inhibiting the expression of toxic factors that affect early bacterial adhesion and promote the formation of the biofilm matrix structure [114] (Figure 1). Additionally, nanomaterials (NMs) can be used in PDT to enhance the effect of PDT by generating heat or transforming NMs into biofilm-destroying species [115,116]. PDT technology is mostly used in research on the treatment of dental plaque-related diseases [117]. PDT technology can be used to combat other implant-related biofilm infections, such as prosthetic joint infections and infections caused by ventilator-associated pneumonia biofilms [118,119]. Although preclinical studies have suggested that PDT is a promising technique for the treatment of biofilm infections, there remain many measures that need to be taken before it can be safely applied in clinical practice. The short-term side effects of PDT are usually mild and self-limiting, such as erythema, edema, and pain in the surrounding skin, while long-term exposure to light may cause cataracts, skin cancer, and other side effects [120]. During PDT, protection measures should be taken to protect the patient’s eyes from laser exposure and to ensure that the photosensitizer and photochemical reactions do not affect the patient’s surrounding tissues [11].
Studies have shown that low-intensity ultrasound at the physiotherapy level can enhance the activity of antibacterial agents, resulting in a powerful biofilm removal effect [121]. Moreover, ultrasound or ultrasound-targeted microbubble destruction (UTMD) can destroy the matrix structure of bacterial biofilms by using cavitation to promote drug entry into the biofilm and can also directly destroy cells and increase the metabolic activity of cells to enhance the bactericidal ability of antibiotics [122,123,124] (Figure 1). These findings provide new insights for the use of ultrasound or UTMD to improve the effectiveness of antibiotic therapy for prosthesis- or catheter-related biofilms, and these approaches have shown promising effects in vivo [124,125]. However, ultrasound-mediated antibiotic activity on bacteria in biofilms is affected by many factors, including the acoustic intensity, duty cycle, frequency, and duration [126]. For example, a greater ultrasound intensity and longer duration may cause skin or tissue damage or cause bacteria to spread to surrounding tissues and affect blood flow [127,128]. Cavitation can also potentially damage the implant [129]. Considering that this technique may be applied to clinical practice in the future, further research is needed to select the optimal ultrasound parameters.
The use of water jets is another effective physical method for removing biofilms on the surface of implants through the mechanical action of pulse and pressure [130]. Water jets can wash away loosely attached plaques, eliminate bacterial cells, and interfere with biofilm maturation [131] (Figure 1). A previous study has shown that under different pressure conditions, the ability of dental water jets to remove biofilms differs. The research results showed that, under pressures of 707, 350, and 102 kPa, the rate of reduction in biofilm thickness was 85.5%, 85.1%, and 63.4%, respectively [132]. A recent study has shown that the combination of PDT and water jets has good application prospects for removing scale from the surface of traditional implants. Compared with an untreated biofilm, the biofilms of Streptococcus mutans grown on a titanium surface treated with PDT were more easily removed by the impact force of a water jet. This showed that PDT can weaken biofilms in different stages, making them easier to separate with a simple washing force [133]. Another study demonstrated that the cavitating jet can also effectively clean biofilms formed on the rough surface of implant screws, especially on the root sector [134].
In addition, mechanical methods, such as the use of a plastic curette, an air-abrasive device (Perio-Flow), a titanium brush (TiBrush), and implantoplasty, and electrochemical treatment can effectively clean dental biofilms and improve hygiene around the implant [135,136]. However, it has been found that the mechanical treatment strategies used to remove implant biofilms may cause titanium dissolution and increase the cytotoxicity. Therefore, the dissolution of implant material should be an important consideration in the clinical choice of treatment of peri-implant inflammation [137].
3.2. Surface Modification
Applying antibacterial agents or antiadhesion agents to the surface of medical devices through surface engineering can effectively inhibit the adhesion and growth of microorganisms and prevent the formation of biofilms, which has become an important strategy for combatting biofilms related to medical devices. Antibacterial coating approaches prevent the formation and development of biofilms on medical devices through bacteriostatic or bactericidal effects. Traditional antibacterial coatings, such as chlorhexidine, gentamicin, minocycline, rifampicin, silver sulfadiazine, amikacin, and vancomycin, have been widely used in clinical practice, showing the efficacy of preventing catheter-related and other implant-related infections [138,139,140]. The combination of multiple antimicrobials, such as the combination of the antibiotics minocycline and rifampicin, has also shown good results in preventing single-drug resistance [138]. On the other hand, the extensive use of antimicrobial coatings may lead to bacterial resistance to antibiotics. To date, studies of antibacterial coatings have focused on the development of novel compound coatings with antibacterial effects or antibiotic delivery functions to prevent the formation of biofilms. We have summarized the novel surface-modifying compounds used against biofilms and mechanisms of action in Table 2.
Antimicrobial compound coatings or antiadhesion coatings are used to modify the surfaces of medical device materials to inhibit biofilm formation by killing bacteria or reducing bacterial adhesion (Figure 1). An isoeugenol coating has been shown to prevent the formation of biofilms on stainless steel and polyethylene surfaces due to its good antibacterial activity [141]. N-acetyl cysteine is an Food and Drug Administration (FDA) -approved drug that is widely used to treat many different clinical diseases, such as chronic bronchitis, cardiac ischemia-reperfusion injury, and paracetamol poisoning [142]. The drug has been suggested to not only have antibacterial activity [143], but also reduce bacterial adhesion and prevent biofilm formation. Because of its excellent safety performance, it has been widely studied as a coating for medical implants, such as bone-related implants [144]. Existing methods of coating biomaterials with antibiotics and other reagents produce surfaces that are prone to degradation over time. Metal-based materials have a long history as antibacterial agents [145], such as the silver coatings used on a variety of medical devices. NMs exhibit enhanced antibacterial activity and a reduced cytotoxicity [146]. Silver nanoparticle (AgNP) coatings are also used to prevent bacteria from becoming attached to medical devices [147]. A recent study has shown that the topographical effects of nanostructure surfaces may have significant impacts on the adherence pattern and stretching degree of the bacterial cell membrane [148]. New alloys and metal–ceramic compounds are also used in anti-medical device biofilms. For example, a copper-bearing titanium alloy has been shown to efficiently kill the attached bacteria by damaging cell membranes and cell walls and strongly inhibit the biofilm formation and prevent bacterial infection associated with dental implants, meaning that it has great potential for application in dental implants with an excellent antibacterial viability and positive effect against bone resorption induced by peri-implantitis [149]. The tantalum processed by micro-Arc oxidation increased its surface roughness, hydrophilicity, and corrosion resistance, and improved its antibacterial activity against S. aureus, meaning that it exhibits great potential for application as a material for hard tissue implants [150]. Polycrystalline Zirconium Dioxide Ceramic presents the same characteristics for S. mutans biofilm formation as polymer-infiltrated ceramic, which can inhibit biofilm formation, and they both have a good biocompatibility with human fibroblasts. Therefore, they have great potential in the application of indirect restoration and implant abutment [151]. Antiadhesion coatings have also been realized by changing the physical and chemical properties of surfaces so that the bacteria no longer adhere easily. Hydrophobin has been used to coat different medically relevant materials and has been shown to be effective in preventing biofilm formation by S. epidermidis on polystyrene surfaces through the formation of a self-assembled amphiphilic layer [152]. The most common denture material, heat-polymerized polymethyl methacrylate, exhibits a high surface roughness, hydrophobicity, and surface free energy, which contribute to the initial adhesion of microorganisms. Cyanoacrylate adhesives can change the characteristics of the resin surface, reduce the surface roughness, increase the hydrophilicity, and change the surface free energy, thus inhibiting the formation of biofilms. Adhesives also have a good biocompatibility, so they are used to coat medical devices [153]. Ultrahigh-molecular-weight polyethylene (UHMWPE) is a kind of solid, inert, and stable polymer and is the most commonly used polymer in total hip and knee arthroplasty. However, its strong hydrophobic surface makes it vulnerable to bacterial adhesion and biofilm formation. Polymer hydrophilic coatings are grafted to the surface of UHMWPE by polymer coating technology, such as covalent chemical bonds, which change the surface chemistry from hydrophobic to hydrophilic and prevent bacterial adhesion and biofilm formation [154]. Many studies on the prevention of biofilm formation have been carried out by changing the surface properties of materials, such as the roughness, hydrophobicity/hydrophilicity, surface free energy, and electric charge, to find more suitable materials that can be implanted into the human body and function favorably. For example, modification of the surface of orthopedic implants, in addition to effective resistance to bacterial adhesion, the functional retention of raw material implants, biocompatibility, the ability to resist tribological corrosion of coatings, and nontoxicity to host cells, should also be considered and evaluated [155]. S. aureus is one of the most important pathogens associated with medical device biofilms and poses a major medical challenge due to its high incidence and drug resistance. The virulence factor surface proteins of S. aureus can specifically bind to host extracellular matrix proteins, such as fibrinogen, fibronectin, and collagen. These proteins have been shown to have a high affinity for implants and become easily attached to the implant surface. Therefore, these proteins mediate the attachment of S. aureus to the surface of the implant [156]. Targeting these mediator proteins to degrade or inhibit their production or reduce their adsorption capacity may provide a viable option for preventing S. aureus biofilm formation on implant surfaces. Fibrinolytic coatings (e.g., with tissue plasminogen activator (tPA)) reduce S. aureus biofilm formation on indwelling medical devices both in vitro and in vivo. The tPA coating prevents the early attachment of bacteria and the subsequent accumulation of biomass by splitting local fibrin. In addition, the tPA coating increases the sensitivity of biofilm infections to antibiotics [157]. Monomeric trimethylsilane and oxygen plasma coatings cause chemical changes on the surface of the material, resulting in changes in the adsorption of intermediate proteins on the coating surface, which can reduce the adhesion of bacteria to the surface, thereby inhibiting the formation of and infection caused by biofilms [158]. It has recently been demonstrated that device-related infections caused by S. aureus biofilms rely on the coagulation pathway to activate prothrombin and fibrin production. Direct thrombin inhibitors can be used to coat biomaterial surfaces to prevent staphylococcal binding and subsequent device-related infections [159].
Another approach is to inhibit the colonization of microorganisms and the formation of biofilms by affecting the structural integrity of the biofilm. The immobilization of biofilm-dispersing enzymes on material surfaces is regarded as one of the most promising strategies for preventing biofilm formation (Figure 1). For example, a deoxyribonuclease I (DNase I) coating has a good anti-infection ability and biocompatibility, which is beneficial for improving the application success rate of implants [160]. Polypyrrole is an organic conductive polymer formed from a pyrrole ring structure that affects the integrity of biofilms by promoting a structure with repeated and positive charge-regulated ionic cross-bridging and subsequent aggregation so that biofilms can be easily washed away by water [161]. The combined application of the above measures may lead to stronger antibiofilm activity. Regardless of the measures taken, the safety of the host must be considered, and the functionality of the raw materials must be ensured.
As mentioned above, in order to better protect implanted medical devices from bacterial invasion, preclinical studies on the surface modification of medical devices have made very successful progress, but there is still a significant discrepancy between the proposed strategies and their clinical application. Most of the coatings being studied may not be suitable for medical implants due to cytotoxicity, immunoreactivity, or genotoxicity problems. However, some technologies have shown sufficient evidence to prove the effectiveness, safety, and durability of antibacterial drug delivery coatings [146,162], such as AgNP coatings, which could be proposed in clinical practice. Some have been applied in clinical practice. For example, the antimicrobial hydrogel-coated prosthesis can safely protect the prosthesis and reduced early surgical site infections without adverse effects at a mean follow-up of 2 years [163]. Another study showed that silver-coated megaprosthesis is effective in terms of a reduction in prosthetic joint infections, with 2.2% local argyria and without systemic side effects [164]. In order to be better applied in clinical practice, the development of device surfaces with excellent antibacterial properties, high biocompatibility, and a long-lasting antibacterial efficacy still needs a breakthrough.

{kind=link}
Table 2.
Summary of novel surface-modifying compounds used against biofilms and mechanisms of action.
Table 2.
Summary of novel surface-modifying compounds used against biofilms and mechanisms of action.
| Surface-Modifying Compounds | Pathogens | Mechanism of Action | Reference |
|---|---|---|---|
| Isoeugenol | S. aureus, L. monocytogenes, and P. fluorescens | Antibacterial | [141] |
| N-acetyl cysteine and chitosan film | S. aureus | Antibacterial and antiadhesion | [144] |
| AgNP | E. coli and S. aureus | Antibacterial and antiadhesion | [146] |
| Silver-containing phosphonate monolayers | E. coli and S. epidermidis | Antiadhesion | [147] |
| Hydrophobin (Vmh2 and Pac3) | S. epidermidis | Antiadhesion | [152] |
| ECA | C. albicans | Antiadhesion | [153] |
| Poly(glycidol) | S. aureus | Antiadhesion | [154] |
| tPA | S. aureus | Antiadhesion, increases the sensitivity of biofilm infections to antibiotics | [157] |
| TMS/O2 | S. aureus | Antiadhesion | [158] |
| Direct thrombin inhibitors (argatroban, hirudin, and dabigatran) | staphylococcal | Antiadhesion | [159] |
| DNase I | S. mutans and S. aureus | Affect the structural integrity of the biofilm | [160] |
| Polypyrrole | S. mutans and S. sanguinis | Affect the integrity of biofilms | [161] |
3.3. Antimicrobial Peptides
Antimicrobial peptides (AMPs), which are a type of small-molecular peptide and are also known as host defense peptides (HDPs) [10], widely exist in nature and are a part of the innate immunity of almost all living organisms. AMPs can be extracted from bacteria, fungi, plants, insects, amphibians, fish, birds, mammals, and even the human body and can also be produced by chemical synthesis. AMPs have broad-spectrum and rapid antibacterial activity and do not easily result in drug resistance. In particular, their killing effect on certain drug-resistant pathogens has attracted attention. They can be used as single antibacterial agents or combined with traditional antibiotics.
To date, a variety of AMPs have been used in in vivo and in vitro studies, and some have been used in clinical practice, such as colicins and daptomycin. AMPs can not only exhibit powerful bactericidal activity, inhibit the formation of biofilms, and reduce or eliminate mature biofilms when the concentration is equal to or higher than the minimum inhibitory concentration (MIC) of the corresponding planktonic cells, but also display antibiofilm activity when the concentration is lower than the MIC of the corresponding planktonic cells [10]. This may be related to the mechanism underlying the multimode antibiofilm activity of AMPs. AMPs act on the bacterial cell membrane, forming different complex structures on the membrane through different mechanisms, such as the formation of barrel-state, toroidal pore, carpet, and aggregate models, which destroy the integrity and stability of the membrane, cause leakage of the cellular contents, and lead to the death of bacterial cells [165,166,167,168,169,170,171]. In addition, there are other potential antibiofilm mechanisms (Figure 1). Some AMPs can interfere with cellular metabolic processes and inhibit cell wall, nucleic acid, or protein biosynthesis by interacting with intracellular targets [172]. For example, B22, which is a derivative of BMAP-27B, can penetrate and kill established Vibrio cholerae and P. aeruginosa biofilms. After treatment with nanomolar concentrations of B22, P. aeruginosa and V. cholerae cells display expression changes in a variety of physiological metabolic processes, including protein synthesis, peptide glycan biosynthesis, respiration, and the detoxification of ROS. The main role of B22 has been shown to be the induction of ROS production, which destroys bacterial molecules, resulting in slowed cell growth or death [172]. LL-37 can downregulate the expression of the biofilm-related genes rhlA and rhlB, inhibit the synthesis of rhamnosyltransferases, reduce biofilm formation, and degrade existing biofilms formed by P. aeruginosa [173]. In addition, LL-37 also has antibacterial effects on biofilms formed by other pathogens. The anti-biofilm effects and mechanisms of LL-37 and LL-37 derivatives against biofilms formed by different pathogens (such as S. epidermidis, E. coli, S. aureus, C. albicans, and A. baumannii) have been well-summarized by Zhang Kaiyu and other colleagues [1]. HBD3 and HBD3-derived peptides interfere with the expression of the adhesin gene and reduce the expression of adhesion proteins, which leads to a decrease in biofilm formation [174,175]. In vivo, HBD3 has also been demonstrated to inhibit Staphylococcus biofilm formation [176]. HBD3 and its derivatives are most commonly used to study anti-biofilm activity, and other defensins have also shown anti-biofilm activity in some studies. For example, HBD2 can inhibit P. aeruginosa biofilm formation independent of biofilm regulatory pathways [177]. HNP-1 derivatives can also inhibit and eradicate the biofilm formation of multidrug-resistant uropathogenic E. coli, which may be used as a potential source for the treatment of biofilm-associated bacterial diseases [178]. The peptide Nal-P-113 inhibits Porphyromonas gingivalis biofilm formation by inhibiting the synthesis of ATP-binding cassette (ABC) transporters and ATP-binding proteins [24,179]. ABC transporters play a role in the cell adhesion phase of biofilm formation [180]. Temporin L (TL), derived from frog skin, has been confirmed to have antibiofilm activity [181]. It has been shown to act on the bacterial FtsZ protein, which is a protein involved in Z-ring formation, to inhibit cell division, thereby slowing bacterial growth [182]. Some AMPs can inhibit microbial cell adhesion or disperse cellular aggregates in the early stages of biofilm formation. LL-37 can also promote twitching in P. aeruginosa and cause the bacteria to move instead of forming biofilms [173,183]. Twitching is a form of surface motility that is regulated by type IV pili [184]. In S. aureus, lL-37- and LL-37-derived peptides have also been proven to inhibit the formation and degradation of existing biofilms [185]. In P. aeruginosa, the antibiofilm peptide 1037 can also directly inhibit biofilms by reducing swimming and swarming motilities, stimulating twitching motility, and suppressing the expression of a variety of genes involved in biofilm formation [186]. Polymyxin B (PmB), which is a cationic bacterial AMP produced by Bacillus polymyxa, can affect V. cholerae flagella, resulting in a decrease in motility and a compromised ability to form biofilms [187]. The formation of biofilms can also be inhibited by interruption of the signaling system of bacterial cells. QS is a cell–cell communication process that regulates multiple physiological and biochemical functions [1]. The human cathelicidin AMP LL-37 can prevent biofilm formation by inhibiting QS in microorganisms [183]. The immunomodulatory peptide innate defense regulator (IDR)-1018 was able to potently prevent biofilm formation and eradicate preformed biofilms formed by diverse species of Gram-negative bacteria and the Gram-positive bacterium S. aureus by blocking (p)ppGpp and signaling nucleotides [188]. Persister cells are a small subgroup of bacterial cells that are in a dormant state and are highly resistant to antibiotics. Some AMPs can prevent the formation of biofilms and promote the degradation of biofilms by interfering with the formation of persister cells [189]. The representative AMP is SAAP-148. S. aureus, which survives under the action of high-dose rifampicin, can be completely cleared within 2 h by low concentrations of SAAP-1448 [190]. Temporin 1Tb, derived from frog skin, can also kill both developing and mature S. epidermidis biofilms [191]. Some AMPs can impair EPS production to cause structural changes and degradation of the biofilm matrix. The synthetic peptide P1, derived from a tick antifreeze protein, significantly reduced S. mutans biofilm formation in an in vitro microplate assay and in a saliva-coated hydroxyapatite disc tooth model through the degradation of EPS and destruction of the biofilm structure [192]. TL can also affect mature biofilms in Pseudomonas fluorescens, and a large portion of the preformed biofilm architecture was clearly perturbed by addition of the peptide, with a significant decrease in all biofilm surface parameters and the overall biomass [181]. Hepcidin 20, which is an antimicrobial peptide derived from the human liver, can reduce the mass of the extracellular matrix and change the structure of biofilms of S. epidermidis by targeting the intracellular polysaccharide adhesion [193]. The fish-derived AMP piscidin-3 has nucleosidase activity and can destroy the extracellular DNA of P. aeruginosa through its N-terminal coordination with Cu2+ [194]. (LIN-SB056-1)2-K was also proven to disrupt the P. aeruginosa biofilm structure in an ex vivo model [195]. Some AMPs, as immune modulators, recruit immune cells at the site of infection, enhance the activity of host immune cells, and regulate the inflammatory response [196]. Some AMPs can resist endotoxin activity and neutralize proinflammatory factors, which may cause serious complications and damage to the body [197]. Some AMPs also display a synergistic effect when combined with traditional antibiotics via the promotion of antibiotic uptake [198]. We have listed the representative AMPs and their antibiofilm effects and mechanisms of action in Table 3.
Some AMPs have been applied to multi-drug resistance in clinical practice, such as colistin and polymyxin B. The modes of their application include intravenous or topical administration (such as intrathecal injection and atomization inhalation) [199]. International consensus guidelines have recommended intravenous doses of colistin and polymyxin B based on the kidney function of patients [200]. Some AMPs are prepared as ointments for infections of superficial tissues or medical device surfaces [190]. Most of the new AMPs have not been applied to humans and the experiments have been carried out in vitro or in animal experiments. However, many studies have provided inspiring results and the in vivo data have provided valuable references for the therapeutic dose, safety, and stability of AMPs for further clinical practice in humans. For example, in an animal experiment, 50 mg/kg administration of the four cathelicidins (Cathelicidin-BF, Pc-CATH1, Cc-CATH2, Cc-CATH3) could significantly reduce the fungal counts in a murine oral candidiasis model induced by clinically isolated C. albicans with slight hemolytic activity. Cathelicidin-BF strongly inhibited C. albicans biofilm formation at 20 μg/mL [171]. In an in vitro experiment simulating real flow conditions in the human body, Temporin-L(TL) had an anti-biofilm effect without increasing cell damage, demonstrating that it has great potential for clinical application [181]. The anti-biofilm doses of AMPs in the above experiments are shown in Table 3.
AMPs represent promising compounds for antibiofilm strategies. However, the activity of AMPs is related to many factors, including the size, sequence, charge, structure, conformation, hydrophobicity, and amphiphilicity, which may lead to toxicity and side effects on host cells under some conditions [11]. Although these AMPs have good antibiofilm activity in vitro, their potential toxicity and poor stability under physiological conditions limit their application in the clinical treatment of biofilm infections [201]. To date, many studies have examined the relationship between the structure of AMPs and their antibiofilm activity and safety. The structure with the best antibacterial activity is determined through the artificial modification and synthesis of AMPs, with the aim of seeking suitable AMPs for clinical application. Many methods have been adopted, for example, substitution, cyclization, hybridization techniques, or modification of the C- and N-termini via acetylation and myristoylation [173,202,203,204,205,206]. The bactericidal activity of some AMPs can be increased by truncating or increasing the overall positive charge [172,207]. In addition, the application of some drug delivery systems can not only help AMPs increase the antibacterial activity, but also improve their stability under physiological conditions and reduce their cytotoxicity [168]. For example, the conjugation of AMPs with nanoparticles (NPs) leads to the higher penetration of biofilms and improves the antibacterial activity. At the same time, NPs provide protection to AMPs, preventing their degradation in different environments, such as enzymatic degradation [208].
In conclusion, AMPs have a variety of active antibiofilm mechanisms and have potential applications in clinical treatment to remove biofilms. Although a large number of preclinical and clinical studies are underway, focusing on improving the activity, reducing the host cell toxicity, and improving the stability under physiological conditions of AMPs, further in vivo studies are needed due to the complexity of the host environment.
3.4. Nanotechnology
Nanotechnology is used to study the properties and applications of NMs. Nanometer is a unit of length that is equal to one billionth of a meter or 10−9 m. NMs are materials with at least one dimension at the nanometer scale (1–100 nm) in three-dimensional space, and these materials exhibit some special physical and chemical properties that are different from those of macrolevel materials. NMs have a wide range of applications in the fields of electronics, optics, manufacturing, and biomedicine, and they have also been developed for use in a variety of medical diagnosis and treatment applications [209]. NMs, especially NPs, have an increased specific surface area and surface functionalization that enable them to interact closely with microbial membranes and exhibit antimicrobial activity, even at very small doses [210]. These properties contribute to the development of antibacterial drugs [210]. The antibacterial and antibiofilm activities of NPs have been widely demonstrated in different pathogens, and their biological activities depend on different characteristics, including the size, shape, surface charge, structure, material type, and concentration [210,211]. The antibiofilm mechanisms of NPs are as follows. NPs can come into contact with bacterial cell walls and damage them through different interactions, induce changes in the membrane shape and function, induce oxidative stress and act as catalysts to interact with microbial cells, disturb the respiratory chain function, cross microbe membranes, interfere with metabolic pathways, interact with the microbial cellular machinery to inhibit enzymes, inactivate proteins, or modify gene expression levels [212,213]. NPs can also be used as drug delivery carriers or as catalysts to promote the penetration of drugs into biofilms; improve the solubility, stability, and biocompatibility of drugs; or enhance the antibiofilm activity by physically destroying the structure of the biofilm [22]. The antibiofilm mechanism of NPs is well-described in the study by Zhang Kaiyu and others [1] (Figure 1). Nanotechnology has developed rapidly. As mentioned above, this technology has been applied for the surface modification and improvement of AMPs. In this section, we will focus on the research progress in NMs that have exhibited promising results against biofilms.
Antibiofilm NMs include inorganic materials (such as gold and silver) [214,215,216,217,218,219], polymers (such as the natural antibacterial substance chitosan, biodegradable polymer polylactic acid, and carbon-based polymer fullerene) [220,221,222,223], lipids (such as liposomes) [224,225], and molecular complexes (such as protein nanocomplexes and cyclodextrin nanocomplexes) [226]. The following studies are examples of different NMs that are used against biofilms. Metal NPs exhibit bactericidal activity against a variety of microorganisms by releasing metal ions and are effective bactericides. At the same time, they have also been proven to have an impact on biofilms. Silver is currently one of the most widely studied metals in NM research. Biosynthetic AgNPs from Phanerochaete chrysosporium with a diameter of 45 nm exhibited a high negative surface charge close to 30 mV and showed colloidal stability from pH 3 to 9 under conditions of a high ionic strength ([NaCl] = 10–500 mM). It was shown that the AgNPs could kill bacteria by acting on fatty acids of E. coli cells to destroy the integrity of cell membranes. Moreover, the results further demonstrated that these biosynthetic AgNPs displayed biofilm disruption activity. These biogenic AgNPs were added to preformed biofilms and incubated for 48 hours, leading to a 29% reduction in the biomass of E. coli biofilms and an 80% reduction in the biomass of C. albicans biofilms [212]. Another study showed that P. aeruginosa biofilms treated with green synthesized seabuckthorn AgNPs (SBT@AgNps) can significantly reduce the bacterial load in biofilms, disrupt the matrix structure and cell wall, and interact with the cell membrane. The results showed a significant reduction in the bacterial load in each biofilm stage. On the peak day, i.e., the 4th day of biofilm formation, a significant reduction of approximately 33% (MIC) and 54% (MBC) in the bacterial load of the SBT@AgNps-treated biofilms was observed compared to that of the control biofilms. A biofilm of P. aeruginosa was established on a coverslip and analyzed by field emission scanning electron microscopy (FESEM) on the peak day. The results showed that the intact three-dimensional structure of the biofilm treated at the MIC and MBC was disrupted to form plaques embedded with biofilm cells. A concentration-dependent biofilm matrix structure was observed upon treatment with SBT-AgNps. In addition, the interaction of SBT@AgNPs with the bacterial cell membrane and disruption of the bacterial wall were also observed. The biofilm disruption resulted from SBT@AgNPs-mediated bacterial killing, and these NPs interacted with bacterial cells. Moreover, SBT@AgNPs showed an excellent anti-QS ability, which may be helpful in controlling bacterial infection at initial stages by blocking bacterial communication [227]. AuNPs are also commonly used nanoparticles, and biosynthesized AuNPs from brown seaweed at sub-MICs could inhibit P. aeruginosa biofilm formation by more than 80% and eradicate mature biofilms. Furthermore, sub-MICs of biosynthesized AuNPs also attenuated the production of several important virulence factors, such as pyoverdine, pyocyanin, and rhamnolipids, and impaired bacterial swarming, swimming, and twitching motilities [228]. The antibacterial activity and antibiofilm potential of chitosan and its nanometer-scale derivatives have been demonstrated in a variety of microorganisms [229]. A recent study proved that chitosan NPs (CNPs) can inhibit S. aureus biofilm formation, damage mature biofilms, reduce EPS production, and lower the cell surface hydrophobicity. The results showed that the biofilms were inhibited by approximately 50% upon treatment with CNPs compared to the control group. The treatment of a preformed biofilm with CNPs also resulted in a reduction of approximately 15% in the biofilm mass. Chrysin, which is a flavone constituent of the Orocylumineicum vent, has already been demonstrated to have antibacterial properties. However, its low water solubility, low bioavailability, and rapid metabolism in the body limit its therapeutic applications. Chitosan was used as a nanocarrier loaded with chrysin to enhance the antibiofilm activity through a synergistic effect [220]. In addition to their inherent antibiofilm activity, NMs can also be used as carriers for antibiofilm agents or other biofilm scavengers. Rifampicin-conjugated silver (Rif-Ag) NPs enhance the biofilm penetrating power compared to rifampicin alone. The synthesized NPs inhibit >90% of biofilms formed by K. pneumoniae and methicillin-resistant Staphylococcus aureus (MRSA) at low doses and show 1.5–2 times more penetrating potential, as observed by biofilm eradication and percentage viability reduction, compared to rifampicin alone [230]. Some NMs, as response carriers, exert antibiofilm effects. For example, mesoporous polydopamine NPs can cooperate with PDT to combat biofilms. These NPs, when integrated on titanium surfaces with an attached indocyanine green photosensitizer, can reduce S. aureus biofilm formation by 95% in murine specimens exposed to light with a wavelength of 808 nm [115]. Another study showed, that under laser irradiation, AuNPs produce vapor nanobubbles (VNB) to interfere with the structure of biofilms, promote the penetration of tobramycin, and improve the bactericidal effect [214]. Some NMs have inherent antibiofilm activity, which can prevent microorganisms from adhering to devices and the formation of biofilms, destroy EPS, and kill biofilm microorganisms through different mechanisms. Some NMs, as carriers of antibiofilm agents or other biofilm scavengers, can enhance the solubility and stability of the drug, overcome the drug resistance mechanism, and increase the biocompatibility of the drug at the target site. However, some NMs act as responsive carriers of external forces such as light to exert antibiofilm effects [22]. NM composites can simultaneously exert antibiofilm effects through all of the above methods. We have summarized the methods and effects of different nanomaterials against biofilms in Table 4.
Nanomaterials have a broad prospect in the application of anti-biofilms for medical devices. Although a large number of studies have been devoted to the antimicrobial activity of nanomaterials, the heterogeneity of these studies (e.g., the study design, synthesis methods, study methods of antimicrobial properties, and bacterial species selection) makes it impossible to cross-interpret between them, which limits nanomaterial application prospects in antibacterial aspects. Therefore, studies on nanomaterials need to be homogenized to obtain more reliable data to provide a solid theoretical basis for clinical treatment [231]. According to the above studies, the application methods can be summarized as device coatings [147], topical applications [223], intraperitoneal injections [221], etc., all of which are in vitro and animal experiments.
3.5. Agents for Degradation of the Extracellular Matrix of Biofilms
The extracellular matrix plays an important role in the formation, development, and survival of biofilms. It is not only a protective barrier against external factors, but also a source of nutrients and enzymes, as well as connectors between cells [232]. Degradation of the matrix is a way to weaken biofilms, and several strategies have been proposed (Figure 1).
First, enzymes can degrade various EPS components. The bacterial extracellular polysaccharide polyβ-1,6-N-acetylglucosamine (PNAG) is the main extracellular matrix component of the biofilms of Gram-positive bacteria and Gram-negative bacteria [233]. Dispersin B, which is a β-N-acetylglucosaminosidase produced by the Gram-negative species Actinobacillus actinomycetemcomitans, shows great potential for hydrolyzing PNAG to disperse biofilms formed by various bacteria, but does not kill the bacteria [234,235]. Due to the biofilm dispersal activity of dispersin B, it is used to generate specific probes for monitoring the production and localization of PNAG in the process of biofilm formation and elucidating the early stages of PNAG-dependent biofilm formation [233]. eDNA contributes to the structural integrity of the matrix [236]. Deoxyribonuclease (DNase) can degrade the eDNA present in the matrix, thus disrupting the stability of the biofilm. The antibiofilm effect of DNase has been studied for several kinds of microorganisms, such as S. aureus, P. aeruginosa, E. coli, A. baumannii, Haemophilus influenzae, and K. pneumonia [237,238]. DNase I immobilized on a titanium surface can prevent the adhesion and biofilm formation of S. mutans and S. aureus and exhibits good biocompatibility [160]. DNase I added to mature biofilms can also degrade biofilms. DNase I was further found to be less effective against mixed-species biofilms than individual biofilms [238]. Many other enzymes, such as proteinase K, lysozyme [239], α amylase [240], and the glycoside hydrolases PelAh and PslGh [241], have been proven to have promising antibiofilm activity.
Second, cation chelators can not only breakdown the extracellular matrix and eradicate biofilms, but also act as enhancers for antimicrobial agents, thereby disabling resistance mechanisms. For example, branched polyethylenimine (BPEI) is a cationic polymer with a low molecular weight of 600 Da. A recent study on treatment with BPEI combined with oxacillin demonstrated that this combination therapy could eradicate methicillin-resistant Staphylococcus epidermidis (MRSE) biofilms and reduce the cell density of the biofilms by 100,000-fold. The strong synergistic effect between BPEI and lactam antibiotics is because BPEI can neutralize the anionic character of the biofilm, resulting in rupture of the EPS matrix, as well as destruction of the hydrophobic barrier, which prevents the permeation of hydrophilic antibiotics, allowing the antibiotics to enter and kill the bacteria [14]. Ethylene diamine tetraacetic acid (EDTA), which is a cationic chelating agent, has been shown to affect the structural integrity of biofilms through the chelation of calcium, iron, and magnesium in staphylococci and Candida spp. [242]. A study revealed that EDTA alone significantly decreased fungal metabolic activity in preformed C. albicans biofilms. Additionally, EDTA combined with fluconazole significantly reduced the growth of biofilms compared with fluconazole alone. In addition, the study also showed that the cell density decreased when biofilm cells were treated with EDTA alone or in combination with fluconazole [243].
In addition, a few compounds, such as ambroxol and N-acetylcysteine, have been reported to have active matrix destruction properties and can be used for antibiofilm purposes [232]. Therefore, matrix disruptive agents such as enzymes, cationic chelators, and compounds provide us with another effective treatment strategy against biofilm-associated infections. These biofilm-dispersing agents can be used to functionalize the surface of medical devices to inhibit the formation of biofilms and can also be combined with other antibiofilm agents or technologies to improve antibiofilm activity [234,235,244,245].
Moreover, several new compounds have also been shown to have antibiofilm activity (Figure 1). These compounds can be extracted from organisms or produced by chemical synthesis. Farnesol, which is a QS molecule, can be secreted by a variety of organisms, mainly Candida, and can also be produced by chemical synthesis [246]. A recent study demonstrated that farnesol has significant antibiofilm activity, disrupting the mycelia of Fusarium keratoplasticum and the biofilm extracellular matrix and preventing conidial adhesion, filamentation, and biofilm formation [247]. In addition, a study on the effects of farnesol on Candida dubliniensis biofilms indicated synergy between farnesol and fluconazole in resistant strains. This led to a reversal of fluconazole resistance, which is a crucial result that suggests a possible application of farnesol as an adjuvant therapeutic agent [248]. Another study showed that caffeine, which is a xanthine alkaloid compound, significantly inhibited the formation of P. aeruginosa biofilms at sub-MIC concentrations by interfering with QS [249]. β-Lapachone and lapachol oxime are semisynthetic derivatives of lapachol that were originally isolated from species of the Bignoniaceae family and have also been proven to be effective in inhibiting biofilm formation [250]. In addition, Warburgia ugandensis Sprague subspecies ugandensis is a plant that is widely distributed in Eastern, Central, and Southern Africa. Warburganal, polygodial, and alpha-linolenic acid (ALA) were the major bioactive compounds isolated from the acetone extract of W. ugandensis. These compounds have been shown to inhibit the formation of C. albicans and S. aureus biofilms and exhibit synergistic effects with azole compounds [251]. However, some compounds have also been shown to enhance biofilm formation. Nicotine, for example, promotes S. aureus biofilm formation by promoting initial bacterial attachment and extracellular DNA release [252]. These new compounds provide new perspectives and possibilities for antibiofilm agents, requiring further verification of the efficacy and safety in clinical applications.
3.6. Bacteriophage Therapy
Bacteriophages (or phages) are viruses that can infect and replicate in bacterial cells. They are widely found in soil and water bodies and play an important role in microbial physiology. Bacteriophages have strict host specificity, reside in susceptible host bacteria, and can lyse bacteria, so they are sometimes used as adjuvant therapy for local infections. However, because of the emergence of bacterial infections for which antibiotics and surgery are not sufficient for eradication, bacteriophage therapy has attracted attention, and it has become a possible choice for the treatment of refractory bacterial infections (Figure 1).
A recent study conducted by Di Luca’s group showed that phages can lyse bacterial cells in vitro, even inside biofilms. In particular, the antibiofilm properties of some phages have been observed, such as the ability to kill persistent cells and degrade the extracellular matrix. These properties may be due to the depolymerase activity of the virus-encoded enzyme, which degrades the extracellular polysaccharide of the matrix, allowing phages or antibiotics to reach the bacteria and kill them [253]. Phage lysins have also been effectively used to remove bacterial biofilms. For example, in vitro, the phage lysin LysGH15 can efficiently prevent the biofilm formation of staphylococcal species. LysGH15 also showed notable disruption of 24-h and 72-h biofilms formed by S. aureus and coagulase-negative species [254]. The ability of LysGH15 to inhibit biofilm formation is most likely attributable to its rapid bactericidal activity prior to biofilm formation. At higher doses, LysGH15 can also destroy biofilms because it can dissolve the bacteria in the biofilm, thereby destroying the integrity of the biofilm instead of destroying the biofilm matrix [254]. Similarly, the effectiveness of phages against biofilms was demonstrated in vivo. In the established murine model of prosthetic joint infection, the combination of PlySs2 (a bacteriophage-derived lysin) (2.5 mg/kg, Intraperitoneal, q24hr) with vancomycin (110 mg/kg, Subcutaneous, q12hr) can significantly reduce the bacterial load in periprosthetic tissues and on implant surfaces [255]. In a recent case reported by Mario Corbellino et al., bacteriophages cured urinary tract infections that were recurrent despite previous treatment with effective antibiotics and the replacement of ureteral stents. After two weeks of lytic phage therapy, the presence of the KPC producer K. pneumoniae was no longer found in the urine and ureteral stent of the patient, and no microbial colonization was observed during the following two consecutive years [256]. However, due to host specificity, phage tolerance, and the complexity of the mature biofilm structure, phages have difficulty completely removing biofilms. Moreover, personalized phage cocktail (mixed phages) therapy has been developed. These phages work together to expand the scope of action to increase the potential of phage therapy. A study of phage cocktails composed of five phages against orthopedic implant biofilm infections caused by S. aureus showed that viable bacterial numbers within biofilms on titanium surfaces were significantly reduced in vitro [257]. In another case reported by Qin Jinhong et al., a cocktail of bacteriophages (5 × 108 pfu/mL, 50 mL bladder irrigation and kidney irrigation, q48hr) was selected for activity against all 21 heterogeneous K. pneumoniae strains to eradicate multifocal colonization, combined with piperacillin/tazobactam treatment. Finally, the patient recovered, with an obviously improved bladder [258].
Bacteriophages are species specific against bacterial infections. Antibiotics can easily cause dysregulation or resistance in the bacterial population. Bacteria are less likely to develop tolerance to bacteriophages. Therefore, bacteriophage therapy has good prospects alone or in combination with antibiotics in the treatment of biofilm infections. However, more scientific and clinical studies are needed to understand its true efficacy, pharmacokinetics, and mode of administration to establish appropriate clinical treatment regimens, and it is also necessary to determine which pathogen infections can be treated with bacteriophage therapy [259].
4. Conclusions
Medical device-related biofilm infections remain a serious problem in clinical services. The high resistance of biofilms to current antibiotic treatments appears to be a major challenge in this field. Antibiotic treatment alone often fails to eradicate microbial biofilms. The surgical removal of implants is effective in preventing the recurrence of biofilm infections; however, not all patients need or can undergo surgical revision for the treatment of local infections. In recent years, with an in-depth understanding of the mechanism of biofilm resistance, many novel approaches have been developed to combat device-related biofilm infections, such as coatings, AMPs, and nanotechnology. Some of these strategies exhibit antibiofilm activities against multiple targets and have been applied to the clinical treatment of medical device-associated biofilms. By combining these promising agents with antibiotics, it may be possible to eliminate device-related biofilms in the future. However, to develop safe, effective, practical, and economically viable strategies to combat medical device-related biofilm infections, further careful study is needed in this area to establish appropriate clinical treatment protocols.
Author Contributions
Conceptualization, X.L. and Y.W.; validation, X.L., L.S., and Y.W.; writing—original draft preparation, X.L., L.S., and P.Z.; writing—review and editing, Y.W.; visualization, X.L. and Y.W.; supervision, Y.W.; project administration, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant No. 81801972.
Acknowledgments
The authors gratefully acknowledge the support from Kaiyu Zhang of the first hospital of Jilin University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, K.; Li, X.; Yu, C.; Wang, Y. Promising Therapeutic Strategies Against Microbial Biofilm Challenges. Front. Cell. Infect. Microbiol. 2020, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Genet. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Tseng, B.S.; Majerczyk, C.D.; Da Silva, D.P.; Chandler, J.R.; Greenberg, E.P.; Parsek, M.R. Quorum Sensing Influences Burkholderia thailandensis Biofilm Development and Matrix Production. J. Bacteriol. 2016, 198, 2643–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Van Gennip, M.; Christensen, L.D.; Alhede, M.; Phipps, R.; Jensen, P.Ø.; Christophersen, L.; Pamp, S.J.; Moser, C.; Mikkelsen, P.J.; Koh, A.Y.; et al. Inactivation of the rhlA gene in Pseudomonas aeruginosa prevents rhamnolipid production, disabling the protection against polymorphonuclear leukocytes. APMIS 2009, 117, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otoole, G.; Kaplan, H.B.; Kolter, R. Biofilm Formation as Microbial Development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Kumar, A.; Alam, A.; Rani, M.; Ehtesham, N.Z.; Hasnain, S.E. Biofilms: Survival and defense strategy for pathogens. Int. J. Med. Microbiol. 2017, 307, 481–489. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. Biochim. Biophys. Acta Biomembr. 2016, 1858, 1044–1060. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.C.; Leonard, A.; Schäuble, M.; Rieckmann, L.M.; Hoyer, J.; Maass, S.; Lalk, M.; Becher, D.; Pané-Farré, J.; Riedel, K. Virulence Factors Produced by Staphylococcus aureus Biofilms Have a Moonlighting Function Contributing to Biofilm Integrity. Mol. Cell. Proteom. 2019, 18, 1036–1053. [Google Scholar] [CrossRef]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial Biofilms: Development, Dispersal, and Therapeutic Strategies in the Dawn of the Postantibiotic Era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.K.; Wouters, C.L.; Moen, E.L.; Pusavat, J.; Rice, C.V. Antibiofilm Synergy of β-Lactams and Branched Polyethylenimine against Methicillin-ResistantStaphylococcus epidermidis. Biomacromolecules 2019, 20, 3778–3785. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.-F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanepps, J.S.; Younger, J.G. Implantable Device-Related Infection. Shock 2016, 46, 597–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryers, J.D. Medical biofilms. Biotechnol. Bioeng. 2008, 100, 1–18. [Google Scholar] [CrossRef]
- Darouiche, R.O. Treatment of Infections Associated with Surgical Implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. The Use of Probiotics to Fight Biofilms in Medical Devices: A Systematic Review and Meta-Analysis. Microorganisms 2020, 9, 27. [Google Scholar] [CrossRef]
- Sánchez-Gómez, S.; Martínez-De-Tejada, G. Antimicrobial Peptides as Anti-biofilm Agents in Medical Implants. Curr. Top. Med. Chem. 2016, 17, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.M.; Tran, H.; Booth, M.A.; Fox, K.E.; Nguyen, T.H.; Tran, N.; Tran, P.A. Nanomaterials for Treating Bacterial Biofilms on Implantable Medical Devices. Nanomaterials 2020, 10, 2253. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural Anti-biofilm Agents: Strategies to Control Biofilm-Forming Pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef] [Green Version]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Grosu, M.; Stavropoulos, E.; Chifiriuc, M.C.; Lazar, V. Microbial Biofilms in Urinary Tract Infections and Prostatitis: Etiology, Pathogenicity, and Combating strategies. Pathogens 2016, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Barat, L.; Torres, A. Biofilms in ventilator-associated pneumonia. Futur. Microbiol. 2016, 11, 1599–1610. [Google Scholar] [CrossRef] [PubMed]
- Malone, M.; Bjarnsholt, T.; McBain, A.; James, G.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R. The prevalence of biofilms in chronic wounds: A systematic review and meta-analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, J.; Machado, D.; Gomes, A.M.; Machado, I.; Santos, C.; Lima, N.; Carvalho, M.J.; Cabrita, A.; Rodrigues, A.; Martins, M. Deciphering the Contribution of Biofilm to the Pathogenesis of Peritoneal Dialysis Infections: Characterization and Microbial Behaviour on Dialysis Fluids. PLoS ONE 2016, 11, e0157870. [Google Scholar] [CrossRef] [Green Version]
- Gominet, M.; Compain, F.; Beloin, C.; Lebeaux, D. Central venous catheters and biofilms: Where do we stand in 2017? APMIS 2017, 125, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Septimus, E.J.; Schweizer, M.L. Decolonization in Prevention of Health Care-Associated Infections. Clin. Microbiol. Rev. 2016, 29, 201–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, W.; Silva, P.; Silva, R.; Silva, G.; Machado, G.; Coelho, L.; Correia, M. Staphylococcus aureus and Staphylococcus epidermidis infections on implants. J. Hosp. Infect. 2018, 98, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Kelkar, A.; Agarwal, A.G.; Jayaswal, D.; Schultz, C.; Jayaswal, A.; Goel, V.K.; Agarwal, A.K.; Gidvani, S. Implant Retention or Removal for Management of Surgical Site Infection After Spinal Surgery. Glob. Spine J. 2019, 10, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Sabir, N.; Ikram, A.; Zaman, G.; Satti, L.; Gardezi, A.; Ahmed, A.; Ahmed, P. Bacterial biofilm-based catheter-associated urinary tract infections: Causative pathogens and antibiotic resistance. Am. J. Infect. Control. 2017, 45, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Sharma, S.; Sharma, P.; Chandola, D.; Dang, S.; Gupta, S.; Gabrani, R. Escherichia coli biofilm: Development and therapeutic strategies. J. Appl. Microbiol. 2016, 121, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa Biofilm, a Programmed Bacterial Life for Fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Piperaki, E.-T.; Syrogiannopoulos, G.A.; Tzouvelekis, L.S.; Daikos, G.L. Klebsiella pneumoniae. Pediatr. Infect. Dis. J. 2017, 36, 1002–1005. [Google Scholar] [CrossRef]
- Eze, E.C.; Chenia, H.Y.; E El Zowalaty, M. Acinetobacter baumannii biofilms: Effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Genet. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Rao, Y.; Shang, W.; Yang, Y.; Zhou, R.; Rao, X. Fighting Mixed-Species Microbial Biofilms with Cold Atmospheric Plasma. Front. Microbiol. 2020, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Y.; Wu, H.; Høiby, N.; Molin, S.; Song, Z. Current understanding of multi-species biofilms. Int. J. Oral Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kreth, J.; Merritt, J.; Qi, F. Bacterial and Host Interactions of Oral Streptococci. DNA Cell Biol. 2009, 28, 397–403. [Google Scholar] [CrossRef]
- Santos, A.P.A.; Watanabe, E.; De Andrade, D. Biofilm on artificial pacemaker: Fiction or reality? Arquivos Brasileiros de Cardiologia 2011, 97, e113–e120. [Google Scholar] [CrossRef] [PubMed]
- Salmeri, M.; Sorbello, M.G.; Mastrojeni, S.; Santanocita, A.; Milazzo, M.; Di Stefano, G.; Scalia, M.; Addamo, A.; Toscano, M.A.; Stefani, S.; et al. Infections of cardiovascular implantable electronic devices: 14 years of experience in an Italian hospital. Infez. Med. 2016, 24, 131–136. [Google Scholar] [PubMed]
- Del Río, A.; Anguera, I.; Miró, J.M.; Mont, L.; Fowler, V.G.; Azqueta, M.; Mestres, C.A.; Hospital Clínic Endocarditis Study Group. Surgical Treatment of Pacemaker and Defibrillator Lead Endocarditis. Chest 2003, 124, 1451–1459. [Google Scholar] [CrossRef]
- Blomström-Lundqvist, C.; Traykov, V.; Erba, P.A.; Burri, H.; Nielsen, J.C.; Bongiorni, M.G.; Poole, J.; Boriani, G.; Costa, R.; Deharo, J.-C.; et al. European Heart Rhythm Association (EHRA) international consensus document on how to prevent, diagnose, and treat cardiac implantable electronic device infections—endorsed by the Heart Rhythm Society (HRS), the Asia Pacific Heart Rhythm Society (APHRS), the Latin American Heart Rhythm Society (LAHRS), International Society for Cardiovascular Infectious Diseases (ISCVID) and the European Society of Clinical Microbiology and Infectious Diseases (ESCMID) in collaboration with the European Association for Cardio-Thoracic Surgery (EACTS). Eurospace 2020, 22, 515–549. [Google Scholar] [CrossRef]
- Cahill, T.J.; Baddour, L.M.; Habib, G.; Hoen, B.; Salaun, E.; Pettersson, G.B.; Schäfers, H.J.; Prendergast, B.D. Challenges in Infective Endocarditis. J. Am. Coll. Cardiol. 2017, 69, 325–344. [Google Scholar] [CrossRef]
- Conklin, E.F.; Giannelli, S., Jr.; Nealon, T.F., Jr. Four hundred consecutive patients with permanent transvenous pacemakers. J Thorac. Cardiovasc. Surg. 1975, 69, 1–7. [Google Scholar] [CrossRef]
- Bluhm, G. Pacemaker infections. A clinical study with special reference to prophylactic use of some isoxazolyl penicillins. Acta Med. Scand. Suppl. 1985, 699, 1–62. [Google Scholar] [PubMed]
- Blanco-Guzman, M.; Wang, X.; Vader, J.M.; Olsen, M.A.; Dubberke, E.R. Epidemiology of Left Ventricular Assist Device Infections: Findings From a Large Nonregistry Cohort. Clin. Infect. Dis. 2021, 72, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Novosad, S.A.; Lake, J.; Nguyen, D.; Soda, E.; Moulton-Meissner, H.; Pho, M.T.; Gualandi, N.; Bepo, L.; Stanton, R.A.; Daniels, J.B.; et al. Multicenter Outbreak of Gram-Negative Bloodstream Infections in Hemodialysis Patients. Am. J. Kidney Dis. 2019, 74, 610–619. [Google Scholar] [CrossRef]
- Martin, K.; Lorenzo, Y.S.P.; Leung, P.Y.M.; Chung, S.; O’Flaherty, E.; Barker, N.; Ierino, F. Clinical Outcomes and Risk Factors for Tunneled Hemodialysis Catheter-Related Bloodstream Infections. Open Forum Infect. Dis. 2020, 7, ofaa117. [Google Scholar] [CrossRef]
- Taylor, G.; Gravel, D.; Johnston, L.; Embil, J.; Holton, D.; Paton, S.; Canadian Nosocomial Infection Surveillance Program; Canadian Hospital Epidemiology Committee. Incidence of bloodstream infection in multicenter inception cohorts of hemodialysis patients. Am. J. Infect. Control. 2004, 32, 155–160. [Google Scholar] [CrossRef]
- Parameswaran, R.; Sherchan, J.B.; Varma, D.M.; Mukhopadhyay, C.; Vidyasagar, S. Intravascular catheter-related infections in an Indian tertiary care hospital. J. Infect. Dev. Ctries. 2010, 5, 452–458. [Google Scholar] [CrossRef]
- Ikram, S.; Heikal, A.; Finke, S.; Hofgaard, A.; Rehman, Y.; Sabri, A.N.; Økstad, O.A. Bacillus cereus biofilm formation on central venous catheters of hospitalised cardiac patients. Biofouling 2019, 35, 204–216. [Google Scholar] [CrossRef] [Green Version]
- Cobrado, L.; Silva-Dias, A.; Azevedo, M.M.; Rodrigues, A. Anti-Candida activity of antimicrobial impregnated central venous catheters. Antimicrob. Resist. Infect. Control 2017, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Hansra, S.; Crockford, G.; Köster, W.; Allan, B.J.; Blondeau, J.M.; Lainesse, C.; White, A.P. Tetrasodium EDTA Is Effective at Eradicating Biofilms Formed by Clinically Relevant Microorganisms from Patients’ Central Venous Catheters. mSphere 2018, 3, e00525-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, R.; Brown, M.; Faria, R.; Fraser, C.; Donohue, C.; Rainford, N.; Grosso, A.; Sinha, A.K.; Dorling, J.; Gray, J.; et al. Antimicrobial-impregnated central venous catheters for preventing neonatal bloodstream infection: The PREVAIL RCT. Health Technol. Assess. 2020, 24, 1–190. [Google Scholar] [CrossRef]
- Khaled, J.M.; Alyahya, S.A.; Chenthis Kanisha, C.; Alharbi, N.S.; Kadaikunnan, S.; Ramachandran, G.; Alanzi, K.F.; Rajivgandhi, G.; Vimala, R.; Manoharan, N. Anti-biofilm activity of LC-MS based Solanum nigrum essential oils against multi drug resistant biofilm forming P. mirabilis. Saudi J. Biol. Sci. 2021, 28, 302–309. [Google Scholar] [CrossRef]
- Townsend, E.M.; Moat, J.; Jameson, E. CAUTI’s next top model—Model dependent Klebsiella biofilm inhibition by bacteriophages and antimicrobials. Biofilm 2020, 2, 100038. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.G.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Genet. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Obručová, H.; Kotásková, I.; Tihelková, R.; Holá, V.; Růžička, F.; Freiberger, T. Fluorescent Capillary Electrophoresis Is Superior to Culture in Detecting Candida Species from Samples of Urinary Catheters and Ureteral Stents with Mono- or Polyfungal Biofilm Growth. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Singhai, M.; Malik, A.; Shahid, M.; Malik, M.A.; Goyal, R. A study on device-related infections with special reference to biofilm production and antibiotic resistance. J. Glob. Infect. Dis. 2012, 4, 193–198. [Google Scholar] [CrossRef]
- Stensballe, J.; Tvede, M.; Looms, D.; Lippert, F.K.; Dahl, B.; Tønnesen, E.; Rasmussen, L.S. Infection Risk with Nitrofurazone-Impregnated Urinary Catheters in Trauma Patients. Ann. Intern. Med. 2007, 147, 285–293. [Google Scholar] [CrossRef]
- Alonso, B.; Fernández-Barat, L.; Di Domenico, E.G.; Marín, M.; Cercenado, E.; Merino, I.; De Pablos, M.; Muñoz, P.; Guembe, M. Characterization of the virulence of Pseudomonas aeruginosa strains causing ventilator-associated pneumonia. BMC Infect. Dis. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Thorarinsdottir, H.R.; Kander, T.; Holmberg, A.; Petronis, S.; Klarin, B. Biofilm formation on three different endotracheal tubes: A prospective clinical trial. Crit. Care 2020, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Papazian, L.; Klompas, M.; Luyt, C.-E. Ventilator-associated pneumonia in adults: A narrative review. Intensiv. Care Med. 2020, 46, 888–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, S.; Al Talhi, Y.M.; Aldabbagh, M.; Baksh, M.; Osman, M.; Azzam, M. The incidence of ventilator-associated pneumonia (VAP) in a tertiary-care center: Comparison between pre- and post-VAP prevention bundle. J. Infect. Public Health 2020, 13, 552–557. [Google Scholar] [CrossRef]
- Hart, R.; McNeill, S.; MacLean, S.; Hornsby, J.; Ramsay, S. The prevalence of suspected ventilator-associated pneumonia in Scottish intensive care units. J. Intensiv. Care Soc. 2019, 21, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Metersky, M.L.; Wang, Y.; Klompas, M.; Eckenrode, S.; Bakullari, A.; Eldridge, N. Trend in Ventilator-Associated Pneumonia Rates Between 2005 and 2013. JAMA 2016, 316, 2427–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, R.; Pesce, M.; Franchelli, S.; Baldelli, I.; De Maria, A.; Marchese, A. Phenotypic and genotypic characterization of Staphylococci causing breast peri-implant infections in oncologic patients. BMC Microbiol. 2015, 15, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, U.D. Breast Augmentation, Antibiotic Prophylaxis, and Infection: Comparative Analysis of 1,628 Primary Augmentation Mammoplasties Assessing the Role and Efficacy of Antibiotics Prophylaxis Duration. Aesthetic Plast. Surg. 2009, 34, 42–47. [Google Scholar] [CrossRef]
- Basile, A.R.; Basile, F. Late infection following breast augmentation with textured silicone gel–filled implants. Aesthetic Surg. J. 2005, 25, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-F.; Lin, S.-F.; Hung, C.-F.; Chou, P. Risk of infection is associated more with drain duration than daily drainage volume in prosthesis-based breast reconstruction. Medicine 2016, 95, e5605. [Google Scholar] [CrossRef] [PubMed]
- Kıvanç, S.A.; Arık, G.; Akova-Budak, B.; Kıvanç, M. Biofilm forming capacity and antibiotic susceptibility of Staphylococcus spp. with the icaA/icaD/bap genotype isolated from ocular surface of patients with diabetes. Malawi Med. J. 2018, 30, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.C.; Bredarioli, P.A.P.; Leandro, F.D.; Ferreira, C.B.R.J.; Veiga, S.M.O.M.; Dias, A.L.T. Ozone against Pseudomonas aeruginosa biofilms in contact lenses storage cases. Revista do Instituto de Medicina Tropical de São Paulo 2019, 61, e23. [Google Scholar] [CrossRef] [PubMed]
- Onurdağ, F.K.; Özkan, S.; Ozgen, S.; Olmuş, H.; Abbasoglu, U. Candida albicans and Pseudomonas aeruginosa adhesion on soft contact lenses. Graefe’s Arch. Clin. Exp. Ophthalmol. 2010, 249, 559–564. [Google Scholar] [CrossRef]
- Steele, K.R.; Szczotka-Flynn, L. Epidemiology of contact lens-induced infiltrates: An updated review. Clin. Exp. Optom. 2017, 100, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Fürst, M.M.; Salvi, G.E.; Lang, N.P.; Persson, G.R. Bacterial colonization immediately after installation on oral titanium implants. Clin. Oral Implant. Res. 2007, 18, 501–508. [Google Scholar] [CrossRef]
- Dhir, S. Biofilm and dental implant: The microbial link. J. Indian Soc. Periodontol. 2013, 17, 5–11. [Google Scholar] [CrossRef]
- Alrabiah, M.; Alshagroud, R.S.; Alsahhaf, A.; Almojaly, S.A.; Abduljabbar, T.; Javed, F. Presence of Candida species in the subgingival oral biofilm of patients with peri-implantitis. Clin. Implant. Dent. Relat. Res. 2019, 21, 781–785. [Google Scholar] [CrossRef]
- Lindhe, J.; Meyle, J.; Group D of the European Workshop on Periodontology. Peri-implant diseases: Consensus Report of the Sixth European Workshop on Periodontology. J. Clin. Periodontol. 2008, 35, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatorska, B.; Groger, M.; Moser, D.; Diab-Elschahawi, M.; Lusignani, L.S.; Presterl, E. Does Extracellular DNA Production Vary in Staphylococcal Biofilms Isolated from Infected Implants versus Controls? Clin. Orthop. Relat. Res. 2017, 475, 2105–2113. [Google Scholar] [CrossRef] [Green Version]
- Fily, F.; Jolivet-Gougeon, A.; Polard, E.; Gicquel, T.; Dupont, M.; Verdier, M.; Arvieux, C. Moxifloxacin-rifampicin combination for the treatment of non-staphylococcal Gram-positive orthopedic implant-related infections. Médecine et Maladies Infectieuses 2019, 49, 540–544. [Google Scholar] [CrossRef]
- Blom, A.W.; Taylor, A.H.; Pattison, G.; Whitehouse, S.L.; Bannister, G.C. Infection after total hip arthroplasty. J. Bone Jt. Surg. Br. Vol. 2003, 85, 956–959. [Google Scholar] [CrossRef] [Green Version]
- Marmor, S.; Kerroumi, Y.; Meyssonnier, V.; Lhotellier, L.; Mouton, A.; Graff, W.; Zeller, V. One-Stage Exchange Arthroplasty for Fistulizing Periprosthetic Joint Infection of the Hip: An Effective Strategy. Front. Med. 2020, 7, 540929. [Google Scholar] [CrossRef] [PubMed]
- Kunutsor, S.K.; Whitehouse, M.R.; Blom, A.W.; Beswick, A.D.; Team, I. Re-Infection Outcomes following One- and Two-Stage Surgical Revision of Infected Hip Prosthesis: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0139166. [Google Scholar] [CrossRef] [Green Version]
- Fowler, T.J.; Sayers, A.; Whitehouse, M.R. Two-stage revision surgery for periprosthetic joint infection following total hip arthroplasty. Ann. Transl. Med. 2019, 7, S261. [Google Scholar] [CrossRef]
- Kisil, O.V.; Efimenko, T.A.; Gabrielyan, N.I.; Efremenkova, O.V. Development of antimicrobial therapy methods to overcome the antibiotic resistance of Acinetobacter baumannii. Acta Nat. 2020, 12, 34–45. [Google Scholar] [CrossRef]
- Tseng, B.S.; Zhang, W.; Harrison, J.J.; Quach, T.P.; Song, J.L.; Penterman, J.; Singh, P.K.; Chopp, D.L.; Packman, A.I.; Parsek, M.R. The extracellular matrix protects Pseudomonas aeruginosabiofilms by limiting the penetration of tobramycin. Environ. Microbiol. 2013, 15, 2865–2878. [Google Scholar] [CrossRef] [Green Version]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA Acidifies Biofilms and Induces Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 60, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA Chelates Cations and Induces Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef] [Green Version]
- Driffield, K.; Miller, K.; Bostock, J.M.; O’Neill, A.J.; Chopra, I. Increased mutability of Pseudomonas aeruginosa in biofilms. J. Antimicrob. Chemother. 2008, 61, 1053–1056. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiao, P.; Wang, Y.; Hao, Y. Mechanisms and Control Measures of Mature Biofilm Resistance to Antimicrobial Agents in the Clinical Context. ACS Omega 2020, 5, 22684–22690. [Google Scholar] [CrossRef]
- Avila-Novoa, M.-G.; Solís-Velázquez, O.-A.; Rangel-López, D.-E.; González-Gómez, J.-P.; Guerrero-Medina, P.-J.; Gutiérrez-Lomelí, M. Biofilm Formation and Detection of Fluoroquinolone- and Carbapenem-Resistant Genes in Multidrug-ResistantAcinetobacter baumannii. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Coquet, L.; Junter, G.A.; Jouenne, T. Resistance of artificial biofilms of Pseudomonas aeruginosa to imipenem and tobramycin. J. Antimicrob. Chemother. 1998, 42, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirijant, N.; Sermswan, R.W.; Wongratanacheewin, S. Burkholderia pseudomallei resistance to antibiotics in biofilm-induced conditions is related to efflux pumps. J. Med. Microbiol. 2016, 65, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ling, N.; Gao, J.; Zhang, M.; Zhang, X.; Tong, L.; Ou, D.; Wang, Y.; Zhang, J.; Wu, Q. Short communication: Roles of outer membrane protein W (OmpW) on survival and biofilm formation of Cronobacter sakazakii under neomycin sulfate stress. J. Dairy Sci. 2018, 101, 2927–2931. [Google Scholar] [CrossRef]
- Mah, T.-F. Biofilm-specific antibiotic resistance. Futur. Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, M.; Brooks, B.D.; Brooks, A.E. The Complex Relationship between Virulence and Antibiotic Resistance. Genes 2017, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Staphylococcus aureus Biofilms Promote Horizontal Transfer of Antibiotic Resistance. Antimicrob. Agents Chemother. 2013, 57, 1968–1970. [Google Scholar] [CrossRef] [Green Version]
- Kouzel, N.; Oldewurtel, E.R.; Maier, B. Gene Transfer Efficiency in Gonococcal Biofilms: Role of Biofilm Age, Architecture, and Pilin Antigenic Variation. J. Bacteriol. 2015, 197, 2422–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Águila-Arcos, S.; Álvarez-Rodríguez, I.; Garaiyurrebaso, O.; Garbisu, C.; Grohmann, E.; Alkorta, I. Biofilm-Forming Clinical Staphylococcus Isolates Harbor Horizontal Transfer and Antibiotic Resistance Genes. Front. Microbiol. 2017, 8, 2018. [Google Scholar] [CrossRef] [PubMed]
- Saginur, R.; Stdenis, M.; Ferris, W.; Aaron, S.D.; Chan, F.; Lee, C.; Ramotar, K. Multiple Combination Bactericidal Testing of Staphylococcal Biofilms from Implant-Associated Infections. Antimicrob. Agents Chemother. 2006, 50, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhang, X.; Tian, Y.; Tian, D.; Zhang, B. Incidence of surgical-site infection following open reduction and internal fixation of a distal femur fracture. Medicine 2019, 98, e14547. [Google Scholar] [CrossRef]
- Hedequist, D.; Haugen, A.; Hresko, T.; Emans, J. Failure of Attempted Implant Retention in Spinal Deformity Delayed Surgical Site Infections. Spine 2009, 34, 60–64. [Google Scholar] [CrossRef]
- Darouiche, R.O. Device-Associated Infections: A Macroproblem that Starts with Microadherence. Clin. Infect. Dis. 2001, 33, 1567–1572. [Google Scholar] [CrossRef]
- Pitiriga, V.; Kanellopoulos, P.; Bakalis, I.; Kampos, E.; Sagris, I.; Saroglou, G.; Tsakris, A. Central venous catheter-related bloodstream infection and colonization: The impact of insertion site and distribution of multidrug-resistant pathogens. Antimicrob. Resist. Infect. Control. 2020, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Blot, S.I.; Depuydt, P.; Annemans, L.; Benoit, D.; Hoste, E.; De Waele, J.J.; Decruyenaere, J.; Vogelaers, D.; Colardyn, F.; Vandewoude, K.H. Clinical and Economic Outcomes in Critically Ill Patients with Nosocomial Catheter-Related Bloodstream Infections. Clin. Infect. Dis. 2005, 41, 1591–1598. [Google Scholar] [CrossRef]
- Subbarao, E.K.; Tarpay, M.M.; Marks, M.I. Soft-Tissue Infections Caused by Mycobacterium fortuitum Complex Following Penetrating Injury. Arch. Pediatr. Adolesc. Med. 1987, 141, 1018–1020. [Google Scholar] [CrossRef]
- Prażmo, E.J.; Kwaśny, M.; Łapiński, M.; Mielczarek, A. Photodynamic Therapy as a Promising Method Used in the Treatment of Oral Diseases. Adv. Clin. Exp. Med. 2016, 25, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Huang, Y.-Y.; Wang, Y.; Wang, X.; Hamblin, M.R. Antimicrobial Photodynamic Therapy to Control Clinically Relevant Biofilm Infections. Front. Microbiol. 2018, 9, 1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbanzadeh, R.; Assadian, H.; Chiniforush, N.; Parker, S.; Pourakbari, B.; Ehsani, B.; Alikhani, M.Y.; Bahador, A. Modulation of virulence in Enterococcus faecalis cells surviving antimicrobial photodynamic inactivation with reduced graphene oxide-curcumin: An ex vivo biofilm model. Photodiagnosis Photodyn. Ther. 2020, 29, 101643. [Google Scholar] [CrossRef]
- Yuan, Z.; Tao, B.; He, Y.; Mu, C.; Liu, G.; Zhang, J.; Liao, Q.; Liu, P.; Cai, K. Remote eradication of biofilm on titanium implant via near-infrared light triggered photothermal/photodynamic therapy strategy. Biomaterials 2019, 223, 119479. [Google Scholar] [CrossRef] [PubMed]
- Teirlinck, E.; Barras, A.; Liu, J.; Fraire, J.C.; Lajunen, T.; Xiong, R.; Forier, K.; Li, C.; Urtti, A.; Boukherroub, R.; et al. Exploring Light-Sensitive Nanocarriers for Simultaneous Triggered Antibiotic Release and Disruption of Biofilms Upon Generation of Laser-Induced Vapor Nanobubbles. Pharmaceutics 2019, 11, 201. [Google Scholar] [CrossRef] [Green Version]
- Sayar, F.; Chiniforush, N.; Bahador, A.; Etemadi, A.; Akhondi, N.; Azimi, C. Efficacy of antimicrobial photodynamic therapy for elimination of Aggregatibacter actinomycetemcomitans biofilm on Laser-Lok titanium discs. Photodiagnosis Photodyn. Ther. 2019, 27, 462–466. [Google Scholar] [CrossRef]
- Briggs, T.; Blunn, G.; Hislop, S.; Ramalhete, R.; Bagley, C.; McKenna, D.; Coathup, M. Antimicrobial photodynamic therapy-a promising treatment for prosthetic joint infections. Lasers Med. Sci. 2017, 33, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Geralde, M.C.; Leite, I.S.; Inada, N.M.; Salina, A.C.G.; Medeiros, A.I.; Kuebler, W.M.; Kurachi, C.; Bagnato, V.S. Pneumonia treatment by photodynamic therapy with extracorporeal illumination—An experimental model. Physiol. Rep. 2017, 5, e13190. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, M.X. A clinical review of phototherapy for psoriasis. Lasers Med. Sci. 2018, 33, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Granick, M.S.; Paribathan, C.; Shanmugam, M.; Ramasubbu, N. Direct-Contact Low-Frequency Ultrasound Clearance of Biofilm from Metallic Implant Materials. Eplasty 2017, 17, e13. [Google Scholar]
- Dong, Y.; Chen, S.; Wang, Z.; Peng, N.; Yu, J. Synergy of ultrasound microbubbles and vancomycin against Staphylococcus epidermidis biofilm. J. Antimicrob. Chemother. 2012, 68, 816–826. [Google Scholar] [CrossRef] [Green Version]
- Carmen, J.C.; Runyan, C.M.; Robison, R.A.; Nelson, J.L.; Beckstead, B.L.; Pitt, W.G.; Schaalje, G.B. Ultrasonic-enhanced gentamicin transport through colony biofilms of Pseudomonas aeruginosa and Escherichia coli. J. Infect. Chemother. 2004, 10, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, N.; Li, L.; Ma, Y.; Zhao, C.; Wu, Q.; Li, Y.; He, N.; Wang, X. The synergistic bactericidal effect of vancomycin on UTMD treated biofilm involves damage to bacterial cells and enhancement of metabolic activities. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Li, J.; Li, P.; Yu, J. Ultrasound Microbubbles Enhance the Activity of Vancomycin Against Staphylococcus epidermidis Biofilms In Vivo. J. Ultrasound Med. 2018, 37, 1379–1387. [Google Scholar] [CrossRef]
- LuTheryn, G.; Glynne-Jones, P.; Webb, J.S.; Carugo, D. Ultrasound-mediated therapies for the treatment of biofilms in chronic wounds: A review of present knowledge. Microb. Biotechnol. 2019, 13, 613–628. [Google Scholar] [CrossRef] [Green Version]
- He, N.; Hu, J.; Liu, H.; Zhu, T.; Huang, B.; Wang, X.; Wu, Y.; Wang, W.; Qu, D. Enhancement of Vancomycin Activity against Biofilms by Using Ultrasound-Targeted Microbubble Destruction. Antimicrob. Agents Chemother. 2011, 55, 5331–5337. [Google Scholar] [CrossRef] [Green Version]
- Rediske, A.M.; Roeder, B.L.; Nelson, J.L.; Robison, R.L.; Schaalje, G.B.; Robison, R.A.; Pitt, W.G. Pulsed Ultrasound Enhances the Killing ofEscherichia coli Biofilms by Aminoglycoside Antibiotics In Vivo. Antimicrob. Agents Chemother. 2000, 44, 771–772. [Google Scholar] [CrossRef] [Green Version]
- Bigelow, T.A.; Thomas, C.L.; Wu, H.; Itani, K.M.F. Impact of High-Intensity Ultrasound on Strength of Surgical Mesh When Treating Biofilm Infections. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2018, 66, 38–44. [Google Scholar] [CrossRef]
- Ng, E.; Lim, L.P. An Overview of Different Interdental Cleaning Aids and Their Effectiveness. Dent. J. 2019, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Frascella, J.A.; Fernández, P.; Gilbert, R.D.; Cugini, M. A randomized, clinical evaluation of the safety and efficacy of a novel oral irrigator. Am. J. Dent. 2000, 13, 55–58. [Google Scholar]
- Kato, K.; Tamura, K.; Nakagaki, H. Quantitative evaluation of the oral biofilm-removing capacity of a dental water jet using an electron-probe microanalyzer. Arch. Oral Biol. 2012, 57, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Sharab, L.; Baier, R.; Ciancio, S.; Mang, T. Influence of Photodynamic Therapy on Bacterial Attachment to Titanium Surface. J. Oral Implant. 2020. [Google Scholar] [CrossRef]
- Yamada, J.; Takiguchi, T.; Saito, A.; Odanaka, H.; Soyama, H.; Yamamoto, M. Removal of Oral Biofilm on an Implant Fixture by a Cavitating Jet. Implant. Dent. 2017, 26, 904–910. [Google Scholar] [CrossRef]
- Toma, S.; Behets, C.; Brecx, M.C.; Lasserre, J.F. In Vitro Comparison of the Efficacy of Peri-Implantitis Treatments on the Removal and Recolonization of Streptococcus gordonii Biofilm on Titanium Disks. Materials 2018, 11, 2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.; Rudolph, M.; Bause, V.; Terfort, A. Electrochemical removal of biofilms from titanium dental implant surfaces. Bioelectrochemistry 2018, 121, 84–94. [Google Scholar] [CrossRef]
- Kotsakis, G.A.; Black, R.; Kum, J.; Berbel, L.; Sadr, A.; Karoussis, I.; Simopoulou, M.; Daubert, D. Effect of implant cleaning on titanium particle dissolution and cytocompatibility. J. Periodontol. 2020. [Google Scholar] [CrossRef]
- Li, H.; Fairfax, M.R.; Dubocq, F.; Darouiche, R.O.; Rajpurkar, A.; Thompson, M.; Tefilli, M.V.; Dhabuwala, C.B. Antibacterial Activity of Antibiotic Coated Silicone Grafts. J. Urol. 1998, 160, 1910–1913. [Google Scholar] [CrossRef]
- Dwyer, A. Reducing Tunneled Hemodialysis Catheter Morbidity: Surface-Treated Catheters—A Review. Semin. Dial. 2008, 21, 542–546. [Google Scholar] [CrossRef]
- Jennings, J.A.; Carpenter, D.P.; Troxel, K.S.; Beenken, K.E.; Smeltzer, M.S.; Courtney, H.S.; Haggard, W.O. Novel Antibiotic-loaded Point-of-care Implant Coating Inhibits Biofilm. Clin. Orthop. Relat. Res. 2015, 473, 2270–2282. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.; Subbiahdoss, G.; Zeng, G.; Salmi, Z.; Kjems, J.; Mygind, T.; Snabe, T.; Meyer, R. Antibacterial isoeugenol coating on stainless steel and polyethylene surfaces prevents biofilm growth. J. Appl. Microbiol. 2017, 124, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 4117–4129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Liu, Y. N-acetylcysteine inhibit biofilms produced by Pseudomonas aeruginosa. BMC Microbiol. 2010, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Sousa, D.M.; Parreira, P.; Lamghari, M.; Gomes, P.; Martins, M.C.L. N-acetylcysteine-functionalized coating avoids bacterial adhesion and biofilm formation. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Clement, J.L.; Jarrett, P.S. Antibacterial Silver. Met. Drugs 1994, 1, 467–482. [Google Scholar] [CrossRef]
- Paladini, F.; Pollini, M.; Deponti, D.; Di Giancamillo, A.; Peretti, G.; Sannino, A. Effect of silver nanocoatings on catheters for haemodialysis in terms of cell viability, proliferation, morphology and antibacterial activity. J. Mater. Sci. Mater. Med. 2013, 24, 1105–1112. [Google Scholar] [CrossRef]
- Tîlmaciu, C.-M.; Mathieu, M.; Lavigne, J.-P.; Toupet, K.; Guerrero, G.; Ponche, A.; Amalric, J.; Noël, D.; Mutin, P.H. In vitro and in vivo characterization of antibacterial activity and biocompatibility: A study on silver-containing phosphonate monolayers on titanium. Acta Biomater. 2015, 15, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zuber, F.; Maniura-Weber, K.; Brugger, J.; Ren, Q. Nanostructured surface topographies have an effect on bactericidal activity. J. Nanobiotechnology 2018, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Tang, Y.; Zeng, L.; Zhao, Y.; Ma, Z.; Sun, Z.; Xiang, L.; Ren, L.; Yang, K. In vitro and in vivo studies of anti-bacterial copper-bearing titanium alloy for dental application. Dent. Mater. 2018, 34, 1112–1126. [Google Scholar] [CrossRef]
- Sopata, M.; Karpiński, T.M.; Jakubowicz, J.; Sopata, M. Development of tantalum with highly hydrophilic surface and antimicrobial properties obtained by micro-arc oxidation process. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020. [Google Scholar] [CrossRef]
- Bottino, M.A.; Pereira, S.; Amaral, M.; Milhan, N.; Pereira, C.A.; Camargo, S.; Carvalho, A.; Melo, R.M. Streptococcus mutans Biofilm Formation and Cell Viability on Polymer-infiltrated Ceramic and Yttria-stabilized Polycrystalline Zirconium Dioxide Ceramic. Oper. Dent. 2019, 44, E271–E278. [Google Scholar] [CrossRef]
- Artini, M.; Cicatiello, P.; Ricciardelli, A.; Papa, R.; Selan, L.; Dardano, P.; Tilotta, M.; Vrenna, G.; Tutino, M.L.; Giardina, P.; et al. Hydrophobin coating prevents Staphylococcus epidermidis biofilm formation on different surfaces. Biofouling 2017, 33, 601–611. [Google Scholar] [CrossRef]
- Távora, F.F.F.; Chocano, A.P.C.; De Oliveira, D.G.; Pereira, J.R.; Almeida, R.S.; Neppelenbroek, K.H.; Porto, V.C. Beneficial Effects of Ethyl-Cyanoacrylate Coating Against Candida Albicans Biofilm Formation. Braz. Dent. J. 2019, 30, 266–271. [Google Scholar] [CrossRef]
- Lockhart, J.N.; Spoonmore, T.J.; McCurdy, M.W.; Rogers, B.R.; Guelcher, S.A.; Harth, E. Poly(glycidol) Coating on Ultrahigh Molecular Weight Polyethylene for Reduced Biofilm Growth. ACS Appl. Mater. Interfaces 2018, 10, 4050–4056. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Wozniak, D.J.; Stoodley, P.; Hall-Stoodley, L. Prevention and treatment ofStaphylococcus aureusbiofilms. Expert Rev. Anti-Infect. Ther. 2015, 13, 1499–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.; Na, M.; Jarneborn, A.; Jacobsson, G.; Peetermans, M.; Verhamme, P.; Jin, T. Tissue Plasminogen Activator Coating on Implant Surfaces Reduces Staphylococcus Aureus Biofilm Formation. Appl. Environ. Microbiol. 2015, 82, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Jones, J.E.; Yu, H.; Yu, Q.; Christensen, G.D.; Chen, M.; Sun, H. Nanoscale Plasma Coating Inhibits Formation of Staphylococcus aureus Biofilm. Antimicrob. Agents Chemother. 2015, 59, 7308–7315. [Google Scholar] [CrossRef] [Green Version]
- Hogan, S.; Kasotakis, E.; Maher, S.; Cavanagh, B.; O’Gara, J.P.; Pandit, A.; Keyes, T.E.; Devocelle, M.; O’Neill, E. A novel medical device coating prevents Staphylococcus aureus biofilm formation on medical device surfaces. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Ye, J.; Shao, C.; Zhang, X.; Guo, X.; Gao, P.; Cen, Y.; Ma, S.; Liu, Y. Effects of DNase I coating of titanium on bacteria adhesion and biofilm formation. Mater. Sci. Eng. C 2017, 78, 738–747. [Google Scholar] [CrossRef]
- Senpuku, H.; Tuna, E.B.; Nagasawa, R.; Nakao, R.; Ohnishi, M. The inhibitory effects of polypyrrole on the biofilm formation of Streptococcus mutans. PLoS ONE 2019, 14, e0225584. [Google Scholar] [CrossRef] [Green Version]
- Souza, J.G.S.; Bertolini, M.M.; Costa, R.C.; Nagay, B.E.; Dongari-Bagtzoglou, A.; Barão, V.A.R. Targeting implant-associated infections: Titanium surface loaded with antimicrobial. iScience 2021, 24, 102008. [Google Scholar] [CrossRef]
- Zoccali, C.; Scoccianti, G.; Biagini, R.; Daolio, P.A.; Giardina, F.L.; Campanacci, D.A. Antibacterial hydrogel coating in joint mega-prosthesis: Results of a comparative series. Eur. J. Orthop. Surg. Traumatol. 2021, 1–9. [Google Scholar] [CrossRef]
- Fiore, M.; Sambri, A.; Zucchini, R.; Giannini, C.; Donati, D.M.; De Paolis, M. Silver-coated megaprosthesis in prevention and treatment of peri-prosthetic infections: A systematic review and meta-analysis about efficacy and toxicity in primary and revision surgery. Eur. J. Orthop. Surg. Traumatol. 2021, 31, 201–220. [Google Scholar] [CrossRef]
- Sochacki, K.A.; Barns, K.J.; Bucki, R.; Weisshaar, J.C. Real-time attack on single Escherichia coli cells by the human antimicrobial peptide LL-37. Proc. Natl. Acad. Sci. USA 2011, 108, E77–E81. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Hanke, M.L.; Mishra, B.; Lushnikova, T.; Heim, C.E.; Chittezham Thomas, V.; Bayles, K.W.; Kielian, T. Transformation of Human Cathelicidin LL-37 into Selective, Stable, and Potent Antimicrobial Compounds. ACS Chem. Biol. 2014, 9, 1997–2002. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yang, G.; Lu, S.; Chen, D.; Fan, S.; Xu, J.; Wu, B.; He, J. Design and antimicrobial activities of LL-37 derivatives inhibiting the formation of Streptococcus mutans biofilm. Chem. Biol. Drug Des. 2019, 93, 1175–1185. [Google Scholar] [CrossRef]
- Patrulea, V.; Borchard, G.; Jordan, O. An Update on Antimicrobial Peptides (AMPs) and Their Delivery Strategies for Wound Infections. Pharmaceutics 2020, 12, 840. [Google Scholar] [CrossRef]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Topical antimicrobial peptide formulations for wound healing: Current developments and future prospects. Acta Biomater. 2020, 103, 52–67. [Google Scholar] [CrossRef]
- Kaplan, C.W.; Sim, J.H.; Shah, K.R.; Kolesnikova-Kaplan, A.; Shi, W.; Eckert, R. Selective Membrane Disruption: Mode of Action of C16G2, a Specifically Targeted Antimicrobial Peptide. Antimicrob. Agents Chemother. 2011, 55, 3446–3452. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Liu, X.; Wang, C.; Qiao, X.; Wu, S.; Wang, H.; Feng, L.; Wang, Y. Assessing the potential of four cathelicidins for the management of mouse candidiasis and Candida albicans biofilms. Biochimie 2016, 121, 268–277. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Kao, A.Y.; Prieto, A.C.; Pu, M.; Kao, C. Cathelicidin Peptides Restrict Bacterial Growth via Membrane Perturbation and Induction of Reactive Oxygen Species. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Dean, S.N.; Bishop, B.M.; Van Hoek, M.L. Susceptibility of Pseudomonas aeruginosa Biofilm to Alpha-Helical Peptides: D-enantiomer of LL-37. Front. Microbiol. 2011, 2, 128. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Tan, H.; Cheng, T.; Shen, H.; Shao, J.; Guo, Y.; Shi, S.; Zhang, X. Human β-defensin 3 inhibits antibiotic-resistant Staphylococcus biofilm formation. J. Surg. Res. 2013, 183, 204–213. [Google Scholar] [CrossRef]
- Geng, H.; Yuan, Y.; Adayi, A.; Zhang, X.; Song, X.; Gong, L.; Zhang, X.; Gao, P. Engineered chimeric peptides with antimicrobial and titanium-binding functions to inhibit biofilm formation on Ti implants. Mater. Sci. Eng. C 2018, 82, 141–154. [Google Scholar] [CrossRef]
- Li, S.; Zhu, C.; Fang, S.; Zhang, W.; He, N.; Xu, W.; Kong, R.; Shang, X. Ultrasound microbubbles enhance human β-defensin 3 against biofilms. J. Surg. Res. 2015, 199, 458–469. [Google Scholar] [CrossRef]
- Parducho, K.R.; Beadell, B.; Ybarra, T.K.; Bush, M.; Escalera, E.; Trejos, A.T.; Chieng, A.; Mendez, M.; Anderson, C.; Park, H.; et al. The Antimicrobial Peptide Human Beta-Defensin 2 Inhibits Biofilm Production of Pseudomonas aeruginosa Without Compromising Metabolic Activity. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Moazzezy, N.; Karam, M.R.A.; Rafati, S.; Bouzari, S.; Oloomi, M. Inhibition and eradication activity of truncated α-defensin analogs against multidrug resistant uropathogenic Escherichia coli biofilm. PLoS ONE 2020, 15, e0235892. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Lin, L.; Tan, L.-S.; Yu, H.-Y.; Cheng, J.-W.; Pan, Y.-P. Molecular pathways underlying inhibitory effect of antimicrobial peptide Nal-P-113 on bacteria biofilms formation of Porphyromonas gingivalis W83 by DNA microarray. BMC Microbiol. 2017, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O’Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef]
- Di Somma, A.; Recupido, F.; Cirillo, A.; Romano, A.; Romanelli, A.; Caserta, S.; Guido, S.; Duilio, A. Antibiofilm Properties of Temporin-L on Pseudomonas fluorescens in Static and In-Flow Conditions. Int. J. Mol. Sci. 2020, 21, 8526. [Google Scholar] [CrossRef] [PubMed]
- Di Somma, A.; Avitabile, C.; Cirillo, A.; Moretta, A.; Merlino, A.; Paduano, L.; Duilio, A.; Romanelli, A. The antimicrobial peptide Temporin L impairs E. coli cell division by interacting with FtsZ and the divisome complex. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129606. [Google Scholar] [CrossRef]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.; Rehm, B.H.; Hancock, R.E. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef] [Green Version]
- Whitchurch, C.B.; Hobbs, M.; Livingston, S.P.; Krishnapillai, V.; Mattick, J.S. Characterisation of a Pseudomonas aeruginosa twitching motility gene and evidence for a specialised protein export system widespread in eubacteria. Gene 1991, 101, 33–44. [Google Scholar] [CrossRef]
- Haisma, E.M.; De Breij, A.; Chan, H.; Van Dissel, J.T.; Drijfhout, J.W.; Hiemstra, P.S.; El Ghalbzouri, A.; Nibbering, P.H. LL-37-Derived Peptides Eradicate Multidrug-Resistant Staphylococcus aureus from Thermally Wounded Human Skin Equivalents. Antimicrob. Agents Chemother. 2014, 58, 4411–4419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente-Núñez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of Bacterial Biofilm Formation and Swarming Motility by a Small Synthetic Cationic Peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomucci, S.; Cros, C.D.-N.; Perron, X.; Mathieu-Denoncourt, A.; Duperthuy, M. Flagella-dependent inhibition of biofilm formation by sub-inhibitory concentration of polymyxin B in Vibrio cholerae. PLoS ONE 2019, 14, e0221431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente-Núñez, C.; Reffuveille, F.; Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Broad-Spectrum Anti-biofilm Peptide That Targets a Cellular Stress Response. PLoS Pathog. 2014, 10, e1004152. [Google Scholar] [CrossRef] [Green Version]
- Conlon, B.P.; Nakayasu, E.S.; Fleck, L.E.; LaFleur, M.D.; Isabella, V.M.; Coleman, K.; Leonard, S.N.; Smith, R.D.; Adkins, J.N.; Lewis, K. Activated ClpP kills persisters and eradicates a chronic biofilm infection. Nat. Cell Biol. 2013, 503, 365–370. [Google Scholar] [CrossRef] [Green Version]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; De Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; Van Der Heijde, T.; Boekema, B.K.; et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci. Transl. Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef] [Green Version]
- Maisetta, G.; Grassi, L.; Di Luca, M.; Bombardelli, S.; Medici, C.; Brancatisano, F.L.; Esin, S.; Batoni, G. Anti-biofilm properties of the antimicrobial peptide temporin 1Tb and its ability, in combination with EDTA, to eradicate Staphylococcus epidermidis biofilms on silicone catheters. Biofouling 2016, 32, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Ansari, J.M.; Abraham, N.M.; Massaro, J.; Murphy, K.; Smith-Carpenter, J.; Fikrig, E. Anti-Biofilm Activity of a Self-Aggregating Peptide against Streptococcus mutans. Front. Microbiol. 2017, 8, 488. [Google Scholar] [CrossRef]
- Brancatisano, F.L.; Maisetta, G.; Di Luca, M.; Esin, S.; Bottai, D.; Bizzarri, R.; Campa, M.; Batoni, G. Inhibitory effect of the human liver-derived antimicrobial peptide hepcidin 20 on biofilms of polysaccharide intercellular adhesin (PIA)-positive and PIA-negative strains of Staphylococcus epidermidis. Biofouling 2014, 30, 435–446. [Google Scholar] [CrossRef]
- Libardo, M.D.J.; Bahar, A.A.; Ma, B.; Fu, R.; McCormick, L.E.; Zhao, J.; McCallum, S.A.; Nussinov, R.; Ren, D.; Angeles-Boza, A.M.; et al. Nuclease activity gives an edge to host-defense peptide piscidin 3 over piscidin 1, rendering it more effective against persisters and biofilms. FEBS J. 2017, 284, 3662–3683. [Google Scholar] [CrossRef] [Green Version]
- Grassi, L.; Batoni, G.; Ostyn, L.; Rigole, P.; Van den Bossche, S.; Rinaldi, A.C.; Maisetta, G.; Esin, S.; Coenye, T.; Crabbé, A. The Antimicrobial Peptide lin-SB056-1 and Its Dendrimeric Derivative Prevent Pseudomonas aeruginosa Biofilm Formation in Physiologically Relevant Models of Chronic Infections. Front. Microbiol. 2019, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E. Host defense peptides: Front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Shurko, J.F.; Galega, R.S.; Li, C.; Lee, G.C. Evaluation of LL-37 antimicrobial peptide derivatives alone and in combination with vancomycin against S. aureus. J. Antibiot. 2018, 71, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Lora-Tamayo, J.; Murillo, O.; Ariza, J. Clinical Use of Colistin in Biofilm-Associated Infections. Adv. Exp. Med. Biol. 2019, 1145, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacother. J. Hum. Pharmacol. Drug Ther. 2019, 39, 10–39. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, S.; Shen, T.; Chen, L.; Zhou, J.; Shi, S.; Wang, Y.; Zhao, Z.; Liao, C.; Wang, C. N-terminal Myristoylation Enhanced the Antimicrobial Activity of Antimicrobial Peptide PMAP-36PW. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.; Zhou, T.; Zhang, J.; Xu, J.; Guo, X.; Hu, H.; Zhang, X.; Hu, M.; Li, J.; Yang, W.; et al. Enhanced cell selectivity of hybrid peptides with potential antimicrobial activity and immunomodulatory effect. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129532. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-L.; Su, P.-Y.; Kuo, S.-C.; Lauderdale, T.-L.Y.; Shih, C. Adding a C-terminal Cysteine (CTC) Can Enhance the Bactericidal Activity of Three Different Antimicrobial Peptides. Front. Microbiol. 2018, 9, 1440. [Google Scholar] [CrossRef]
- Lu, J.; Xu, H.; Xia, J.; Ma, J.; Xu, J.; Li, Y.; Feng, J. D- and Unnatural Amino Acid Substituted Antimicrobial Peptides with Improved Proteolytic Resistance and Their Proteolytic Degradation Characteristics. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Kao, C.; Lin, X.; Yi, G.; Zhang, Y.; Rowe-Magnus, D.A.; Bush, K. Cathelicidin Antimicrobial Peptides with Reduced Activation of Toll-Like Receptor Signaling Have Potent Bactericidal Activity against Colistin-Resistant Bacteria. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Almaaytah, A.; Mohammed, G.K.; Abualhaijaa, A.; Al-Balas, Q. Development of novel ultrashort antimicrobial peptide nanoparticles with potent antimicrobial and antibiofilm activities against multidrug-resistant bacteria. Drug Des. Dev. Ther. 2017, 11, 3159–3170. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Beack, S.; Han, S.; Shin, M.; Lee, T.; Park, Y.; Kim, K.S.; Yetisen, A.K.; Yun, S.H.; Kwon, W.; et al. Multifunctional Photonic Nanomaterials for Diagnostic, Therapeutic, and Theranostic Applications. Adv. Mater. 2018, 30. [Google Scholar] [CrossRef] [PubMed]
- Wnorowska, U.; Fiedoruk, K.; Piktel, E.; Prasad, S.V.; Sulik, M.; Janion, M.; Daniluk, T.; Savage, P.B.; Bucki, R. Nanoantibiotics containing membrane-active human cathelicidin LL-37 or synthetic ceragenins attached to the surface of magnetic nanoparticles as novel and innovative therapeutic tools: Current status and potential future applications. J. Nanobiotechnol. 2020, 18, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Manosalva, N.; Tortella, G.; Cristina Diez, M.; Schalchli, H.; Seabra, A.B.; Durán, N.; Rubilar, O. Green synthesis of silver nanoparticles: Effect of synthesis reaction parameters on antimicrobial activity. World J. Microbiol. Biotechnol. 2019, 35, 88. [Google Scholar] [CrossRef]
- Estevez, M.B.; Raffaelli, S.; Mitchell, S.G.; Faccio, R.; Alborés, S. Biofilm Eradication Using Biogenic Silver Nanoparticles. Molecules 2020, 25, 2023. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, C.; Guzmán-Moreno, J.; Ángeles-Chávez, C.; Rodríguez-González, V.; Ortega-Sigala, J.J.; Ramírez-Santoyo, R.M.; Vidales-Rodríguez, L.E. Biosynthesis of silver nanoparticles by Fusarium scirpi and its potential as antimicrobial agent against uropathogenic Escherichia coli biofilms. PLoS ONE 2020, 15, e0230275. [Google Scholar] [CrossRef] [Green Version]
- Teirlinck, E.; Xiong, R.; Brans, T.; Forier, K.; Fraire, J.; Van Acker, H.; Matthijs, N.; De Rycke, R.; De Smedt, S.C.; Coenye, T.; et al. Laser-induced vapour nanobubbles improve drug diffusion and efficiency in bacterial biofilms. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Habibipour, R.; Moradi-Haghgou, L.; Farmany, A. Green synthesis of AgNPs@PPE and its Pseudomonas aeruginosa biofilm formation activity compared to pomegranate peel extract. Int. J. Nanomed. 2019, 14, 6891–6899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, N.; James, B.; Devadathan, A.; Johny, M.K.; Mathew, J.; Jacob, J. Comparative Evaluation of Antibiofilm Efficacy of Chitosan Nanoparticle- and Zinc Oxide Nanoparticle-Incorporated Calcium Hydroxide-Based Sealer: An In vitro Study. Contemp. Clin. Dent. 2018, 9, 434–439. [Google Scholar]
- Niemirowicz, K.; Piktel, E.; Wilczewska, A.Z.; Markiewicz, K.H.; Durnaś, B.; Wątek, M.; Puszkarz, I.; Wróblewska, M.; Niklińska, W.; Savage, P.B.; et al. Core–shell magnetic nanoparticles display synergistic antibacterial effects against Pseudomonas aeruginosa and Staphylococcus aureus when combined with cathelicidin LL-37 or selected ceragenins. Int. J. Nanomed. 2016, 11, 5443–5455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban Florez, F.L.; Hiers, R.D.; Larson, P.; Johnson, M.; O’Rear, E.; Rondinone, A.J.; Khajotia, S.S. Antibacterial dental adhesive resins containing nitrogen-doped titanium dioxide nanoparticles. Mater. Sci. Eng. C 2018, 93, 931–943. [Google Scholar] [CrossRef]
- Duncan, B.; Li, X.; Landis, R.F.; Kim, S.T.; Gupta, A.; Wang, L.-S.; Ramanathan, R.; Tang, R.; Boerth, J.A.; Rotello, V.M. Nanoparticle-Stabilized Capsules for the Treatment of Bacterial Biofilms. ACS Nano 2015, 9, 7775–7782. [Google Scholar] [CrossRef] [Green Version]
- Siddhardha, B.; Pandey, U.; Kaviyarasu, K.; Pala, R.; Syed, A.; Bahkali, A.H.; Elgorban, A.M. Chrysin-Loaded Chitosan Nanoparticles Potentiates Antibiofilm Activity against Staphylococcus aureus. Pathogens 2020, 9, 115. [Google Scholar] [CrossRef] [Green Version]
- Scott, C.J.; Abdelghany, S.M.; Quinn, D.J.; Ingram, R.J.; Gilmore, B.F.; Donnelly, R.F.; Taggart, C.C. Gentamicin-loaded nanoparticles show improved antimicrobial effects towards Pseudomonas aeruginosa infection. Int. J. Nanomed. 2012, 7, 4053–4063. [Google Scholar] [CrossRef] [Green Version]
- Liakos, I.L.; Grumezescu, A.M.; Holban, A.M.; Florin, I.; D’Autilia, F.; Carzino, R.; Bianchini, P.; Athanassiou, A. Polylactic Acid—Lemongrass Essential Oil Nanocapsules with Antimicrobial Properties. Pharmaceuticals 2016, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Bregnocchi, A.; Zanni, E.; Uccelletti, D.; Marra, F.; Cavallini, D.; De Angelis, F.; De Bellis, G.; Bossù, M.; Ierardo, G.; Polimeni, A.; et al. Graphene-based dental adhesive with anti-biofilm activity. J. Nanobiotechnol. 2017, 15. [Google Scholar] [CrossRef]
- Guo, P.; Buttaro, B.A.; Xue, H.Y.; Tran, N.T.; Wong, H.L. Lipid-polymer hybrid nanoparticles carrying linezolid improve treatment of methicillin-resistant Staphylococcus aureus (MRSA) harbored inside bone cells and biofilms. Eur. J. Pharm. Biopharm. 2020, 151, 189–198. [Google Scholar] [CrossRef]
- Vera-González, N.; Bailey-Hytholt, C.M.; Langlois, L.; de Camargo Ribeiro, F.; de Souza Santos, E.L.; Junqueira, J.C.; Shukla, A. Anidulafungin liposome nanoparticles exhibit antifungal activity against planktonic and biofilm Candida albicans. J. Biomed. Mater. Res. Part A 2020, 108, 2263–2276. [Google Scholar] [CrossRef]
- Hu, D.; Deng, Y.; Jia, F.; Jin, Q.; Ji, J. Surface Charge Switchable Supramolecular Nanocarriers for Nitric Oxide Synergistic Photodynamic Eradication of Biofilms. ACS Nano 2019, 14, 347–359. [Google Scholar] [CrossRef]
- Gondil, V.S.; Kalaiyarasan, T.; Bharti, V.K.; Chhibber, S. Antibiofilm potential of Seabuckthorn silver nanoparticles (SBT@AgNPs) against Pseudomonas aeruginosa. 3 Biotech 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Manivasagan, P.; Lee, J.-W.; Pham, D.T.N.; Oh, J.; Kim, Y.-M. Fucoidan-Stabilized Gold Nanoparticle-Mediated Biofilm Inhibition, Attenuation of Virulence and Motility Properties in Pseudomonas aeruginosa PAO1. Mar. Drugs 2019, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.; Guo, F.; Niu, H.; Liu, Q.; Wang, S.; Duan, J. Chitosan Improves Anti-Biofilm Efficacy of Gentamicin through Facilitating Antibiotic Penetration. Int. J. Mol. Sci. 2014, 15, 22296–22308. [Google Scholar] [CrossRef] [Green Version]
- Farooq, U.; Ahmad, T.; Khan, A.; Sarwar, R.; Shafiq, J.; Raza, Y.; Ahmed, A.; Ullah, S.; Ur Rehman, N.; Al-Harrasi, A. Rifampicin conjugated silver nanoparticles: A new arena for development of antibiofilm potential against methicillin resistant Staphylococcus aureus and Klebsiella pneumoniae. Int. J. Nanomed. 2019, 14, 3983–3993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, R.E.; Gouyau, J.; Lamouroux, E. Limitations of Recent Studies Dealing with the Antibacterial Properties of Silver Nanoparticles: Fact and Opinion. Nanomaterials 2019, 9, 1775. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.M.; Soares, F.A.; Reis, S.; Nunes, C.; Van Dijck, P. Innovative Strategies Toward the Disassembly of the EPS Matrix in Bacterial Biofilms. Front. Microbiol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Eddenden, A.; Kitova, E.N.; Klassen, J.S.; Nitz, M. An Inactive Dispersin B Probe for Monitoring PNAG Production in Biofilm Formation. ACS Chem. Biol. 2020, 15, 1204–1211. [Google Scholar] [CrossRef]
- Donelli, G.; Francolini, I.; Romoli, D.; Guaglianone, E.; Piozzi, A.; Ragunath, C.; Kaplan, J.B. Synergistic Activity of Dispersin B and Cefamandole Nafate in Inhibition of Staphylococcal Biofilm Growth on Polyurethanes. Antimicrob. Agents Chemother. 2007, 51, 2733–2740. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-J.; Lee, C.-K. Twofold enhanced dispersin B activity by N-terminal fusion to silver-binding peptide for biofilm eradication. Int. J. Biol. Macromol. 2018, 118, 419–426. [Google Scholar] [CrossRef]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle–Biofilm Interactions: The Role of the EPS Matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef]
- Fleming, D.; Rumbaugh, K.P. Approaches to Dispersing Medical Biofilms. Microorganisms 2017, 5, 15. [Google Scholar] [CrossRef]
- Sharma, K.; Singh, A.P. Antibiofilm Effect of DNase against Single and Mixed Species Biofilm. Foods 2018, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- ElAdawy, M.; El-Mowafy, M.; El-Sokkary, M.M.A.; Barwa, R. Effects of Lysozyme, Proteinase K, and Cephalosporins on Biofilm Formation by Clinical Isolates of Pseudomonas aeruginosa. Interdiscip. Perspect. Infect. Dis. 2020, 2020, 6156720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, G.N.; Rodrigues, L.P.; Lopes, L.M.; Parisotto, T.M.; Steiner-Oliveira, C.; Nobre-Dos-Santos, M. Relationship amongαamylase and carbonic anhydrase VI in saliva, visible biofilm, and early childhood caries: A longitudinal study. Int. J. Paediatr. Dent. 2017, 27, 174–182. [Google Scholar] [CrossRef]
- Baker, P.; Hill, P.J.; Snarr, B.D.; Alnabelseya, N.; Pestrak, M.J.; Lee, M.J.; Jennings, L.K.; Tam, J.; Melnyk, R.A.; Parsek, M.R.; et al. Exopolysaccharide biosynthetic glycoside hydrolases can be utilized to disrupt and prevent Pseudomonas aeruginosa biofilms. Sci. Adv. 2016, 2, e1501632. [Google Scholar] [CrossRef] [Green Version]
- Raad, I.; Chatzinikolaou, I.; Chaiban, G.; Hanna, H.; Hachem, R.; Dvorak, T.; Cook, G.; Costerton, W. In Vitro and Ex Vivo Activities of Minocycline and EDTA against Microorganisms Embedded in Biofilm on Catheter Surfaces. Antimicrob. Agents Chemother. 2003, 47, 3580–3585. [Google Scholar] [CrossRef] [Green Version]
- Casalinuovo, I.A.; Sorge, R.; Bonelli, G.; Di Francesco, P. Evaluation of the antifungal effect of EDTA, a metal chelator agent, on Candida albicans biofilm. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1413–1420. [Google Scholar]
- Shao, C.; Zhang, X.; Ye, J.; Li, Y.-C.; Bao, Y.-J.; Li, Z.-H.; Huang, Y.; Liu, Y. Surface functionalization of titanium substrates with Deoxyribonuclease I inhibit peri-implant bacterial infection. Dent. Mater. J. 2020, 2020–2055. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Ludwig, R.; Schneider-Stickler, B. Co-immobilization of cellobiose dehydrogenase and deoxyribonuclease I on chitosan nanoparticles against fungal/bacterial polymicrobial biofilms targeting both biofilm matrix and microorganisms. Mater. Sci. Eng. C 2020, 108, 110499. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Černáková, L. Farnesol and Tyrosol: Secondary Metabolites with a Crucial quorum-sensing Role in Candida Biofilm Development. Genes 2020, 11, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kischkel, B.; Souza, G.K.; Chiavelli, L.U.R.; Pomini, A.M.; Svidzinski, T.I.E.; Negri, M. The ability of farnesol to prevent adhesion and disrupt Fusarium keratoplasticum biofilm. Appl. Microbiol. Biotechnol. 2020, 104, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Jabra-Rizk, M.A.; Shirtliff, M.; James, C.; Meiller, T. Effect of farnesol onCandida dubliniensisbiofilm formation and fluconazole resistance. FEMS Yeast Res. 2006, 6, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; Dastidar, D.G.; Paul, P.; Dutta, S.; Basu, D.; Sharma, S.R.; Basu, S.; Sarker, R.K.; Sen, A.; Sarkar, A.; et al. Inhibition of biofilm formation of Pseudomonas aeruginosa by caffeine: A potential approach for sustainable management of biofilm. Arch. Microbiol. 2019, 202, 623–635. [Google Scholar] [CrossRef]
- Fernandes, A.W.C.; Santos, V.L.D.A.; Araújo, C.R.M.; De Oliveira, H.P.; Da Costa, M.M. Anti-biofilm Effect of β-Lapachone and Lapachol Oxime Against Isolates of Staphylococcus aureus. Curr. Microbiol. 2019, 77, 204–209. [Google Scholar] [CrossRef]
- Kipanga, P.N.; Liu, M.; Panda, S.K.; Mai, A.H.; Veryser, C.; Van Puyvelde, L.; De Borggraeve, W.M.; Van Dijck, P.; Matasyoh, J.; Luyten, W. Biofilm inhibiting properties of compounds from the leaves of Warburgia ugandensis Sprague subsp ugandensis against Candida and staphylococcal biofilms. J. Ethnopharmacol. 2020, 248, 112352. [Google Scholar] [CrossRef]
- Shi, L.; Wu, Y.; Yang, C.; Ma, Y.; Zhang, Q.-Z.; Huang, W.; Zhu, X.-Y.; Yan, Y.-J.; Wang, J.-X.; Zhu, T.; et al. Effect of nicotine on Staphylococcus aureus biofilm formation and virulence factors. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Tkhilaishvili, T.; Lombardi, L.; Klatt, A.-B.; Trampuz, A.; Di Luca, M. Bacteriophage Sb-1 enhances antibiotic activity against biofilm, degrades exopolysaccharide matrix and targets persisters of Staphylococcus aureus. Int. J. Antimicrob. Agents 2018, 52, 842–853. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, M.; Zhang, H.; Dai, J.; Guo, Z.; Li, X.; Ji, Y.; Cai, R.; Xi, H.; Wang, X.; et al. Antibacterial Effects of Phage Lysin LysGH15 on Planktonic Cells and Biofilms of Diverse Staphylococci. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, B.R.; Niu, Y.; Turajane, K.; Staats, K.; Suhardi, V.; Carli, A.; Fischetti, V.; Bostrom, M.; Yang, X. 2020 John Charnley Award: The antimicrobial potential of bacteriophage-derived lysin in a murine debridement, antibiotics, and implant retention model of prosthetic joint infection. Bone Jt. J. 2020, 102 (Suppl. B), 3–10. [Google Scholar] [CrossRef]
- Corbellino, M.; Kieffer, N.; Kutateladze, M.; Balarjishvili, N.; Leshkasheli, L.; Askilashvili, L.; Tsertsvadze, G.; Rimoldi, S.G.; Nizharadze, D.; Hoyle, N.; et al. Eradication of a Multidrug-Resistant, Carbapenemase-Producing Klebsiella pneumoniae Isolate Following Oral and Intra-rectal Therapy with a Custom Made, Lytic Bacteriophage Preparation. Clin. Infect. Dis. 2019, 70, 1998–2001. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.; Kelly, N.; Elliott, L.; Grant, A.; Wilkinson, M.; Hazratwala, K.; McEwen, P. Evaluation of Bacteriophage Anti-Biofilm Activity for Potential Control of Orthopedic Implant-Related Infections Caused by Staphylococcus aureus. Surg. Infect. 2019, 20, 16–24. [Google Scholar] [CrossRef]
- Qin, J.; Wu, N.; Bao, J.; Shi, X.; Ou, H.; Ye, S.; Zhao, W.; Wei, Z.; Cai, J.; Li, L.; et al. Heterogeneous Klebsiella pneumoniae Co-infections Complicate Personalized Bacteriophage Therapy. Front. Cell. Infect. Microbiol. 2021, 10. [Google Scholar] [CrossRef]
- Cesta, N.; Di Luca, M.; Corbellino, M.; Tavio, M.; Galli, M.; Andreoni, M. Bacteriophage therapy: An overview and the position of Italian Society of Infectious and Tropical Diseases. Le Infezioni Medicina 2020, 28, 322–331. [Google Scholar]
Figure 1.
Stages of medical device-associated biofilm formation and approaches against biofilms.

Table 1.
The main microorganisms involved in the formation of biofilms and infection rates for different medical devices.
Table 1.
The main microorganisms involved in the formation of biofilms and infection rates for different medical devices.
| Medical Devices | Main Microorganisms | Infection Rates | Reference |
|---|---|---|---|
| Cardiac implantable devices | S. epidermidis, S. aureus, S. hominis | 0.13%–38.6% | [42,43,44,45,46,47,48,49] |
| Hemodialyzers | Staphylococci, P. aeruginosa, S. marcescens | ~18% | [28,50,51,52] |
| Center venous catheters | S. aureus, S. epidermidis, P. aeruginosa, candida, B. cereus | 3%–14% | [17,53,54,55,56,57] |
| Urinary catheters | E. coli, E. cloacae, P. mirabilis, Klebsiella, Enterococcus spp., P. aeruginosa, Candida | 9.1%–26.6% | [33,58,59,60,61,62,63] |
| Ventilator | P. aeruginosa, A. baumannii, Klebsiella species, S. maltophilia, Enterococcus spp., Candida spp. | 5%–40% | [37,62,64,65,66,67,68,69] |
| Artificial breasts | S. aureus and S. epidermidis | 0.5%–5.1% | [70,71,72,73] |
| Contact lenses | Staphylococcus spp., P. aeruginosa, C. albicans | 2.5%–6% | [74,75,76,77] |
| Dental prosthetics | Streptococcus, Fusobacterium, Capnocytophaga, Actinomyces and Candida species | 5.9%–56% | [78,79,80,81] |
| Orthopedic prosthetics | MRSA, MRSE, S. epidermis, S. aureus, P. acnes, E. faecalis | 1%–15% | [32,82,83,84,85,86,87] |
Table 3.
Representative AMPs and their antibiofilm effects and mechanisms of action.
| AMP | Pathogen | Infection Model | MIC | MBIC/EC | Effects and Mechanism of Action | Reference |
|---|---|---|---|---|---|---|
| IG-13-1 IG-13-2 (M) | S. mutans | in vitro | 5 μM 5 μM | 6.92 μM 7.58 μM | Disrupt the bacterial membrane, causing leakage of the contents; regulate the inflammatory response | [167] |
| B22 (M) | P. aeruginosa V. cholerae | in vitro | 4 μM 2 μM | 100 nM | Penetrate the membrane and kill bacteria by regulating physiological metabolic processes, including protein synthesis, peptide glycan biosynthesis, respiration, and detoxification of ROS | [172] |
| Nal-P-113 (M) | P. gingivalis | in vitro | NP | 6.25 μg/mL | Inhibit biofilm formation by inhibiting the synthesis of proteins that promote bacterial adhesion | [179] |
| Temporin-L (N) | P. fluorescens | ex vivo | >512 μM | 25 μM | Inhibit biofilm formation and disrupt biofilm structure | [181] |
| Temporin 1Tb (N) | S. epidermidis | in vitro | 25–50 μg/mL | 25–50 μg/mL | Eradicate established biofilms | [191] |
| 1037 (S) | P. aeruginosa B. cenocepacia L. monocytogenes | in vitro | 304 μg/mL >608 μg/mL 25 μg/mL | 10 μg/mL 5 μg/mL 0.63 μg/mL | Reduce swimming and swarming motilities, stimulate twitching motility, and suppress the expression of genes involved in biofilm formation | [186] |
| D-LL-37 (M) | P. aeruginosa | in vitro | > 1 μg/mL | 1 μg/mL | Promote bacterial twitching motility and inhibit biofilm formation | [173] |
| LL-37 (N) | P. aeruginosa | in vitro | 64 μg/mL | 0.5 μg/mL | Reduce swimming and swarming motilities, stimulate twitching motility, and interfere with QS | [183] |
| 1018 (S) | P. aeruginosa, E. coli A. baumannii B. cenocepacia S. enterica serovar Typhimurium K. pneumoniae, MRSA | in vitro | 64 µg/mL 32 µg/mL 128 µg/mL >256 µg/mL 64 µg/mL 8 µg/mL 64 µg/mL | 10 µg/mL 10 µg/mL 10 µg/mL 10 µg/mL 20 µg/mL 2 µg/mL 2.5 µg/mL | Inhibit biofilm formation by blocking the signal molecule (p)ppGpp, kill bacteria | [188] |
| ADEP4 (S) | MRSA | in vivo | 0.5 µg/mL | 5 µg/mL | Kill persister cells and eradicate biofilm infections by activating the ClpP protease | [189] |
| SAAP-148 (S) | S. aureus A. baumannii | in vivo | NP | 3.2 μM 1.6 μM | Prevent the formation of the extracellular matrix and promote its breakdown and eradication, eradicate persister cells | [190] |
| P1 (S) | S. mutans | in vitro | NP | 25 μg/mL | Disturb the biofilm architecture, resulting in a drastic reduction in attached biofilm biomass | [192] |
| Hepcidin 20 (S) | S. epidermidis | in vitro | >50 μM | 25 μM | Reduce the mass of the extracellular matrix and change the structure of biofilms by targeting PIA | [193] |
| HBD-3 (N) | S. epidermidis S. aureus | in vivo | 4 μg/mL 8 μg/mL | 40 μg/mL 80 μg/mL | Inhibit biofilm formation | [176] |
| TBP-1-GGG-hBD3-3 (S) | S. oralis S. gordonii S. sanguinis | in vitro | 320 μg/mL 500 μg/mL 500 μg/mL | 640 μg/mL 800 μg/mL 850 μg/ml | Reduce the expression of adhesion protein and inhibit biofilm formation | [175] |
| HDP3-Cu (M) | P. aeruginosa | in vitro | 32 μM | 2 μM | Destroy extracellular DNA | [194] |
| C16G2 (S) | S. mutans | in vitro | ~5 μM | 20 μM | Kill bacteria through disruption of the cell membrane | [170] |
| lin-SB056-1 (lin-SB056-1)2-K (S) | P. aeruginosa PAO1 | ex vivo | 4.8 μM 19.25 μM | NP 19.25 μM | Inhibit biofilm formation, kill bacteria, disrupt biofilm structure | [195] |
| Polymyxin B (N) | V. cholerae | in vitro | 100 μg/mL | 25 μg/mL | Affect flagella, reduce motility, inhibit biofilm formation | [187] |
| BF Pc-CATH1 Cc-CATH2 Cc-CATH3 (N) | C. albicans | in vivo | 1–16 μg/mL | 2.5 μg/mL NP NP NP | Inhibit biofilm formation, destroy preformed biofilms | [171] |
Note: N: naturally occurring AMPs; M: modification of naturally occurring AMPs; S: synthetic AMPs; MRSA: methicillin-resistant Staphylococcus aureus; PIA: polysaccharide intercellular adhesin; ROS: reactive oxygen species; MIC: minimum inhibitory concentration; MBIC: minimal biofilm inhibitory concentration; EC: experimental concentration; NP: not provided.
Table 4.
Summary of nanomaterials used against biofilms on medical devices.
| Category | Material | Shape and Size | Pathogens | MIC | MBIC/EC | Anti-Biofilm Mode | Effects | Reference |
|---|---|---|---|---|---|---|---|---|
| Inorganic | Biosynthesized AgNPs from Phanerochaete chrysosporium | spherical, ~45 nm | E. coli C. albicans | 0.25 nM NP | NP NP | Intrinsic antibiofilm activity | Reduce biomass of mature biofilms: 29% and 80% in E. coli and C. albicans biofilms, respectively | [212] |
| Biosynthesized AgNPs from Fusarium scirpi | spherical, 2–20 nm | E. coli | 25 mg/mL | 7.5 mg/L | Intrinsic antibiofilm activity | Inhibit 97% biofilm formation, kill 80% bacteria | [213] | |
| Biosynthesized AgNPs from pomegranate peel | spherical, 32–85 nm | P. aeruginosa | NP | 0.1 mg/mL | Intrinsic antibiofilm activity | Inhibit biofilm formation | [215] | |
| Biosynthesized AgNPs from seabuckthorn | spherical, 10–40 nm | P. aeruginosa | 2 μg/mL | 2 μg/mL | Intrinsic antibiofilm activity | Kill bacteria, disrupt biofilm structure and cell wall, interfere with cell membrane, and inhibit QS | [227] | |
| Biosynthesized AuNPs from brown seaweed | spherical, 15–119 nm | P. aeruginosa | 512 µg/mL | 128 µg/mL | Intrinsic antibiofilm activity | Inhibit>80% biofilm formation; eradicate mature biofilm; attenuate the production of virulence factors; impair bacterial swarming, swimming, and twitching motilities | [228] | |
| AgNPs + rifampicin | ellipsoidal and clumpy, 15–18 ± 4 nm | MRSA K. pneumoniae | 4 µg/mL 0.025 µg/mL | 1 µg/mL 0.003 µg/mL | Intrinsic antibiofilm activity and carrier | Inhibit >90% biofilm formation, eradicate 40-70% mature biofilms, kill ~50% bacteria, enhance the biofilm penetrating power of the drug | [230] | |
| ZnO NPs | NR | E. faecalis | NP | NP | Intrinsic antibiofilm activity | Disrupt biofilm structure, kill bacteria | [216] | |
| Core-shell MNPs (MNP@NH2, MNP@Au, MNP@PQAS) + LL-37 or LL-37 derivatives | spherical, 12, 11, and 13 nm | S. aureus P. aeruginosa | 128, 128, 128 μg/mL 128, 16, 64 μg/mL | 64, 64, 128 μg/mL 256, 64, 128 μg/mL | Carrier and magnetic field catalyst | Inhibit biofilm formation by 40%–85% | [217] | |
| AuNPs | 70 mm | B. multivorans, P. aeruginosa, and S. aureus | NP | 1.4E + 10 AuNP/mL | Light/thermal catalyst | Produce VNB that interfere with biofilm structure and disrupt biofilms | [214] | |
| TiO2 NPs+ nitrogen | spherical, 12 nm | S. mutans | NA | NA | Light/thermal catalyst | Kill bacteria | [218] | |
| SiO2 NPs+ peppermint oil + cinnamaldehyde | ~150 nm | E. coli, P. aeruginosa, and E. cloacae complex and S. aureus | NA | NA | Carrier | Deliver the essential oil payloads, effectively eradicate biofilms | [219] | |
| Polymer-based | Chitosan NPs | NR | E. faecalis | NP | NP | Intrinsic antibiofilm activity | Disrupt biofilm structure, kill bacteria | [216] |
| Chitosan NPs + chrysin | spherical, ~355 nm | S. aureus | 1024 µg/mL | 768 µg/mL | Intrinsic antibiofilm activity and carrier | Inhibit ~66% biofilm formation, disturb ~43% mature biofilm, reduce EPS production and bacterial cell surface hydrophobicity | [220] | |
| Mesoporous polydopamine NPs | 15.6 mm | S. aureus | NA | NA | Light/thermal catalyst | Inhibit biofilm formation by 95%, synergistic PDT/PTT effect (ROS generation/local hyperthermia) | [115] | |
| PLGA NPs + gentamicin | 251 nm | P. aeruginosa | 3 µg/mL | 6 μg/mL | Carrier | Control drug release for up to 16 days and enhance antimicrobial activity | [221] | |
| PLA NCs | spherical, 300 ± 110 nm | S. aureus, P. aeruginosa, E. coli, and C. albicans | NA | NA | Intrinsic antibiofilm activity and carrier | Inhibit biofilm formation | [222] | |
| Carbon-based | Graphene nanoplatelets | Thickness: ~1–~25 nm | S. mutans | NA | NA | Intrinsic antibiofilm activity | Inhibit bacterial adhesion, mechanical damage of cell wall, inhibit biofilm formation | [223] |
| Lipid | Lipid-polymer hybrid NPs + linezolid | - | MRSA | 0.9–1.9 µg/mL | 32 µg/mL | Carrier | Control drug release, suppress MRSA biofilm growth to 35%–60% | [224] |
| Liposomal NPs + anidulafungin | ~100 nm | C. albicans | 1.56–12.50 µg/mL | 1.25 mg/mL | Carrier | Disrupt preformed C. albicans biofilms, reduce fungal burden by as much as 99% | [225] | |
| Molecular polymer | Supramolecular polymer α-CD-Ce6-NO-DA | spherical ~136 nm | MRSA | NA | NA | Antibiofilm activity, carrier, respond to acid and light | Promote effective penetration of biofilms at pH 5.5, trigger the rapid release of NO, kill bacteria and improve the efficiency of PDT by reducing GSH and generating RNS, eradicate biofilms | [226] |
Note: MRSA: methicillin-resistant Staphylococcus aureus; MNPs: magnetic nanoparticles; MNP@NH2: aminosilane-coated MNPs; MNP@Au: gold-coated MNPs; MNP@PQAS: PQAS-coated MNPs; VNB: vapor nanobubbles; PLA: polylactic acid; PLGA: poly(lactide-co-glycolide); NC: nanocapsule; PDT: photodynamic therapy; GSH: glutathione; RNS: reactive nitrogen species; α-CD-Ce6-NO-DA: glutathione (GSH)-sensitive α-cyclodextrin (α-CD)-conjugated nitric oxide (NO) prodrug (α-CD-NO) and chlorin e6 (Ce6) prodrug (α-CD-Ce6) formed in a pH-sensitive poly(ethylene glycol) (PEG) block polypeptide copolymer (PEG-(KLAKLALALADA-2); NP: not provided; NA: not applicable.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, X.; Sun, L.; Zhang, P.; Wang, Y. Novel Approaches to Combat Medical Device-Associated BioFilms. Coatings 2021, 11, 294. https://0-doi-org.brum.beds.ac.uk/10.3390/coatings11030294
AMA Style
Li X, Sun L, Zhang P, Wang Y. Novel Approaches to Combat Medical Device-Associated BioFilms. Coatings. 2021; 11(3):294. https://0-doi-org.brum.beds.ac.uk/10.3390/coatings11030294
Chicago/Turabian StyleLi, Xin, Luyao Sun, Peng Zhang, and Yang Wang. 2021. "Novel Approaches to Combat Medical Device-Associated BioFilms" Coatings 11, no. 3: 294. https://0-doi-org.brum.beds.ac.uk/10.3390/coatings11030294
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.