Alterations in Glucose Metabolism Due to Decreased Expression of Heterogeneous Nuclear Ribonucleoprotein M in Pancreatic Ductal Adenocarcinoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Human Tissue cDNA, and Cell Line
2.2. Polymerase Chain Reaction (PCR)
2.3. Cell Cultures
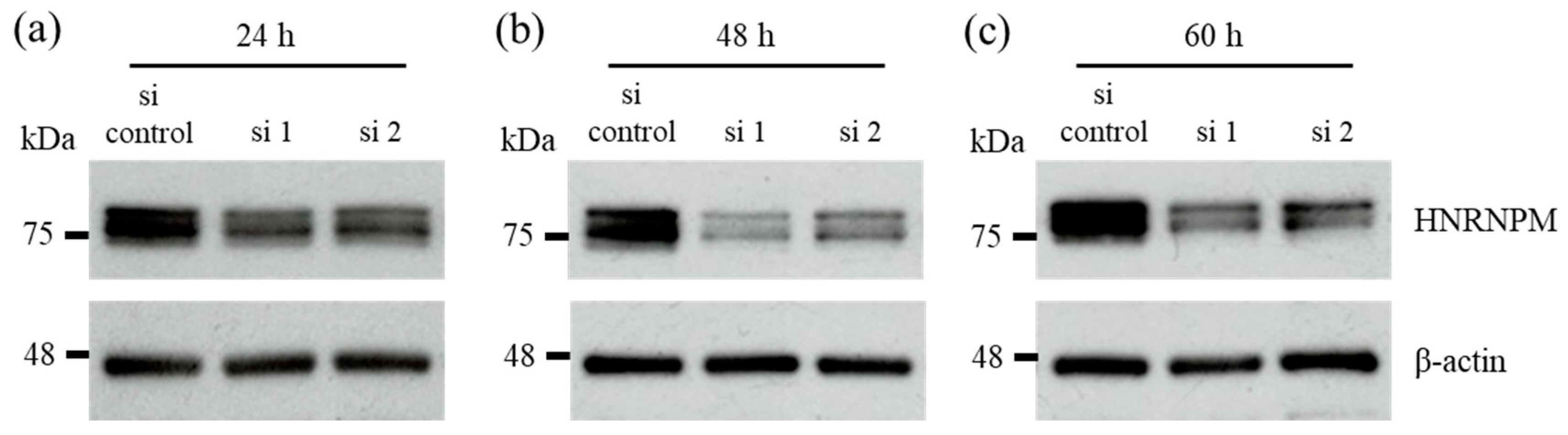
2.4. HNRNPM Knockdown
2.5. Preparation of Cell Lysates and Western Blotting Analysis
2.6. Cell Proliferation Assay
2.7. Glucose and Lactate Concentration in the Medium
2.8. Glucose Uptake
2.9. Statistical Analysis
3. Results
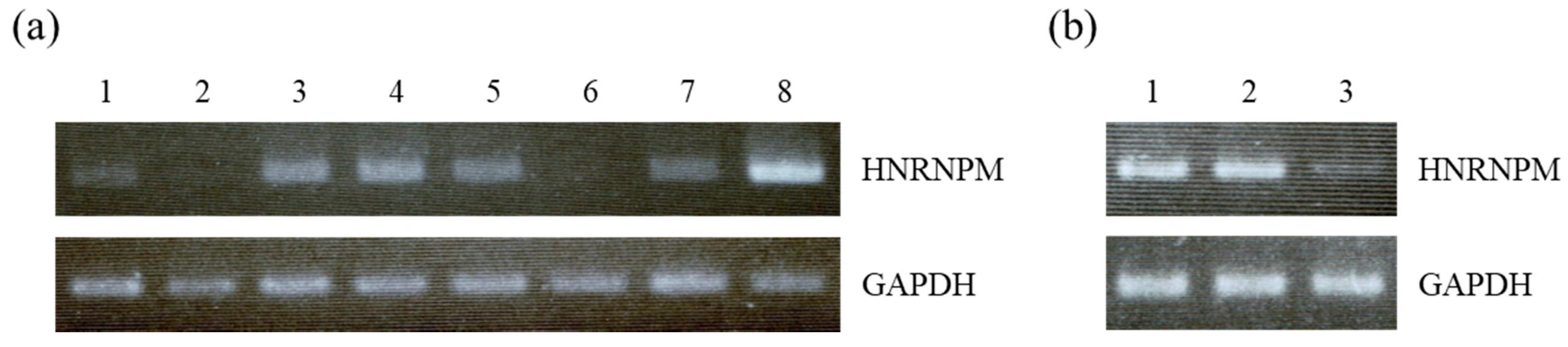
3.1. HNRNPM Is Highly Expressed in Pancreatic Tissues but Expression Is Reduced in PDA Tissues
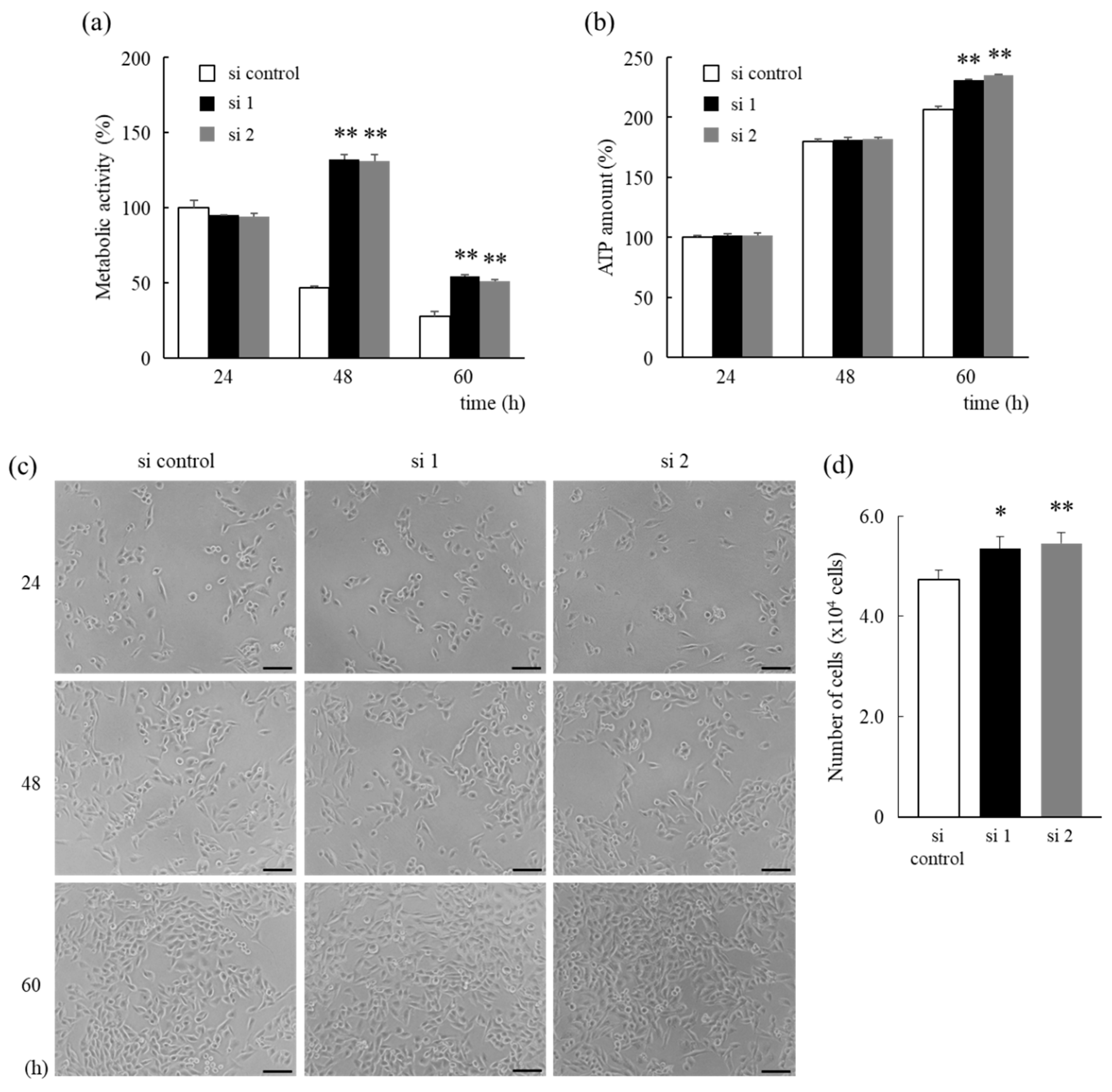
3.2. HNRNPM Knockdown Prolongs Cell Survival of MIA PaCa-2 Cells under Low-Glucose Conditions
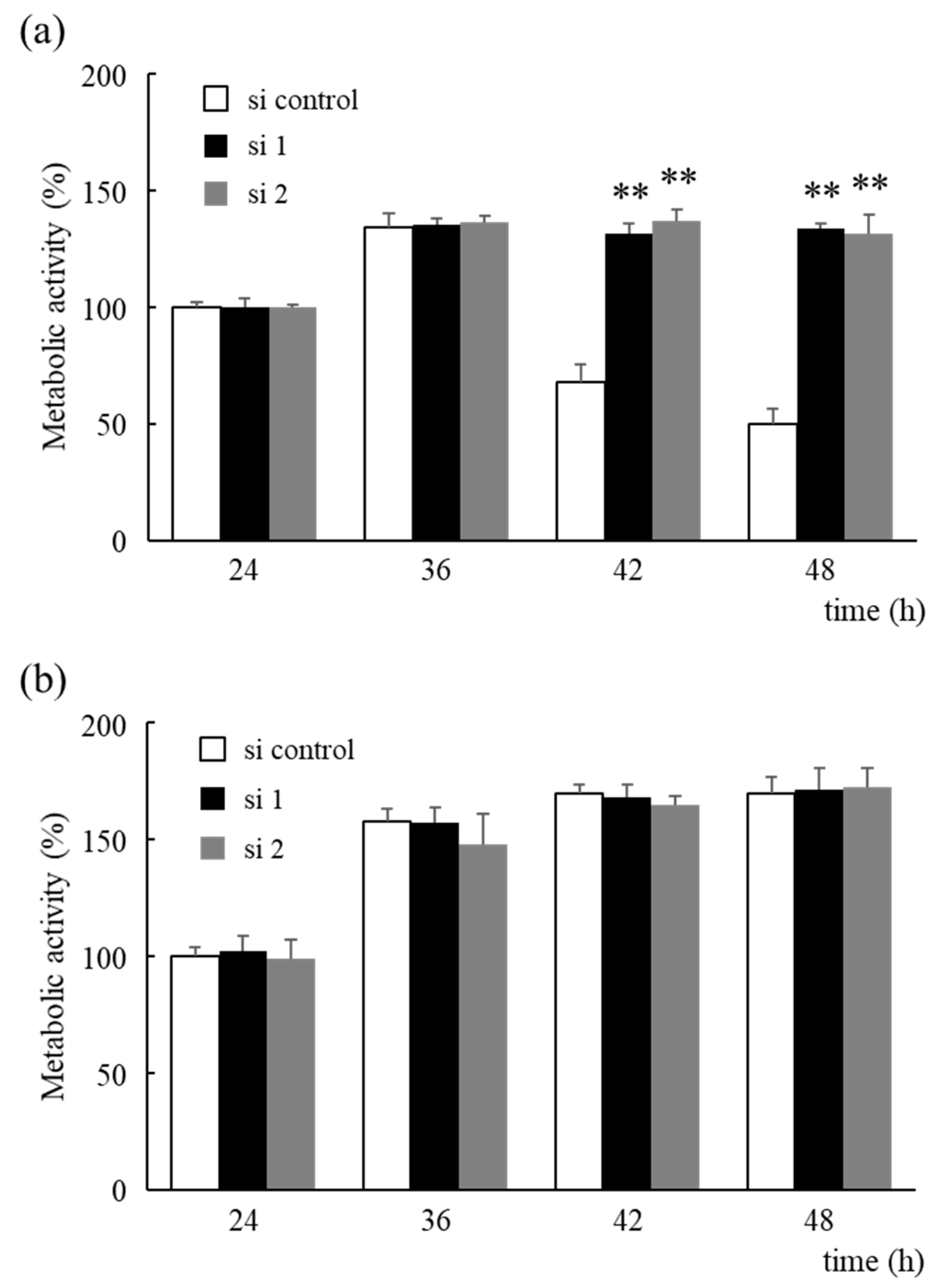
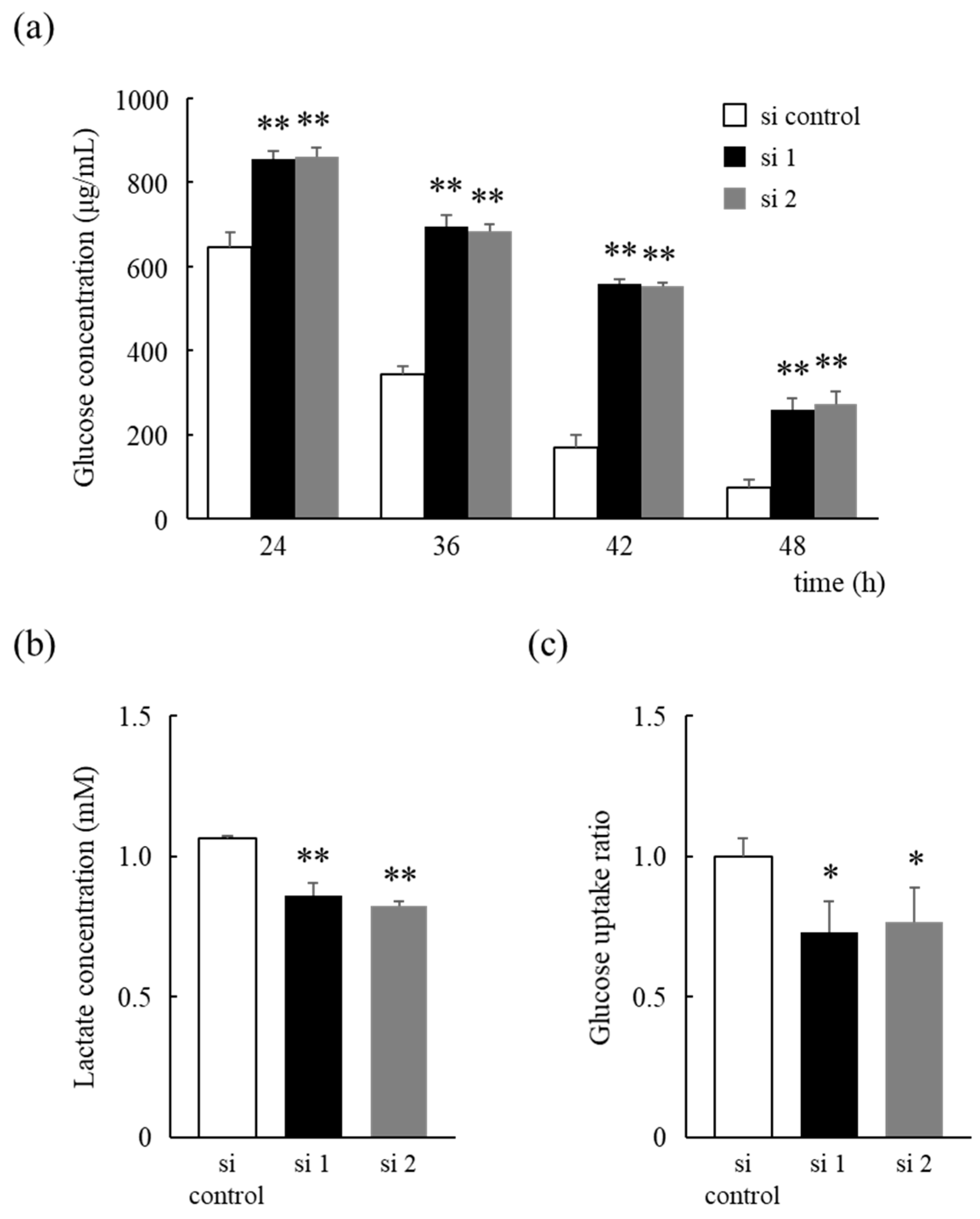
3.3. HNRNPM Knockdown Reduces Glucose Consumption of MIA PaCa-2 Cells by Altering Glucose Metabolism
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cancer Facts & Figures. 2020. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2020.html (accessed on 11 November 2020).
- Goral, V. Pancreatic cancer: Pathogenesis and diagnosis. Asian Pac. J. Cancer Prev. 2015, 16, 5619–5624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaziri-Gohar, A.; Zarei, M.; Brody, J.R.; Winter, J.M. Metabolic dependencies in pancreatic cancer. Front Oncol. 2018, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.R.; O’Reilly, E.M. New treatment strategies for metastatic pancreatic ductal adenocarcinoma. Drugs 2020, 80, 647–669. [Google Scholar] [CrossRef] [PubMed]
- Neesse, A.; Michl, P.; Frese, K.K.; Feig, C.; Cook, N.; Jacobetz, M.A.; Lolkema, M.P.; Buchholz, M.; Olive, K.P.; Gress, T.M.; et al. Stromal biology and therapy in pancreatic cancer. Gut 2011, 60, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Izuishi, K.; Kato, K.; Ogura, T.; Kinoshita, T.; Esumi, H. Remarkable tolerance of tumor cells to nutrient deprivation: Possible new biochemical target for cancer therapy. Cancer Res. 2000, 60, 6201–6207. [Google Scholar] [PubMed]
- Datar, K.V.; Dreyfuss, G.; Swanson, M.S. The human hnRNP M proteins: Identification of a methionine/arginine-rich repeat motif in ribonucleoproteins. Nucleic Acids Res. 1993, 21, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Passacantilli, I.; Frisone, P.; De Paola, E.; Fidaleo, M.; Paronetto, M.P. hnRNPM guides an alternative splicing program in response to inhibition of the PI3K/AKT/mTOR pathway in ewing sarcoma cells. Nucleic Acids Res. 2017, 45, 12270–12284. [Google Scholar] [CrossRef]
- Bjersand, K.; Seidal, T.; Sundström-Poromaa, I.; Åkerud, H.; Skirnisdottir, I. The clinical and prognostic correlation of HRNPM and SLC1A5 in pathogenesis and prognosis in epithelial ovarian cancer. PLoS ONE 2017, 12, e0179363. [Google Scholar] [CrossRef]
- Chen, T.M.; Lai, M.C.; Li, Y.H.; Chan, Y.L.; Wu, C.H.; Wang, Y.M.; Chien, C.W.; Huang, S.Y.; Sun, H.S.; Tsai, S.J. hnRNPM induces translation switch under hypoxia to promote colon cancer development. EBioMedicine 2019, 41, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Palombo, R.; Verdile, V.; Paronetto, M.P. Poison-exon inclusion in DHX9 reduces its expression and sensitizes ewing sarcoma cells to chemotherapeutic treatment. Cells 2020, 9, 328. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhang, J.; Duan, L.; Zhang, Y.; Li, C.; Liu, D.; Ouyang, C.; Lu, F.; Liu, X. Identification of HnRNP M as a novel biomarker for colorectal carcinoma by quantitative proteomics. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Gao, X.D.; Lee, J.H.; Huang, H.; Tan, H.; Ahn, J.; Reinke, L.M.; Peter, M.E.; Feng, Y.; Gius, D.; et al. Cell type-restricted activity of hnRNPM promotes breast cancer metastasis via regulating alternative splicing. Genes Dev. 2014, 28, 1191–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Liu, T.; Zhu, D.; Dong, X.; Liu, F.; Liang, X.; Chen, C.; Shao, B.; Wang, M.; Wang, Y. HnRNPM and CD44s expression affects tumor aggressiveness and predicts poor prognosis in breast cancer with axillary lymph node metastases. Genes Chromosomes Cancer 2017, 56, 598–607. [Google Scholar] [CrossRef]
- Yang, W.H.; Ding, M.J.; Cui, G.Z.; Yang, M.; Dai, D.L. Heterogeneous nuclear ribonucleoprotein M promotes the progression of breast cancer by regulating the axin/β-catenin signaling pathway. Biomed. Pharm. 2018, 105, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Cao, J.L.; Xie, H.Y.; Sun, R.; Yang, L.F.; Shao, Z.M.; Li, D.Q. Cancer-associated MORC2-mutant M276I regulates an hnRNPM-mediated CD44 splicing switch to promote invasion and metastasis in triple-negative breast cancer. Cancer Res. 2018, 78, 5780–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; An, Z.; Zhang, C.; Wang, Z.; Wang, X.; Liu, Y.; Du, E.; Liu, R.; Zhang, Z.; Xu, Y. HnRNPM is a potential mediator of YY1 which promotes EMT in prostate cancer cells. Prostate 2019, 79, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Daley, D.T.; Luscombe, N.M.; Berman, H.M.; Thornton, J.M. Protein-RNA interactions: A structural analysis. Nucleic Acids Res. 2001, 29, 943–954. [Google Scholar] [CrossRef]
- Kim, H.; Jeong, E.; Lee, S.W.; Han, K. Computational analysis of hydrogen bonds in protein-RNA complexes for interaction patterns. FEBS Lett. 2003, 552, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, D.; Delsaux, N.; Charloteaux, B.; Thomas, A.; Brasseur, R. Protein-nucleic acid recognition: Statistical analysis of atomic interactions and influence of DNA structure. Proteins 2005, 61, 258–271. [Google Scholar] [CrossRef]
- Ellis, J.J.; Broom, M.; Jones, S. Protein-RNA interactions: Structural analysis and functional classes. Proteins 2007, 66, 903–911. [Google Scholar] [CrossRef]
- Ciriello, G.; Gallina, C.; Guerra, C. Analysis of interactions between ribosomal proteins and RNA structural motifs. BMC Bioinform. 2010, 11, S41. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Lal, S.; Parker, S.J.; Nevler, A.; Vaziri-Gohar, A.; Dukleska, K.; Mambelli-Lisboa, N.C.; Moffat, C.; Blanco, F.F.; Chand, S.N.; et al. Posttranscriptional upregulation of IDH1 by HuR establishes a powerful survival phenotype in pancreatic cancer cells. Cancer Res. 2017, 77, 4460–4471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birsoy, K.; Possemato, R.; Lorbeer, F.K.; Bayraktar, E.C.; Thiru, P.; Yucel, B.; Wang, T.; Chen, W.W.; Clish, C.B.; Sabatini, D.M. Metabolic determinants of cancer cell sensitivity to glucose limitation and biguanides. Nature 2014, 508, 108–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeown, S.R. Defining normoxia, physoxia and hypoxia in tumours-implications for treatment response. Br. J. Radiol. 2014, 87, 20130676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumsey, W.L.; Schlosser, C.; Nuutinen, E.M.; Robiolio, M.; Wilson, D.F. Cellular energetics and the oxygen dependence of respiration in cardiac myocytes isolated from adult rat. J. Biol. Chem. 1990, 265, 15392–15402. [Google Scholar]
- Weinberg, S.E.; Chandel, N.S. Targeting mitochondria metabolism for cancer therapy. Nat. Chem. Biol. 2015, 11, 9–15. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takino, J.-i.; Sato, T.; Hiraishi, I.; Nagamine, K.; Hori, T. Alterations in Glucose Metabolism Due to Decreased Expression of Heterogeneous Nuclear Ribonucleoprotein M in Pancreatic Ductal Adenocarcinoma. Biology 2021, 10, 57. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010057
Takino J-i, Sato T, Hiraishi I, Nagamine K, Hori T. Alterations in Glucose Metabolism Due to Decreased Expression of Heterogeneous Nuclear Ribonucleoprotein M in Pancreatic Ductal Adenocarcinoma. Biology. 2021; 10(1):57. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010057
Chicago/Turabian StyleTakino, Jun-ichi, Takuma Sato, Isamu Hiraishi, Kentaro Nagamine, and Takamitsu Hori. 2021. "Alterations in Glucose Metabolism Due to Decreased Expression of Heterogeneous Nuclear Ribonucleoprotein M in Pancreatic Ductal Adenocarcinoma" Biology 10, no. 1: 57. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10010057