Microscopy Methods for Biofilm Imaging: Focus on SEM and VP-SEM Pros and Cons

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Microscopy Techniques Applied to Biofilm Imaging
2.1. Light Microscopy (LM)
2.2. Confocal Laser Scanning Microscopy (CLSM)
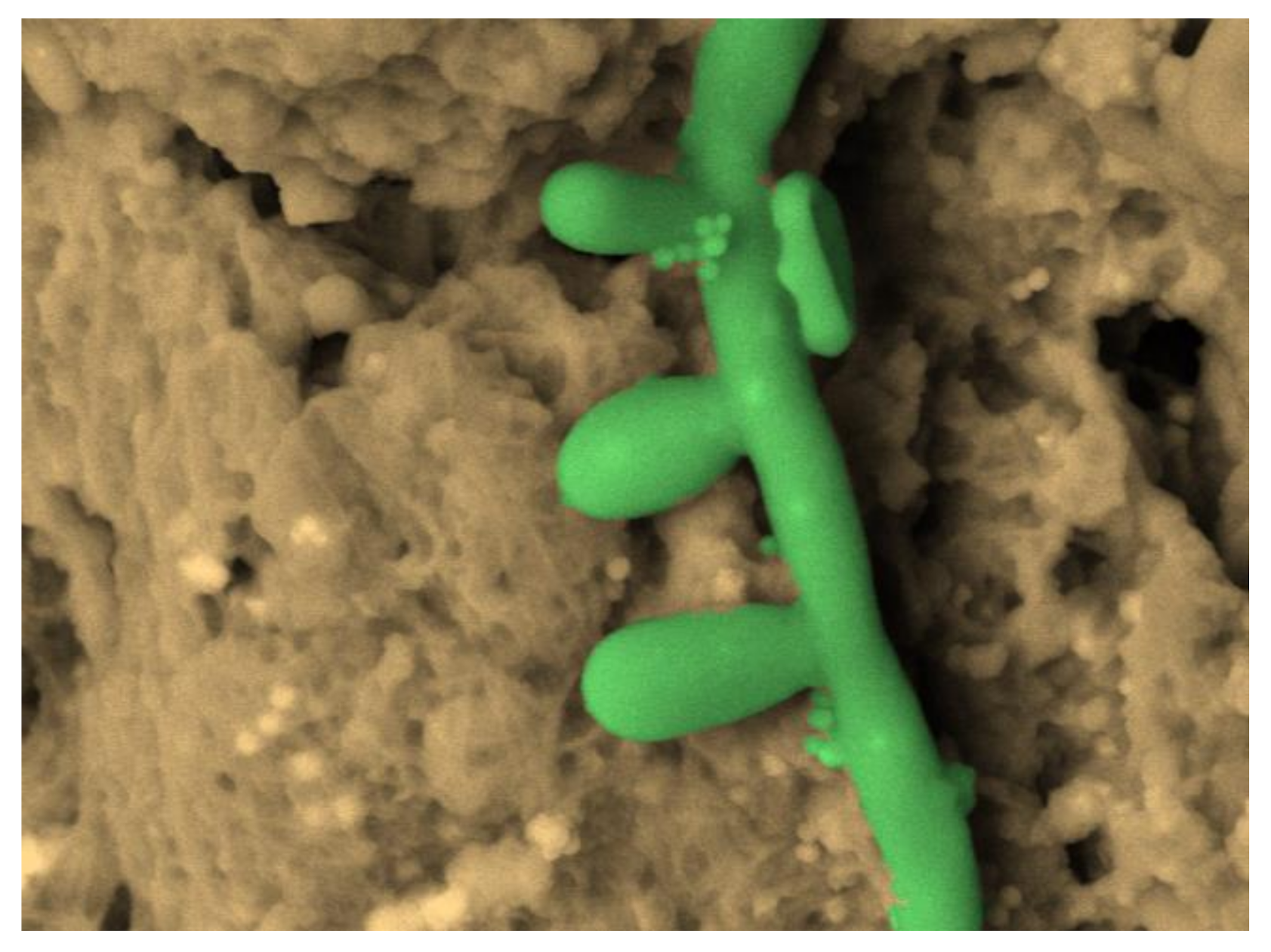
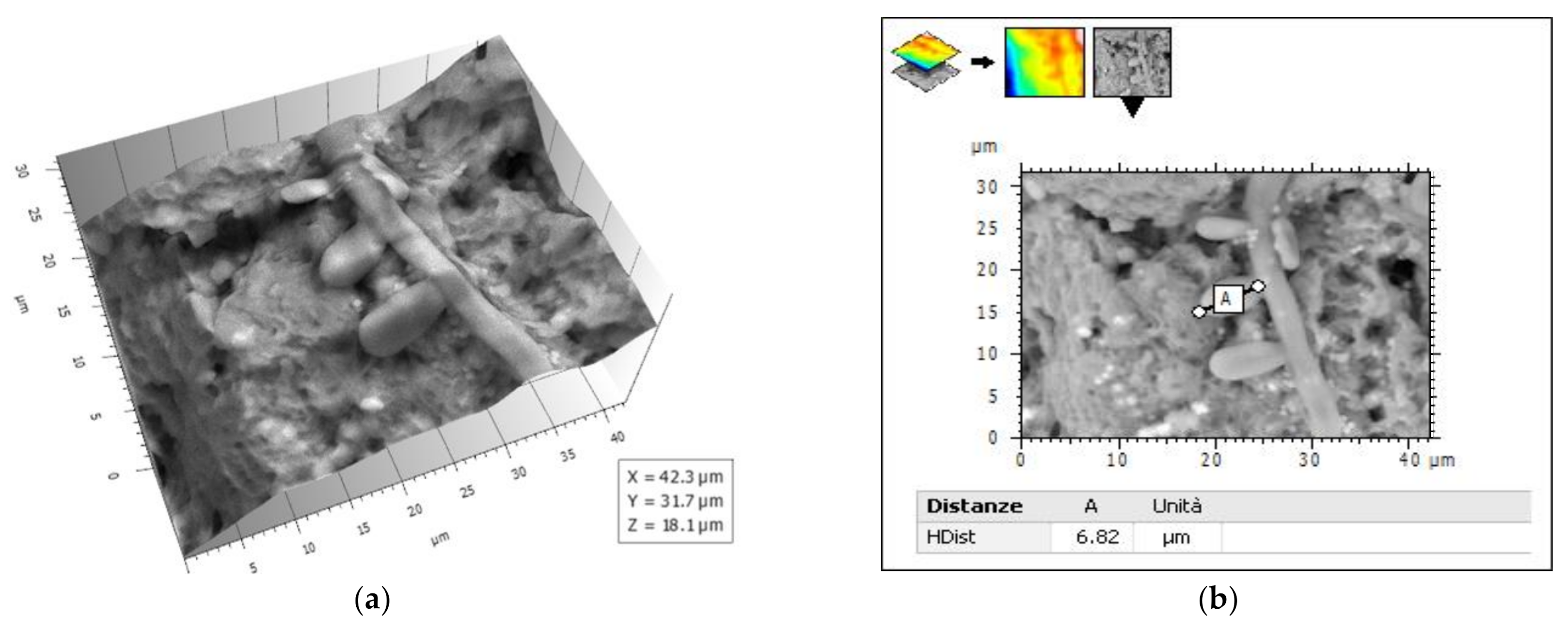
2.3. Atomic Force Microscopy (AFM)
3. Scanning Electron Microscopy Techniques Applied to Biofilm Study
3.1. Conventional Scanning Electron Microscope (SEM)
3.2. Variable Pressure Scanning Electron Microscopy (VP-SEM)
3.3. Comparing Conventional SEM and VP-SEM on S. mutans Samples
3.3.1. S. mutans Observed by Conventional SEM
3.3.2. S. mutans Observed by VP-SEM
3.3.3. S. mutans OsO4-RR-TA-IL Procedure Observed by SEM (High Vacuum, High Voltage Conditions)
3.4. Candida Albicans OsO4-RR-TA-IL Procedure Observed at SEM (High Vacuum, High Voltage Conditions)
3.5. Pros and Cons of Different SEM Protocols
4. Cryo-SEM
5. Environmental Scanning Electron Microscopy (ESEM)
6. Atmospheric Scanning Electron Microscopy (ASEM)
7. Focused Ion Beam-SEM (FIB-SEM)
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costerton, J.W.; Geesey, G.G.; Cheng, K.-J. How Bacteria Stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E. Oral Microbial Communities: Biofilms, Interactions, and Genetic Systems. Annu. Rev. Microbiol. 2000, 54, 413–437. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Thinking about bacterial populations as multicellular organisms. Annu. Rev. Microbiol. 1998, 52, 81–104. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, P. Extracellular polymeric substances, a key element in understanding biofilm phenotype. AIMS Microbiol. 2018, 4, 274–288. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Yadav, M.K.; Song, J.-J.; Singh, B.P.; Vidal, J.E. Chapter 1—Microbial biofilms and human disease: A concise review. In New and Future Developments in Microbial-l Biotechnology and Bioengineering; Current Research and Future Trends in Microbial Biofilms; Yadav, M.K., Singh, B.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–13. [Google Scholar]
- Suetens, C.; Latour, K.; Kärki, T.; Ricchizzi, E.; Kinross, P.; Moro, M.L.; Jans, B.; Hopkins, S.; Hansen, S.; Lyytikäinen, O.; et al. Prevalence of healthcare-associated infections, estimated incidence and composite antimicrobial resistance index in acute care hospitals and long-term care facilities: Results from two European point prevalence surveys, 2016 to 2017. Eurosurveillance 2018, 23, 1800516. [Google Scholar] [CrossRef] [Green Version]
- Breidenstein, E.B.; De La Fuente-Nunez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiao, P.; Wang, Y.; Hao, Y. Mechanisms and Control Measures of Mature Biofilm Resistance to Antimicrobial Agents in the Clinical Context. ACS Omega 2020, 5, 22684–22690. [Google Scholar] [CrossRef]
- Papa, R.; Garzoli, S.; Vrenna, G.; Sabatino, M.; Sapienza, F.; Relucenti, M.; Donfrancesco, O.; Fiscarelli, E.V.; Artini, M.; Selan, L.; et al. Essential Oils Biofilm Modulation Activity, Chemical and Machine Learning Analysis. Application on Staphylococcus aureus Isolates from Cystic Fibrosis Patients. Int. J. Mol. Sci. 2020, 21, 9258. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.D.; Dhong, H.-J.; Chung, S.-K.; Kim, H.Y.; Park, J.; Ha, S.Y. Hematoxylin and Eosin Staining for Detecting Biofilms: Practical and Cost-Effective Methods for Predicting Worse Outcomes After Endoscopic Sinus Surgery. Clin. Exp. Otorhinolaryngol. 2014, 7, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Bulut, F.; Meric, F.; Yorgancilar, E.; Nergiz, Y.; Akkus, M.; Nergiz, S.; Nasir, Y. Effects of N-acetyl-cysteine and acetylsalicylic acid on the tonsil bacterial biofilm tissues by light and electron microscopy. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 3720–3725. [Google Scholar] [PubMed]
- De Carvalho, C.C.C.R.; Da Fonseca, M.M.R.; Da Fonseca, M.M.R. Assessment of three-dimensional biofilm structure using an optical microscope. BioTechniques 2007, 42, 616–620. [Google Scholar] [CrossRef] [Green Version]
- Richardson, N.; Mordan, N.J.; Figueiredo, J.A.P.; Ng, Y.; Gulabivala, K. Microflora in teeth associated with apical periodontitis: A methodological observational study comparing two protocols and three microscopy techniques. Int. Endod. J. 2009, 42, 908–921. [Google Scholar] [CrossRef]
- Klausen, M.; Heydorn, A.; Ragas, P.; Lambertsen, L.; Aaes-Jørgensen, A.; Molin, S.; Tolker-Nielsen, T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol. 2003, 48, 1511–1524. [Google Scholar] [CrossRef]
- Bridier, A.; Dubois-Brissonnet, F.; Boubetra, A.; Thomas, V.; Briandet, R. The biofilm architecture of sixty opportunistic pathogens deciphered using a high throughput CLSM method. J. Microbiol. Methods 2010, 82, 64–70. [Google Scholar] [CrossRef]
- Guilbaud, M.; Piveteau, P.; Desvaux, M.; Brisse, S.; Briandet, R. Exploring the Diversity of Listeria monocytogenes Biofilm Architecture by High-Throughput Confocal Laser Scanning Microscopy and the Predominance of the Honeycomb-Like Morphotype. Appl. Environ. Microbiol. 2014, 81, 1813–1819. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liao, K.; Wang, D. Effects of Magnolol and Honokiol on Adhesion, Yeast-Hyphal Transition, and Formation of Biofilm by Candida albicans. PLoS ONE 2015, 10, e0117695. [Google Scholar] [CrossRef] [Green Version]
- Villacorte, L.O.; Ekowati, Y.; Neu, T.R.; Kleijn, J.M.; Winters, H.; Amy, G.; Schippers, J.; Kennedy, M. Characterisation of algal organic matter produced by bloom-forming marine and freshwater algae. Water Res. 2015, 73, 216–230. [Google Scholar] [CrossRef]
- Thornton, R.B.; Rigby, P.J.; Wiertsema, S.P.; Filion, P.; Langlands, J.; Coates, H.; Vijayasekaran, S.; Keil, A.D.; Richmond, P.C. Multi-species bacterial biofilm and intracellular infection in otitis media. BMC Pediatr. 2011, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridier, A.; Briandet, R.; Bouchez, T.; Jabot, F. A model-based approach to detect interspecific interactions during biofilm development. Biofouling 2014, 30, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Sanchez-Vizuete, M.D.P.; Le Coq, D.; Aymerich, S.; Meylheuc, T.; Maillard, J.-Y.; Thomas, V.; Dubois-Brissonnet, F.; Briandet, R. Biofilms of a Bacillus subtilis Hospital Isolate Protect Staphylococcus aureus from Biocide Action. PLoS ONE 2012, 7, e44506. [Google Scholar] [CrossRef] [PubMed]
- Doroshenko, N.; Tseng, B.S.; Howlin, R.P.; Deacon, J.; Wharton, J.A.; Thurner, P.J.; Gilmore, B.F.; Parsek, M.R.; Stoodley, P. Extracellular DNA Impedes the Transport of Vancomycin in Staphylococcus epidermidis Biofilms Preexposed to Subinhibitory Concentrations of Vancomycin. Antimicrob. Agents Chemother. 2014, 58, 7273–7282. [Google Scholar] [CrossRef] [Green Version]
- Marchal, M.; Briandet, R.; Halter, D.; Koechler, S.; Dubow, M.S.; Lett, M.-C.; Bertin, P.N. Subinhibitory Arsenite Concentrations Lead to Population Dispersal in Thiomonas sp. PLoS ONE 2011, 6, e23181. [Google Scholar] [CrossRef]
- Verma, V.; Harjai, K.; Chhibber, S. Structural changes induced by a lytic bacteriophage make ciprofloxacin effective against older biofilm ofKlebsiella pneumoniae. Biofouling 2010, 26, 729–737. [Google Scholar] [CrossRef]
- Hope, C.K.; Clements, D.; Wilson, M. Determining the spatial distribution of viable and nonviable bacteria in hydrated microcosm dental plaques by viability profiling. J. Appl. Microbiol. 2002, 93, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Dufrêne, Y.F.; Persat, A. Mechanomicrobiology: How bacteria sense and respond to forces. Nat. Rev. Microbiol. 2020, 18, 227–240. [Google Scholar] [CrossRef]
- Li, J.; Busscher, H.J.; Swartjes, J.J.T.M.; Chen, Y.; Harapanahalli, A.K.; Norde, W.; Van Der Mei, H.C.; Sjollema, J. Residence-time dependent cell wall deformation of different Staphylococcus aureus strains on gold measured using surface-enhanced-fluorescence. Soft Matter 2014, 10, 7638–7646. [Google Scholar] [CrossRef]
- Perozo, E.; Kloda, A.; Cortes, D.M.; Martinac, B. Physical principles underlying the transduction of bilayer defor-mation forces during mechanosensitive channel gating. Nat. Struct. Biol. 2002, 9, 696–703. [Google Scholar] [CrossRef]
- Baro, A.M.; Reifenberger, R.G. Atomic Force Microscopy in Liquid: Biological Applications; Wiley-VCH: Weinheim, Germany, 2012. [Google Scholar]
- Beaussart, A.; El-Kirat-Chatel, S.; Sullan, R.M.A.; Alsteens, D.; Herman, P.; Derclaye, S.; Dufrêne, Y.F. Quantifying the forces guiding microbial cell adhesion using single-cell force spectroscopy. Nat. Protoc. 2014, 9, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.W.; He, Y.; Ren, Y.; Zerdoum, A.; Libera, M.R.; Sharma, P.K.; Van Winkelhoff, A.-J.; Neut, D.; Stoodley, P.; Van Der Mei, H.C.; et al. Viscoelasticity of biofilms and their recalcitrance to mechanical and chemical challenges. FEMS Microbiol. Rev. 2015, 39, 234–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.; Müller, T.; Meyer, R.L. Single-Cell Force Spectroscopy of Bacteria Enabled by Naturally Derived Proteins. Langmuir 2014, 30, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Tarafdar, A.; Lee, J.-U.; Jeong, J.-E.; Lee, H.; Jung, Y.; Oh, H.-B.; Woo, H.Y.; Kwon, J.-H. Biofilm development of Bacillus siamensis ATKU1 on pristine short chain low-density polyethylene: A case study on microbe-microplastics interaction. J. Hazard. Mater. 2020, 124516. [Google Scholar] [CrossRef] [PubMed]
- Boyd, C.D.; Smith, T.J.; El-Kirat-Chatel, S.; Newell, P.D.; Dufrêne, Y.F.; O’Toole, G.A. Structural Features of the Pseudomonas fluorescens Biofilm Adhesin LapA Required for LapG-Dependent Cleavage, Biofilm Formation, and Cell Surface Localization. J. Bacteriol. 2014, 196, 2775–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, V.; Znaidi, S.; Walker, L.A.; Martin-Yken, H.; Dague, E.; Legrand, M.; Lee, K.; Chauvel, M.; Firon, A.; Rossignol, T.; et al. Targeted Changes of the Cell Wall Proteome Influence Candida albicans Ability to Form Single- and Multi-strain Biofilms. PLOS Pathog. 2014, 10, e1004542. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Cui, Y.; Oh, Y.J.; Park, J.R.; Jo, W.; Cho, Y.-H.; Park, S. Studying the effect of alginate overproduction on Pseudomonas aeruginosa biofilm by atomic force microscopy. J. Nanosci. Nanotechnol. 2011, 11, 5676–5681. [Google Scholar] [CrossRef]
- Ovchinnikova, E.S.; Krom, B.P.; Harapanahalli, A.K.; Busscher, H.J.; Van Der Mei, H.C. Surface Thermodynamic and Adhesion Force Evaluation of the Role of Chitin-Binding Protein in the Physical Interaction between Pseudomonas aeruginosaand Candida albicans. Langmuir 2013, 29, 4823–4829. [Google Scholar] [CrossRef]
- Potthoff, E.; Ossola, D.; Zambelli, T.; Vorholt, J.A. Bacterial adhesion force quantification by fluidic force microscopy. Nanoscale 2015, 7, 4070–4079. [Google Scholar] [CrossRef]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Al-Waili, N.; Sharma, D.; Nuru, A.; Al-Attal, Y. Effect of Jujube Honey on Candida albicans Growth and Biofilm Formation. Arch. Med. Res. 2013, 44, 352–360. [Google Scholar] [CrossRef]
- Chatterjee, S.; Biswas, N.; Datta, A.; Dey, R.; Maiti, P. Atomic force microscopy in biofilm study. Microscopy (Oxf. Engl.) 2014, 63, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danin, P.-. Éric; Girou, E.; Legrand, P.; Louis, B.; Fodil, R.; Christov, C.; Devaquet, J.; Isabey, D.; Brochard, L. Description and Microbiology of Endotracheal Tube Biofilm in Mechanically Ventilated Subjects. Respir. Care 2014, 60, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangalappalli-Illathu, A.K.; Vidovicć, S.; Korber, D. Differential Adaptive Response and Survival of Salmonella enterica Serovar Enteritidis Planktonic and Biofilm Cells Exposed to Benzalkonium Chloride. Antimicrob. Agents Chemother. 2008, 52, 3669–3680. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, K.; Obika, H.; Utsumi, A.; Ooie, T.; Yano, T. In vitro laser ablation of laboratory developed biofilms using an Nd:YAG laser of 532 nm wavelength. Biotechnol. Bioeng. 2004, 86, 729–736. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.; Hu, Y.; Chi, Q.; Mortensen, N.P.; Qu, D.; Molin, S.; Ulstrup, J. Organic compounds inhibiting S. epidermidis adhesion and biofilm formation. Ultramicroscopy 2009, 109, 881–888. [Google Scholar] [CrossRef]
- Sharma, S.; Cross, S.E.; Hsueh, C.; Wali, R.P.; Stieg, A.Z.; Gimzewski, J.K. Nanocharacterization in Dentistry. Int. J. Mol. Sci. 2010, 11, 2523–2545. [Google Scholar] [CrossRef]
- Bossù, M.; Selan, L.; Artini, M.; Relucenti, M.; Familiari, G.; Papa, R.; Vrenna, G.; Spigaglia, P.; Barbanti, F.; Salucci, A.; et al. Characterization of Scardovia wiggsiae Biofilm by Original Scanning Electron Microscopy Protocol. Microorganisms 2020, 8, 807. [Google Scholar] [CrossRef]
- Hung, C.; Zhou, Y.; Pinkner, J.S.; Dodson, K.W.; Crowley, J.R.; Heuser, J.; Chapman, M.R.; Hadjifrangiskou, M.; Henderson, J.P.; Hultgren, S.J. Escherichia coli Biofilms Have an Organized and Complex Extracellular Matrix Structure. mBio 2013, 4, e00645-13. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, D.A.F.; Bañobre-López, M.; Espiña, B.; Rivas, J.; Azeredo, J. Effect of magnetic hyperthermia on the structure of biofilm and cellular viability of a food spoilage bacterium. Biofouling 2013, 29, 1225–1232. [Google Scholar] [CrossRef]
- Di Bonaventura, G.; Piccolomini, R.; Paludi, D.; D’Orio, V.; Vergara, A.; Conter, M.; Ianieri, A. Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 2008, 104, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, G.; Pompilio, A.; Picciani, C.; Iezzi, M.; D’Antonio, D.; Piccolomini, R. Biofilm Formation by the Emerging Fungal Pathogen Trichosporon asahii: Development, Architecture, and Antifungal Resistance. Antimicrob. Agents Chemother. 2006, 50, 3269–3276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Danishuddin, M.; Khan, A.U. Inhibitory effect of zingiber officinale towards Streptococcus mutans virulence and caries development: In vitro and in vivo studies. BMC Microbiol. 2015, 15, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.W.; Li, Y.F.; Gao, R.; Liu, K.Y.; Zhang, H.Q.; Fu, Q.H.; Guo, G.; Zou, Q.M.; Qing, S.L. Inhibited biofilm formation and improved antibacterial activity of a novel nanoemulsion against cariogenic Streptococcus mutans in vitro and in vivo. Int. J. Nanomed. 2015, 10, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Orsinger-Jacobsen, S.J.; Patel, S.S.; Vellozzi, E.M.; Gialanella, P.; Nimrichter, L.; Miranda, K.; Martinez, L.R. Use of a stainless steel washer platform to study Acinetobacter baumannii adhesion and biofilm formation on abiotic surfaces. Microbiology (Read. Engl.) 2013, 159, 2594–2604. [Google Scholar] [CrossRef] [Green Version]
- Samaranayake, Y.H.; Cheung, B.P.K.; Yau, J.Y.Y.; Yeung, S.K.W.; Samaranayake, Y.H. Human Serum Promotes Candida albicans Biofilm Growth and Virulence Gene Expression on Silicone Biomaterial. PLoS ONE 2013, 8, e62902. [Google Scholar] [CrossRef] [Green Version]
- Van Laar, T.A.; Chen, T.; You, T.; Leung, K.P. Sublethal Concentrations of Carbapenems Alter Cell Morphology and Genomic Expression of Klebsiella pneumoniae Biofilms. Antimicrob. Agents Chemother. 2015, 59, 1707–1717. [Google Scholar] [CrossRef] [Green Version]
- Bressan, E.; Tessarolo, F.; Sbricoli, L.; Caola, I.; Nollo, G.; Di Fiore, A. Effect of Chlorhexidine in Preventing Plaque Biofilm on Healing Abutment: A crossover controlled study. Implant. Dent. 2014, 23, 64–68. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Caola, I.; Nollo, G.; Cavallo, M.; Rinaldi, M.; Fracchia, L. Inhibition of Candida albicans adhesion on medical-grade silicone by a Lactobacillus -derived biosurfactant. J. Appl. Microbiol. 2015, 118, 1116–1125. [Google Scholar] [CrossRef]
- Garcez, A.S.; Núñez, S.C.; Azambuja, N., Jr.; Fregnani, E.R.; Rodriguez, H.M.; Hamblin, M.R.; Suzuki, H.; Ribeiro, M.S. Effects of Photodynamic Therapy on Gram-Positive and Gram-Negative Bacterial Biofilms by Bioluminescence Imaging and Scanning Electron Microscopic Analysis. Photomed. Laser Surg. 2013, 31, 519–525. [Google Scholar] [CrossRef]
- Nishitani, K.; Sutipornpalangkul, W.; Bentley, K.L.D.M.; Varrone, J.J.; Bello-Irizarry, S.N.; Ito, H.; Matsuda, S.; Kates, S.L.; Daiss, J.L.; Schwarz, E.M. Quantifying the natural history of biofilm formation in vivo during the establishment of chronic implant-associated Staphylococcus aureus osteomyelitis in mice to identify critical pathogen and host factors. J. Orthop. Res. 2015, 33, 1311–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhede, M.; Qvortrup, K.; Liebrechts, R.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Combination of microscopic techniques reveals a comprehensive visual impression of biofilm structure and composition. FEMS Immunol. Med. Microbiol. 2012, 65, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Timp, W.; Matsudaira, P. Chapter 14 Electron Microscopy of Hydrated Samples. Methods Cell Biol. 2008, 89, 391–407. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, J.C.; Téran, F.C.; Oliveira, R.A.; Nour, E.A.A.; Montenegro, M.A.P.; Campos, J.R.; Vazoller, R.F. Comparison of hexamethyldisilazane and critical point drying treatments for SEM analysis of anaerobic biofilms and granular sludge. QJM Int. J. Med. 2003, 52, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Karcz, J.; Bernas, T.; Nowak, A.; Talik, E.; Woznica, A. Application of lyophilization to prepare the nitrifying bacterial biofilm for imaging with scanning electron microscopy. Scanning 2011, 34, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Bossù, M.; Matassa, R.; Relucenti, M.; Iaculli, F.; Salucci, A.; Di Giorgio, G.; Familiari, G.; Polimeni, A.; Di Carlo, S. Morpho-Chemical Observations of Human Deciduous Teeth Enamel in Response to Biomimetic Toothpastes Treatment. Materials 2020, 13, 1803. [Google Scholar] [CrossRef] [Green Version]
- Bossù, M.; Saccucci, M.; Salucci, A.; Di Giorgio, G.; Bruni, E.; Uccelletti, D.; Sarto, M.S.; Familiari, G.; Relucenti, M.; Polimeni, A. Enamel remineralization and repair results of Biomimetic Hydroxyapatite toothpaste on deciduous teeth: An effective option to fluoride toothpaste. J. Nanobiotechnol. 2019, 17, 17. [Google Scholar] [CrossRef]
- Redler, A.; Miglietta, S.; Monaco, E.; Matassa, R.; Relucenti, M.; Daggett, M.; Ferretti, A.; Familiari, G. Ultrastructural Assessment of the Anterolateral Ligament. Orthop. J. Sports Med. 2019, 7, 2325967119887920. [Google Scholar] [CrossRef] [Green Version]
- Priester, J.H.; Horst, A.M.; Van De Werfhorst, L.C.; Saleta, J.L.; Mertes, L.A.; Holden, P.A. Enhanced visualization of microbial biofilms by staining and environmental scanning electron microscopy. J. Microbiol. Methods 2007, 68, 577–587. [Google Scholar] [CrossRef]
- Weber, K.; Delben, J.; Bromage, T.G.; Duarte, S. Comparison of SEM and VPSEM imaging techniques with respect toStreptococcus mutansbiofilm topography. FEMS Microbiol. Lett. 2013, 350, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Ishii, S.; Koki, J.; Unno, H.; Hori, K. Two Morphological Types of Cell Appendages on a Strongly Adhesive Bacterium, Acinetobacter sp. Strain Tol 5. Appl. Environ. Microbiol. 2004, 70, 5026–5029. [Google Scholar] [CrossRef] [Green Version]
- Weimer, P.J.; Price, N.P.J.; Kroukamp, O.; Joubert, L.-M.; Wolfaardt, G.M.; Van Zyl, W.H. Studies of the Extracellular Glycocalyx of the Anaerobic Cellulolytic Bacterium Ruminococcus albus 7. Appl. Environ. Microbiol. 2006, 72, 7559–7566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, L.-M.; Ferreira, J.A.; A Stevens, D.; Nazik, H.; Cegelski, L. Visualization of Aspergillus fumigatus biofilms with Scanning Electron Microscopy and Variable Pressure-Scanning Electron Microscopy: A comparison of processing techniques. J. Microbiol. Methods 2017, 132, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Bozzola, J.J.; Russell, L.D. Electron. Microscopy: Principles and Techniques for Biologists; Jones & Bartlett Learning: Burlington, MA, USA, 1999. [Google Scholar]
- Fassel, T.A.; Edmiston, C.E., Jr. Ruthenium Red and the Bacterial Glycocalyx. Biotech. Histochem. 1999, 74, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlandsen, S.L.; Kristich, C.J.; Dunny, G.M.; Wells, C.L. High-resolution Visualization of the Microbial Glycocalyx with Low-voltage Scanning Electron Microscopy: Dependence on Cationic Dyes. J. Histochem. Cytochem. 2004, 52, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Reichhardt, C.; Ferreira, J.A.G.; Joubert, L.-M.; Clemons, K.V.; Stevens, D.A.; Cegelski, L. Analysis of the Aspergillus fumigatus Biofilm Extracellular Matrix by Solid-State Nuclear Magnetic Resonance Spectroscopy. Eukaryot. Cell 2015, 14, 1064–1072. [Google Scholar] [CrossRef] [Green Version]
- Trent, J.S.; Scheinbeim, J.I.; Couchman, P.R. Ruthenium tetraoxide staining of polymers for electron micros-copy. Macromolecules 1983, 16, 589–598. [Google Scholar] [CrossRef]
- Behnke, O.; Zelander, T. Preservation of intercellular substances by the cationic dye alcian blue in preparative procedures for electron microscopy. J. Ultrastruct. Res. 1970, 31, 424–438. [Google Scholar] [CrossRef]
- Fassel, T.A.; Mozdziak, P.E.; Sanger, J.R.; Edmiston, C.E. Paraformaldehyde effect on ruthenium red and ly-sine preservation and staining of the staphylococcal glycocalyx. Microsc. Res. Tech. 1997, 36, 422–427. [Google Scholar] [CrossRef]
- Forte, M.; Bianchi, F.; Cotugno, M.; Marchitti, S.; De Falco, E.; Raffa, S.; Stanzione, R.; Di Nonno, F.; Chimenti, I.; Palmerio, S.; et al. Pharmacological restoration of autophagy reduces hypertension-related stroke occurrence. Autophagy 2020, 16, 1468–1481. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, A.; Serpe, C.; Chece, G.; Nigro, V.; Sarra, A.; Ruzicka, B.; Relucenti, M.; Familiari, G.; Ruocco, G.; Pascucci, G.R.; et al. Microglia-Derived Microvesicles Affect Microglia Phenotype in Glioma. Front. Cell. Neurosci. 2019, 13, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardoni, M.; Della Valle, E.; Liberti, M.; Relucenti, M.; Casadei, M.A.; Paolicelli, P.; Apollonio, F.; Petralito, S. Can Pulsed Electromagnetic Fields Trigger On-Demand Drug Release from High-Tm Magnetoliposomes? Nanomaterials 2018, 8, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayat, M.A. Stains and Cytochemical Methods; Hayat, M.A., Ed.; Plenum Press: New York, NY, USA, 1993; Volume 1, pp. 336–347. [Google Scholar]
- Tapia, J.C.; Kasthuri, N.; Hayworth, K.J.; Schalek, R.; Lichtman, J.W.; Smith, S.J.; Buchanan, J. High-contrast en bloc staining of neuronal tissue for field emission scanning electron microscopy. Nat. Protoc. 2012, 7, 193–206. [Google Scholar] [CrossRef]
- Murakami, T.; Iida, N.; Taguchi, T.; Ohtani, O.; Kikuta, A.; Ohtsuka, A.; Itoshima, T. Conductive staining of biological specimens for scanning electron microscopy with special reference to ligand-mediated osmium impregnation. Scanning Electron. Microsc. 1983, 235–246. [Google Scholar]
- Murakami, T. A revised tannin-osmium method for non-coated SEM specimens. Arch. Histol. Jpn. 1974, 36, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T. A metal impregnation method of biological specimens for SEM. Arch. Histol. Jpn. 1973, 35, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T. Puncture perfusion of small tissue pieces for SEM. Arch. Histol. Jpn. 1976, 39, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Yamamoto, K.; Itoshima, T.; Irino, S. Modified tannin osmium conductive staining method for non-coated SEM specimens. Its application to microdissection SEM of the spleen. Arch. Histol. Jpn. 1977, 40, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Gunji, T.; Wakita, M.; Kobayashi, S. Conductive staining in SEM with especial reference to tissue transparency. Scanning 1980, 3, 227–232. [Google Scholar] [CrossRef]
- Arimoto, S.; Sugimura, M.; Kageyama, H.; Torimoto, T.; Kuwabata, S. Development of new techniques for scanning electron microscope observation using ionic liquid. Electrochim. Acta 2008, 53, 6228–6234. [Google Scholar] [CrossRef]
- Tsuda, T.; Nemoto, N.; Kawakami, K.; Mochizuki, E.; Kishida, S.; Tajiri, T.; Kushibiki, T.; Kuwabata, S. SEM Observation of Wet Biological Specimens Pretreated with Room-Temperature Ionic Liquid. ChemBioChem 2011, 12, 2547–2550. [Google Scholar] [CrossRef] [PubMed]
- Brodusch, N.; Waters, K.; Demers, H.; Gauvin, R. Ionic liquid-based observation technique for nonconductive materials in the scanning electron microscope: Application to the characterization of a rare earth ore. Microsc. Res. Tech. 2014, 77, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Imashuku, S.; Kawakami, T.; Zea, L.; Kawai, J. Possibility of Scanning Electron Microscope Observation and Energy Dispersive X-Ray Analysis in Microscale Region of Insulating Samples Using Diluted Ionic Liquid. Microsc. Microanal. 2012, 18, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torto, F.L.; Relucenti, M.; Familiari, G.; Vaia, N.; Casella, D.; Matassa, R.; Miglietta, S.; Marinozzi, F.; Bini, F.; Fratoddi, I.; et al. The Effect of Postmastectomy Radiation Therapy on Breast Implants: Material Analysis on Silicone and Polyurethane Prosthesis. Ann. Plast. Surg. 2018, 81, 228–234. [Google Scholar] [CrossRef]
- Relucenti, M.; Miglietta, S.; Bove, G.; Donfrancesco, O.; Battaglione, E.; Familiari, P.; Barbaranelli, C.; Covelli, E.; Barbara, M.; Familiari, G. SEM BSE 3D Image Analysis of Human Incus Bone Affected by Cholesteatoma Ascribes to Osteoclasts the Bone Erosion and VpSEM dEDX Analysis Reveals New Bone Formation. Scanning 2020, 2020, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Relucenti, M.; Miglietta, S.; Covelli, E.; Familiari, P.; Battaglione, E.; Familiari, G.; Barbara, M. Ciliated cell observation by SEM on the surface of human incudo-malleolar-joint articular cartilage: Are they a new chondrocyte phenotype? Acta Oto-Laryngol. 2019, 139, 439–443. [Google Scholar] [CrossRef]
- Fulcher, T.P.; Dart, J.K.; McLaughlin-Borlace, L.; Howes, R.; Matheson, M.; Cree, I. Demonstration of biofilm in infectious crystalline keratopathy using ruthenium red and electron microscopy. Ophthalmology 2001, 108, 1088–1092. [Google Scholar] [CrossRef]
- Luft, J.H. Electron microscopy of cell extraneous coats as revealed by ruthenium red staining. J. Cell Biol. 1964, 23, 54A–55A. [Google Scholar]
- Fassel, T.A.; Schaller, M.J.; Remsen, C.C. Comparison of Alcian blue and ruthenium red effects on preservation of outer envelope ultrastructure in methanotrophic bacteria. Microsc. Res. Tech. 1992, 20, 87–94. [Google Scholar] [CrossRef]
- Bleck, C.; Merz, A.; Gutierrez, M.G.; Walther, P.; Dubochet, J.; Zuber, B.; Griffiths, G. Comparison of different methods for thin section EM analysis of Mycobacterium smegmatis. J. Microsc. 2009, 237, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Hrubanova, K.; Nebesarova, J.; Růžička, F.; Krzyzanek, V. The innovation of cryo-SEM freeze-fracturing methodology demonstrated on high pressure frozen biofilm. Micron 2018, 110, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, H.; Koizumi, M.; Ogawa, K.; Kitamura, S.; Konyuba, Y.; Watanabe, Y.; Ohbayashi, N.; Fukuda, M.; Suga, M.; Sato, C. Atmospheric scanning electron microscope system with an open sample chamber: Configuration and applications. Ultramicroscopy 2014, 147, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Y.; Ebihara, T.; Nishiyama, H.; Suga, M.; Sato, C. Immuno EM–OM correlative microscopy in solution by atmospheric scanning electron microscopy (ASEM). J. Struct. Biol. 2012, 180, 259–270. [Google Scholar] [CrossRef]
- Hirano, K.; Kinoshita, T.; Uemura, T.; Motohashi, H.; Watanabe, Y.; Ebihara, T.; Nishiyama, H.; Sato, M.; Suga, M.; Maruyama, Y.; et al. Electron microscopy of primary cell cultures in solution and correlative optical microscopy using ASEM. Ultramicroscopy 2014, 143, 52–66. [Google Scholar] [CrossRef]
- Sato, C.; Manaka, S.; Nakane, D.; Nishiyama, H.; Suga, M.; Nishizaka, T.; Miyata, M.; Maruyama, Y. Rapid imaging of mycoplasma in solution using Atmospheric Scanning Electron Microscopy (ASEM). Biochem. Biophys. Res. Commun. 2012, 417, 1213–1218. [Google Scholar] [CrossRef]
- Sugimoto, S.; Okuda, K.-I.; Miyakawa, R.; Sato, M.; Arita-Morioka, K.-I.; Chiba, A.; Yamanaka, K.; Ogura, T.; Mizunoe, Y.; Sato, C. Imaging of bacterial multicellular behaviour in biofilms in liquid by atmospheric scanning electron microscopy. Sci. Rep. 2016, 6, 25889. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Valdevit, A.; Chou, T.-M.; Libera, M.R. Substrate effects on cell-envelope deformation during early-stage Staphylococcus aureus biofilm formation. Soft Matter 2017, 13, 2967–2976. [Google Scholar] [CrossRef]
- Cao, Y.; Su, B.; Chinnaraj, S.; Jana, S.; Bowen, L.; Charlton, S.; Duan, P.; Jakubovics, N.S.; Chen, J. Nanostructured titanium surfaces exhibit recalcitrance towards Staphylococcus epidermidis biofilm formation. Sci. Rep. 2018, 8, 1071. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Microscopy | CLSM | AFM | |
|---|---|---|---|
| Pros | Simple protocols Cheap and easy to perform Large investigation area | Allows single cell visualization and 3-D imaging | Nondestructive technique that works under physiological-like conditions, allowing living biofilms qualitative and quantitative assessment with few treatments, sample 3-D structure reconstruction at nanometer scale. |
| Cons | Low resolution and magnification power, need for sample staining, gross morphological differentiation, finest details not visible | Use of fluorophores, limited number of reporter molecules, intrinsic biofilm fluorescence can interfere with probes fluorescence | Small scan area (max 150 × 150 µm), no image of bacterial cells sidewalls, possible surface damage during imaging due to tip interactions. |
| Applications | Visualization of biofilm formation and quantitative assessment of its biomass | Assessment of biofilm structural parameters, Biofilm 3D structure, identification and localization of living and death cells | Quantitative biofilm analysis, determination of adhesion forces, biofilm topography, in situ imaging. |
| Protocols | |||
|---|---|---|---|
| Steps | Conventional SEM | VP-SEM | OsO4-RR-TA-IL |
| Fixation | Glutaraldehyde 2.5% in PB 0.1 M pH 7.4 at least 48 h | ||
| Washing | 10 min × 2 times in PB 0.1 M pH 7.4 | ||
| Post-fixation | OsO4 2% 1 h | OsO4 2% in 1 h | OsO4 2% + RR 0.2% 1:1 solution, 1 h |
| Washing | 10 min × 2 times in dH20 | ||
| Impregnation | None | None | Tannic Acid 1% in d H20 30 min |
| Washing | - | - | 10 min × 2 times in dH20 |
| Dehydration | Ascending ethanol series | None | None |
| Drying | Ascending HMDS 1 series | None | None |
| Pt Sputter coating | 15 mA, 2 min | None | Replaced by IL |
| Operating conditions | 15–20 kV, high vacuum | 5–10 kV 30 Pa | 15–20 kV, high vacuum |
| Protocols | |||
|---|---|---|---|
| Parameters | Conventional SEM | VP-SEM | OsO4-RR-TA-IL |
| Procedure time | 2 days | 1 h and 30 min | 2 h and 10 min |
| Sample loss | Steps produce sample loss of about 60% | about 20% | about 20% |
| Dehydration and drying | yes | None | None |
| Pt Sputter coating | yes | None | Replaced by IL |
| Resistance in vacuum | Excellent, it is possible to observe for hours | Good for 1 h | Excellent, it is possible to observe for hours |
| Operating conditions | 15–20 kV, high vacuum | 5–10 kV 30 Pa | 15–20 kV, high vacuum |
| Image magnification | Good up to 40 k | Good up to 10 k | Good up to 30 k |
| Image quality | Excellent up to 30 k | Good up to 8 k | Excellent up to 30 k |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relucenti, M.; Familiari, G.; Donfrancesco, O.; Taurino, M.; Li, X.; Chen, R.; Artini, M.; Papa, R.; Selan, L. Microscopy Methods for Biofilm Imaging: Focus on SEM and VP-SEM Pros and Cons. Biology 2021, 10, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010051
Relucenti M, Familiari G, Donfrancesco O, Taurino M, Li X, Chen R, Artini M, Papa R, Selan L. Microscopy Methods for Biofilm Imaging: Focus on SEM and VP-SEM Pros and Cons. Biology. 2021; 10(1):51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010051
Chicago/Turabian StyleRelucenti, Michela, Giuseppe Familiari, Orlando Donfrancesco, Maurizio Taurino, Xiaobo Li, Rui Chen, Marco Artini, Rosanna Papa, and Laura Selan. 2021. "Microscopy Methods for Biofilm Imaging: Focus on SEM and VP-SEM Pros and Cons" Biology 10, no. 1: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010051