
Comparative Phenotypic and Agronomic Assessment of Transgenic Potato with 3R-Gene Stack with Complete Resistance to Late Blight Disease

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
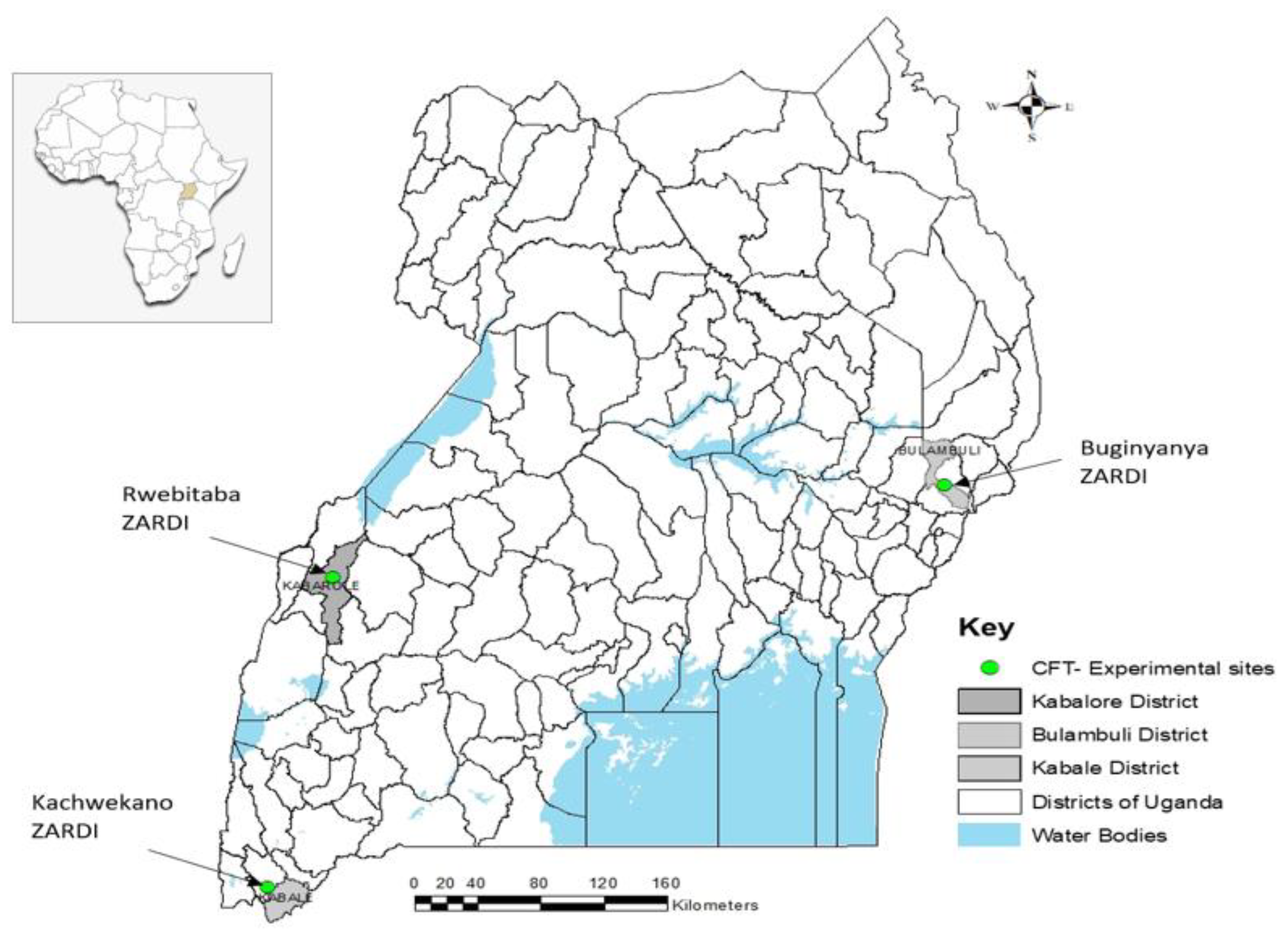
2.2. Experimental Sites
2.3. Experimental Design
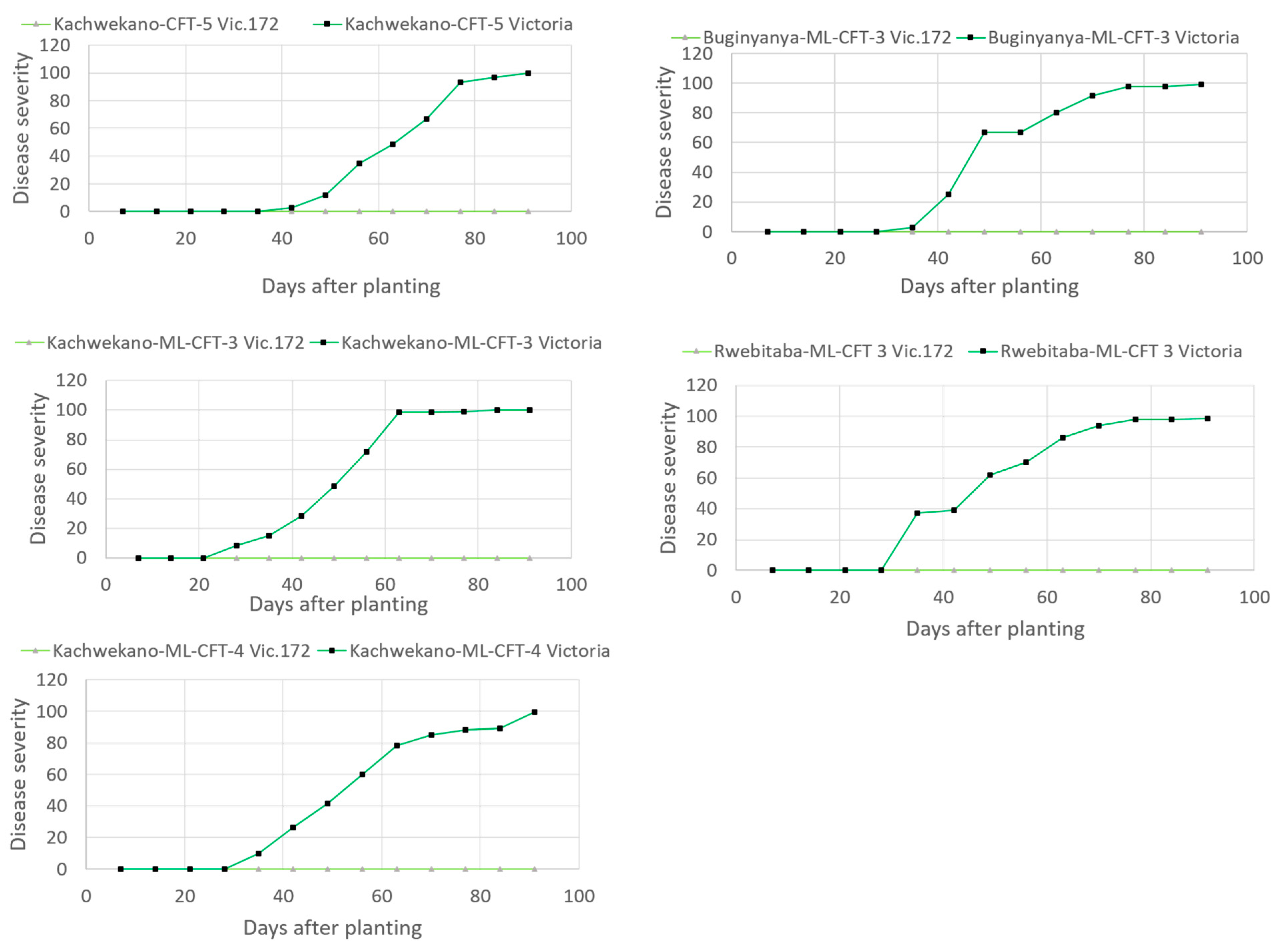
2.4. Late Blight Disease Scoring
2.5. Phenotypic Characterization
2.6. Agronomic Performance Assessment
2.7. Data Analysis
3. Results
3.1. Trait Efficacy in the Field
3.2. Phenotypic Characterization
3.2.1. Stems
3.2.2. Leaves
3.2.3. Flowers
3.2.4. Harvested Tubers
3.3. Agronomic Performances
3.3.1. Sprouting and Emergence
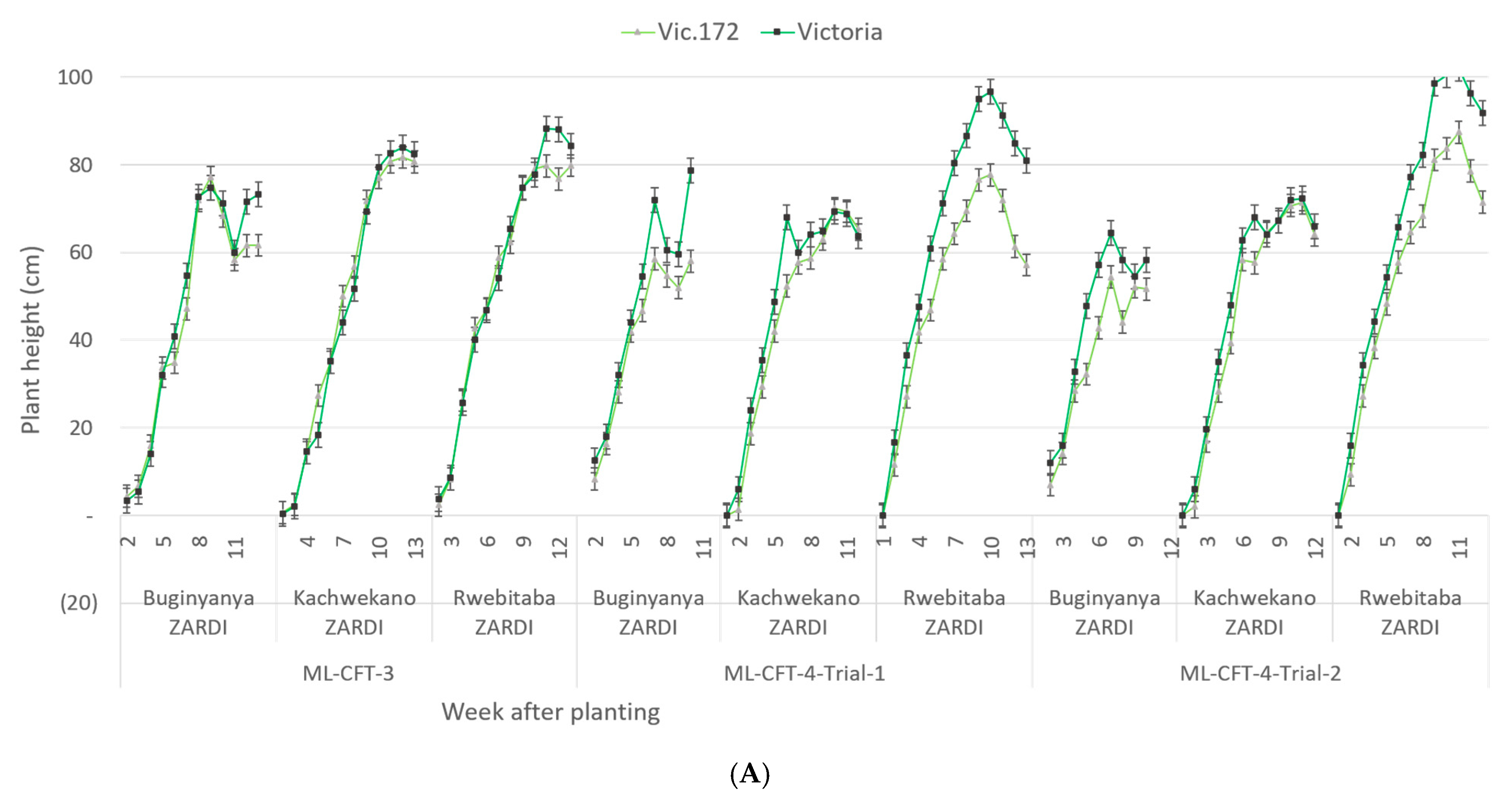
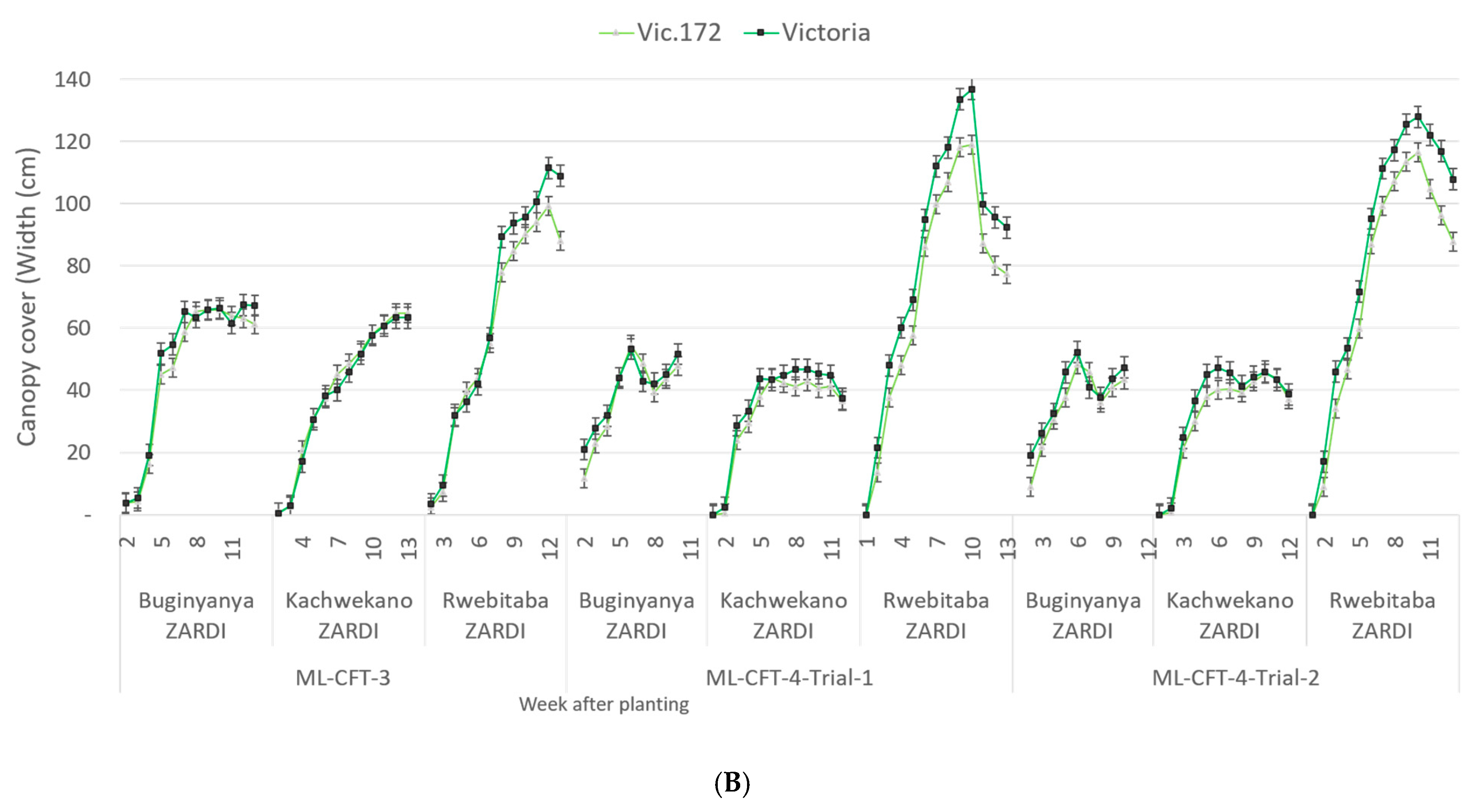
3.3.2. Plant and Foliage Growth Characterization
3.3.3. Plant Vigor
3.3.4. Yield Estimation
Marketable Yield of Both Genotypes across Locations
Number of Marketable Tubers of Each Genotype across Locations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global food security, contributions from sustainable potato agri-food systems. In The Potato Crop; Springer: Cham, Switzerland, 2020; pp. 3–35. [Google Scholar]
- UBOS; MAAIF. Uganda Census of Agriculture 2008/2009 Volume IV: Crop Area and Production Report December 2010; Uganda Bureau of Statistics (UBOS) and Ministry of Agriculture, Animal Industry and Fisheries (MAAIF): Kampala, Uganda, 2010.
- Hareau, G.; Kleinwechter, U.; Pradel, W.; Suarez, V.; Okello, J.; Vikraman, S. Strategic Assessment of Research Priorities for Potato. Lima (Peru) CGIAR Research Program on Roots, Tubers and Bananas (RTB) 2014, Working Paper 2014-8. Available online: www.rtb.cgiar.org (accessed on 8 August 2021).
- UBOS. Uganda Bureau of Statistics 2021. Available online: https://www.ubos.org/explore-statistics/2/ (accessed on 10 June 2021).
- UBOS. Uganda Census of Agriculture Crop Area and Production Report 2010; UBOS: Kampala, Uganda, 2010; Volume 4, p. 178. [Google Scholar]
- FAO. FAOSTAT Crop Data 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 June 2021).
- Namugga, P.; Sibiya, J.; Melis, R.; Barekye, A. Yield Response of Potato (Solanum tuberosum L.) Genotypes to late blight caused by Phytophthora infestans in Uganda. Am. J. Potato Res. 2018, 95, 423–434. [Google Scholar] [CrossRef]
- Kakuhenzire, R. Development of Decision Support Tools for Integrated Management of Potato Late Blight in the Highlands of Uganda. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2009. [Google Scholar]
- Namugga, P. Development of High Yielding and Early Maturing Potato (Solanum tuberosum L.) Genotypes with Resistance to Phytophthora infestans in Uganda. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2017. [Google Scholar]
- Malcolmson, J.F.; Black, W. New R genes in Solanum demissum Lindl. and their complementary races of Phytophthora infestans (Mont.) de Bary. Euphytica 1966, 15, 199–203. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Review and analysis of limitations in ways to improve conventional potato breeding. Potato Res. 2017, 60, 171–193. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Struik, P.C.; Visser, R.G.F.; Jacobsen, E. Applied biotechnology to combat late blight in potato caused by Phytophthora infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Kuhl, J.C.; Zarka, K.; Coombs, J.; Kirk, W.W.; Douches, D.S. Late blight resistance of RB transgenic potato lines. J. Am. Soc. Hortic. Sci. 2007, 132, 783–789. [Google Scholar] [CrossRef] [Green Version]
- Halterman, D.A.; Kramer, L.C.; Wielgus, S.; Jiang, J. Performance of transgenic potato containing the late blight resistance gene RB. Plant Dis. 2008, 92, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Witek, K.; Verweij, W.; Jupe, F.; Cooke, D.; Dorling, S.; Tomlinson, L.; Smoker, M.; Perkins, S.; Foster, S. Elevating crop disease resistance with cloned genes. Phil. Trans. R. Soc. B 2014, 369, 20130087. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Duangpan, S.; Bhaskar, P.B.; Wielgus, S.M.; Jiang, J. Constitutively expressed RB gene confers a high level but unregulated resistance to potato late blight. Am. J. Potato Res. 2018, 95, 575–583. [Google Scholar] [CrossRef]
- Richael, C.M. Development of the Genetically Modified Innate® Potato. Plant Breed. Rev. 2021, 44, 57–78. [Google Scholar]
- Birch, P.R.; Boevink, P.C.; Gilroy, E.M.; Hein, I.; Pritchard, L.; Whisson, S.C. Oomycete RXLR effectors: Delivery, functional redundancy and durable disease resistance. Curr. Opin. Plant Biol. 2008, 11, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Vleeshouwers, V.G.; Raffaele, S.; Vossen, J.H.; Champouret, N.; Oliva, R.; Segretin, M.E.; Rietman, H.; Cano, L.M.; Lokossou, A.; Kessel, G.; et al. Understanding and exploiting late blight resistance in the age of effectors. Annu. Rev. Phytopathol. 2011, 49, 507–531. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, V.G.; Oliver, R.P. Effectors as tools in disease resistance breeding against biotrophic, hemibiotrophic, and necrotrophic plant pathogens. Mol. Plant Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Li, Y.; Vossen, J.H.; Visser, R.G.; Jacobsen, E. Functional stacking of three resistance genes against Phytophthora infestans in potato. Transgenic Res. 2012, 21, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Haesaert, G.; Vossen, J.H.; Custers, R.; De Loose, M.; Haverkort, A.; Heremans, B.; Hutten, R.; Kessel, G.; Landschoot, S.; Van Droogenbroeck, B.; et al. Transformation of the potato variety Desiree with single or multiple resistance genes increases resistance to late blight under field conditions. Crop Prot. 2015, 77, 163–175. [Google Scholar] [CrossRef]
- Webi, E.N.; Kariuki, D.; Kinyua, J.; Njoroge, A.; Ghislain, M.; Magembe, E. Extreme resistance to late blight disease by transferring 3 R genes from wild relatives into African farmer-preferred potato varieties. Afr. J. Biotechnol. 2019, 18, 845–856. [Google Scholar]
- Ghislain, M.; Byarugaba, A.A.; Magembe, E.; Njoroge, A.; Rivera, C.; Román, M.L.; Tovar, J.C.; Gamboa, S.; Forbes, G.A.; Kreuze, J.F.; et al. Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnol. J. 2019, 17, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA GMO Panel (EFSA Panel on Genetically Modified Organisms). Guidance on the agronomic and phenotypic characterization of genetically modified plants. EFSA J. 2015, 13, 4128. [Google Scholar] [CrossRef] [Green Version]
- Forbes, G.A.; Charkowski, A.; Andrade-Piedra, J.; Parker, M.L.; Schulte-Geldermann, E. Potato seed systems. In The Potato Crop; Springer: Cham, Switzerland, 2020; pp. 431–447. [Google Scholar]
- Conner, A.J.; Williams, M.K.; Abernethy, D.J.; Fletcher, P.J.; Genet, R.A. Field performance of transgenic potatoes. N. Z. J. Crop Hortic. Sci. 1994, 22, 361–371. [Google Scholar] [CrossRef]
- Barrell, P.J.; Conner, A.J. Facilitating the recovery of phenotypically normal transgenic lines in clonal crops: A new strategy illustrated in potato. Theor. Appl. Genet. 2011, 122, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Larkin, P.J.; Scowcroft, W.R. Somaclonal variation—A novel source of variability from cell cultures for plant improvement. Theor. Appl. Genet. 1981, 60, 197–214. [Google Scholar] [CrossRef]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassar, A.M.; Abdulnour, J.; Leclerc, Y.; Li, X.Q.; Donnelly, D.J. Intraclonal selection for improved processing of NB ‘Russet Burbank’potato. Am. J. Potato Res. 2011, 88, 387–397. [Google Scholar] [CrossRef]
- Thiele, G.; Hareau, G.; Suarez, V.; Chujoy, E.; Bonierbale, M.; Maldonado, L. Varietal change in potatoes in developing countries and the contribution of the International Potato Center: 1972–2007. In Social Sciences Working Paper; International Potato Center (CIP): Lima, Peru, 2008; Volume 6, p. 46. [Google Scholar]
- Forbes, G.; Perez, W.; Andrade-Piedra, J. Field Assessment of Resistance In Potato to Phytophthora infestans: International Cooperators Guide; International Potato Center: Lima, Peru, 2014. [Google Scholar]
- Millett, B.P.; Gao, L.; Iorizzo, M.; Carputo, D.; Bradeen, J.M. Potato tuber blight resistance phenotypes correlate with RB transgene transcript levels in an age-dependent manner. Phytopathology 2015, 105, 1131–1136. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.W. GenStat. Wiley Interdiscip. Rev. Comput. Stat. 2009, 1, 255–258. [Google Scholar] [CrossRef]
- Williams, L.J.; Abdi, H. Fisher’s least significant difference (LSD) test. Encycl. Res. Des. 2010, 218, 840–853. [Google Scholar]
- Fuglie, K.O. Priorities for potato research in developing countries: Results of a survey. Am. J. Potato Res. 2007, 84, 353–365. [Google Scholar] [CrossRef]
- Njoroge, A.W.; Andersson, B.; Lees, A.K.; Mutai, C.; Forbes, G.A.; Yuen, J.E.; Pelle, R. Genotyping of Phytophthora infestans in Eastern Africa reveals a dominating invasive European lineage. Phytopathology 2019, 109, 670–680. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f. 1 | s.s. 2 | m.s. 3 | v.r. 4 | F pr. 5 |
|---|---|---|---|---|---|
| Genotype | 1 | 129.98 | 129.98 | 2.7 | 0.126 |
| Location | 2 | 1409.71 | 704.86 | 14.65 | <0.001 |
| Genotype X Location | 2 | 76.04 | 38.02 | 0.79 | 0.476 |
| Residual | 12 | 577.4 | 48.12 | ||
| Total | 17 | 2193.13 |
| Source of Variation | d.f. 1 | s.s. 2 | m.s. 3 | v.r. 4 | F pr.5 |
|---|---|---|---|---|---|
| Genotype | 1 | 5.675 | 5.675 | 1.26 | 0.284 |
| Location | 2 | 18.88 | 9.44 | 2.09 | 0.166 |
| Genotype X Location | 2 | 0.105 | 0.053 | 0.01 | 0.988 |
| Residual | 12 | 54.177 | 4.515 | ||
| Total | 17 | 78.837 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byarugaba, A.A.; Baguma, G.; Jjemba, D.M.; Faith, A.K.; Wasukira, A.; Magembe, E.; Njoroge, A.; Barekye, A.; Ghislain, M. Comparative Phenotypic and Agronomic Assessment of Transgenic Potato with 3R-Gene Stack with Complete Resistance to Late Blight Disease. Biology 2021, 10, 952. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100952
Byarugaba AA, Baguma G, Jjemba DM, Faith AK, Wasukira A, Magembe E, Njoroge A, Barekye A, Ghislain M. Comparative Phenotypic and Agronomic Assessment of Transgenic Potato with 3R-Gene Stack with Complete Resistance to Late Blight Disease. Biology. 2021; 10(10):952. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100952
Chicago/Turabian StyleByarugaba, Arinaitwe Abel, Gerald Baguma, Douglas Mutebi Jjemba, Aharinta Kenneth Faith, Arthur Wasukira, Eric Magembe, Anne Njoroge, Alex Barekye, and Marc Ghislain. 2021. "Comparative Phenotypic and Agronomic Assessment of Transgenic Potato with 3R-Gene Stack with Complete Resistance to Late Blight Disease" Biology 10, no. 10: 952. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100952