
Transcriptome and Metabolome Analysis Revealed the Freezing Resistance Mechanism in 60-Year-Old Overwintering Camellia sinensis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. Transcriptome Sequencing, Identification, and Analysis of Differentially Expressed Genes (DEGs)
2.3. Untargeted Metabolomics Analysis
2.4. Metabolome Data Processing and Analysis
3. Results
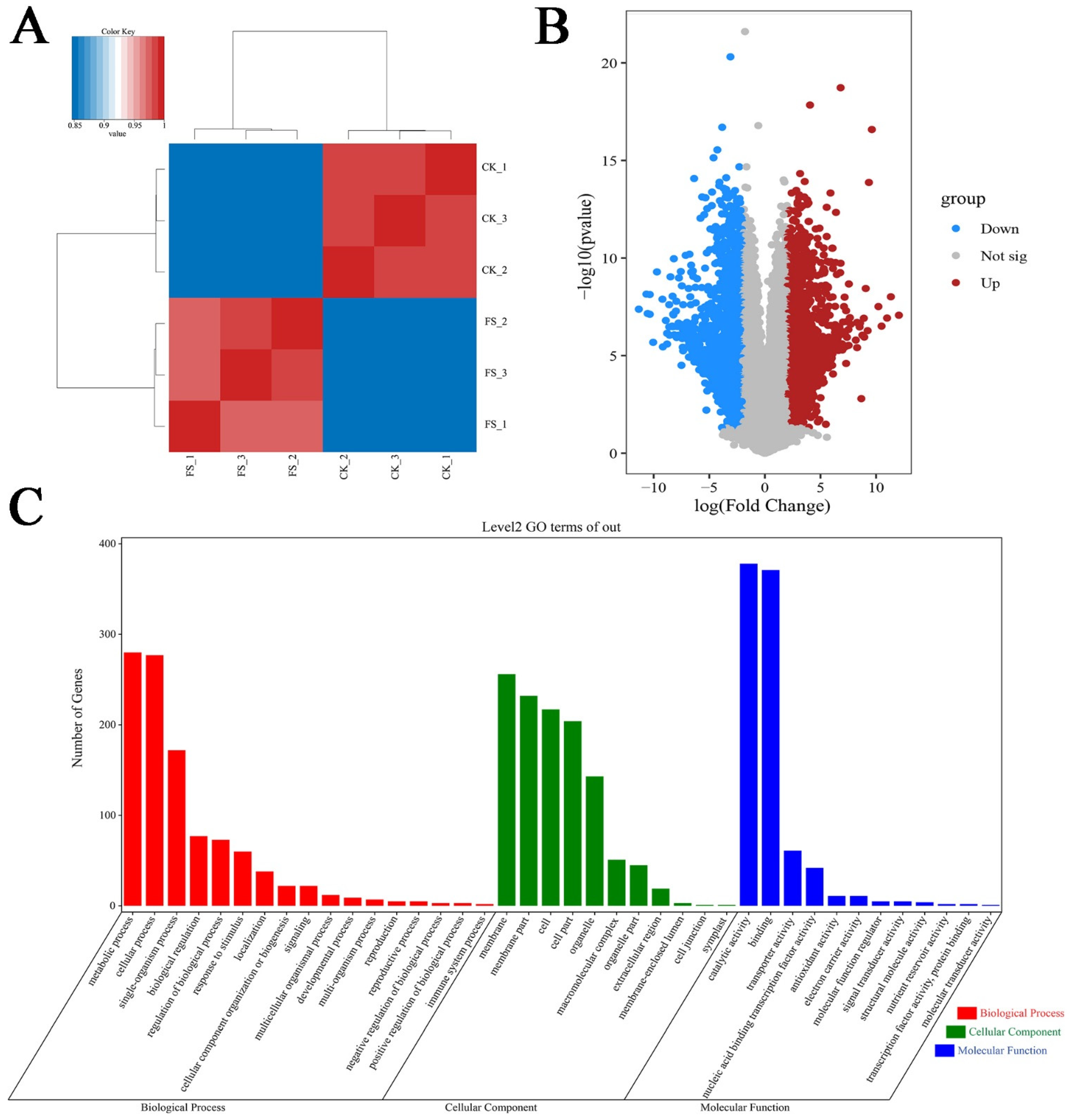
3.1. C. sinensis Transcriptome Sequencing Results and Data Analyses
3.2. Transcriptional Characteristics of C. sinensis Response to Freezing Stress
3.3. Pathway Analysis in Response to Freezing Stress
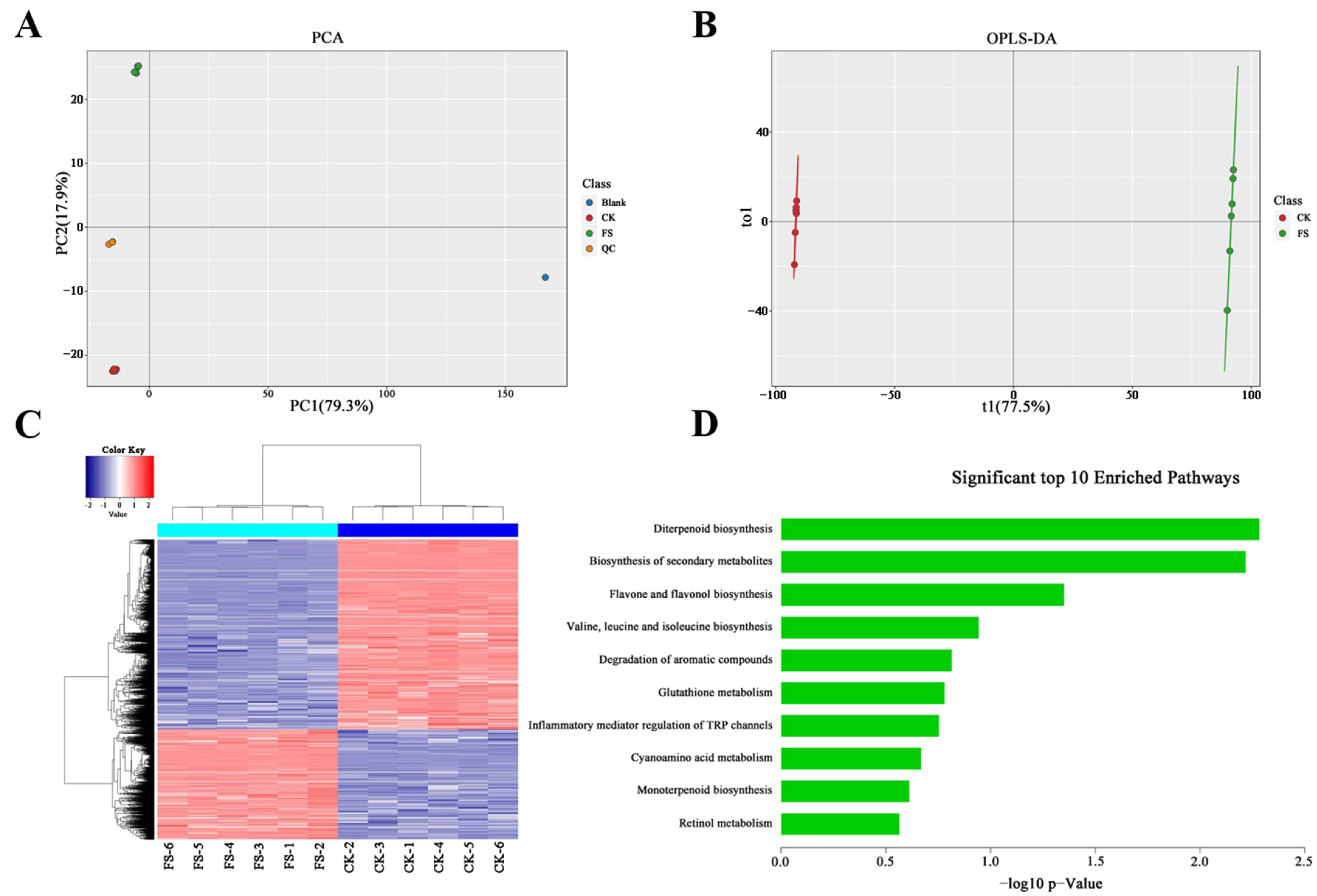
3.4. Metabolic Characteristics of C. sinensis Response to Freezing Stress
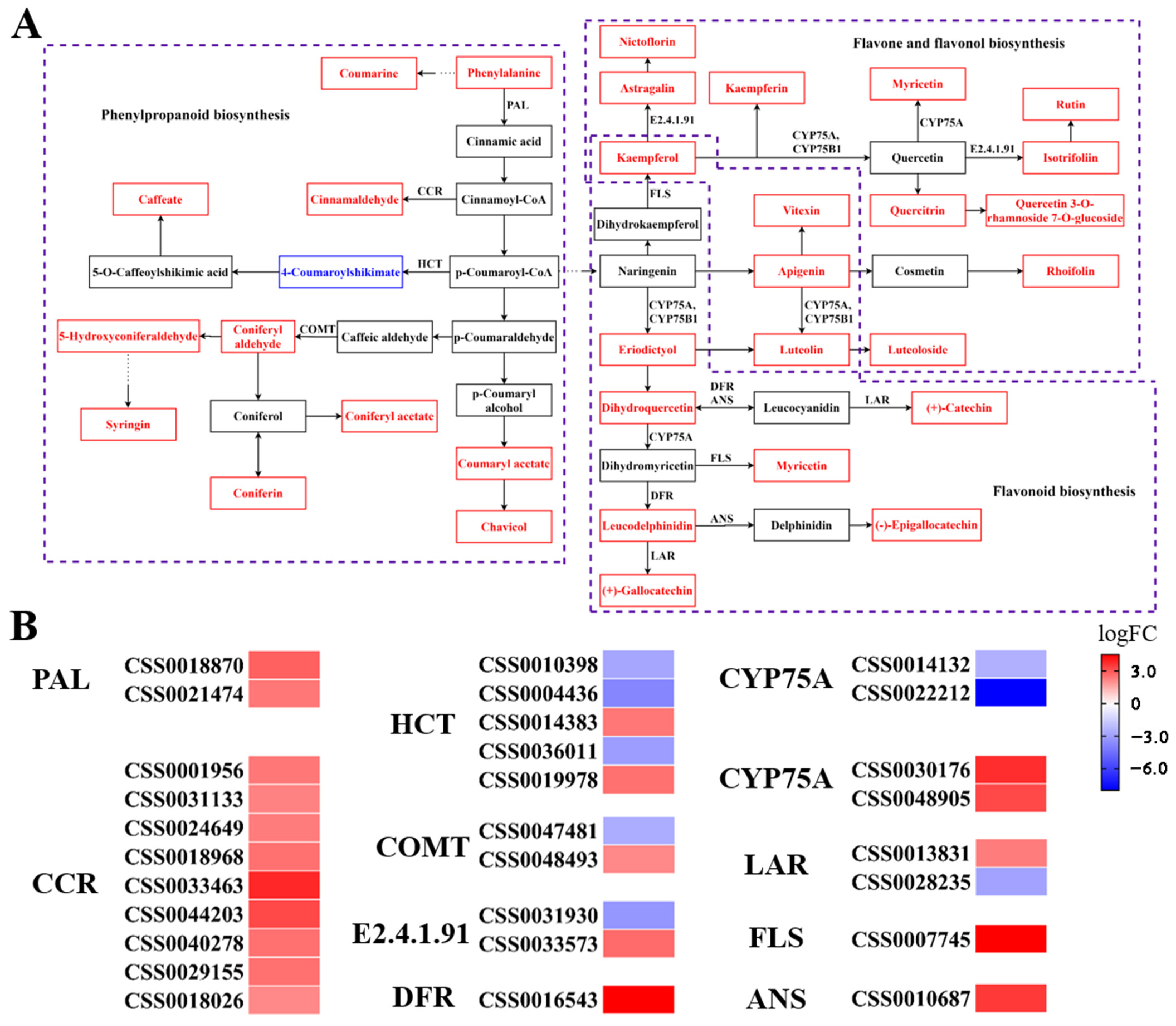
3.5. Correlation Analysis of Gene Expression and Metabolite Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The reference genome of tea plant and resequencing of 81 diverse accessions provide insights into genome evolution and adaptation of tea plants. Mol. Plant. 2020, 13, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Li, X.; Liu, A.; Chen, S. Physiological and defense responses of tea plants to elevated CO2: A review. Front. Plant. Sci. 2020, 11, 305. [Google Scholar] [CrossRef]
- Karak, T.; Kutu, F.R.; Nath, J.R.; Sonar, I.; Paul, R.K.; Boruah, R.K.; Sanyal, S.; Sabhapondit, S.; Dutta, A.K. Micronutrients (B, Co, Cu, Fe, Mn, Mo, and Zn) content in made tea (Camellia sinensis L.) and tea infusion with health prospect: A critical review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2996–3034. [Google Scholar] [CrossRef]
- Wang, X.; Feng, H.; Chang, Y.; Ma, C.; Wang, L.; Hao, X.; Li, A.; Cheng, H.; Wang, L.; Cui, P.; et al. Population sequencing enhances understanding of tea plant evolution. Nat. Commun. 2020, 11, 4447. [Google Scholar] [CrossRef]
- Bredow, M.; Walker, V.K. Ice-binding proteins in plants. Front. Plant. Sci. 2017, 8, 2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Cai, Z.; Li, D.; Lu, J.; Ye, J.; Liang, Y.; Zheng, X. Effect of freezing on photosystem II and assessment of freezing tolerance of tea cultivar. Plants 2019, 8, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Zhao, L.; Wang, Y.; Shen, J.; Zhang, Y.; Jia, S.; Li, Y.; Ding, Z. Integrated RNA-Seq and sRNA-Seq analysis identifies chilling and freezing responsive key molecular players and pathways in tea plant (Camellia sinensis). PLoS ONE 2015, 10, e0125031. [Google Scholar] [CrossRef] [PubMed]
- Charrier, G.; Ngao, J.; Saudreau, M.; Améglio, T. Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees. Front. Plant. Sci. 2015, 6, 259. [Google Scholar] [CrossRef] [Green Version]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Jaikumar, N.S.; Dorn, K.M.; Baas, D.; Wilke, B.; Kapp, C.; Snapp, S.S. Nucleic acid damage and DNA repair are affected by freezing stress in annual wheat (Triticum aestivum) and by plant age and freezing in its perennial relative (Thinopyrum intermedium). Am. J. Bot. 2020, 107, 1693–1709. [Google Scholar] [CrossRef]
- Lim, C.J.; Park, J.; Shen, M.; Park, H.J.; Cheong, M.S.; Park, K.S.; Baek, D.; Bae, M.J.; Ali, A.; Jan, M.; et al. The histone-modifying complex PWR/HOS15/HD2C epigenetically regulates cold tolerance. Plant. Physiol. 2020, 184, 1097–1111. [Google Scholar] [CrossRef]
- Sun, S.; Fang, J.; Lin, M.; Hu, C.; Qi, X.; Chen, J.; Zhong, Y.; Muhammad, A.; Li, Z.; Li, Y. Comparative metabolomic and transcriptomic studies reveal key metabolism pathways contributing to freezing tolerance under cold stress in kiwifruit. Front. Plant Sci. 2021, 12, 628969. [Google Scholar] [CrossRef]
- Lu, H.; Xue, L.; Cheng, J.; Yang, X.; Xie, H.; Song, X.; Qiang, S. Polyploidization-driven differentiation of freezing tolerance in Solidago Canadensis. Plant Cell Environ. 2020, 43, 1394–1403. [Google Scholar] [CrossRef]
- do Amaral, M.C.F.; Frisbie, J.; Crum, R.J.; Goldstein, D.L.; Krane, C.M. Hepatic transcriptome of the freeze-tolerant Cope’s gray treefrog, Dryophytes chrysoscelis: Responses to cold acclimation and freezing. BMC Genom. 2020, 21, 226. [Google Scholar] [CrossRef] [PubMed]
- Hajihashemi, S.; Brestic, M.; Landi, M.; Skalicky, M. Resistance of Fritillaria imperialis to freezing stress through gene expression, osmotic adjustment and antioxidants. Sci Rep. 2020, 10, 10427. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Lv, Z.; Khanzada, A.; Huang, M.; Cai, J.; Zhou, Q.; Huo, Z.; Jiang, D. Effects of cold and salicylic acid priming on free proline and sucrose accumulation in winter wheat underfreezing stress. J. Plant Growth Regul. 2021, 204, 1–14. [Google Scholar]
- Pu, Y.; Liu, L.; Wu, J.; Zhao, Y.; Bai, J.; Ma, L.; Yue, J.; Jin, J.; Niu, Z.; Fang, Y.; et al. Transcriptome profile analysis of winter rapeseed (Brassica napus L.) in response to freezing stress, reveal potentially connected events to freezing stress. Int. J. Mol. Sci. 2019, 20, 2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Wang, Y.H.; Liu, J.X.; Feng, K.; Xu, Z.S.; Xiong, A.S. Advances in genomic, transcriptomic, proteomic, and metabolomic approaches to study biotic stress in fruit crops. Crit. Rev. Biotechnol. 2019, 39, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Xiaodong, M.; Yunhe, C.; Junxiang, L.; Junzhu, Z.; Feifei, Z.; Zhenyuan, S.; Lei, H. Transcriptomic and metabolomic insights into the adaptive response of Salix viminalis to phenanthrene. Chemosphere 2021, 262, 127573. [Google Scholar] [CrossRef]
- Lin, S.; Zeng, S.; Yang, X.; Yang, T.; Zheng, G.; Mao, G.; Wang, Y. Integrative analysis of transcriptome and metabolome reveals salt stress orchestrating the accumulation of specialized metabolites in Lycium barbarum L. fruit. Int. J. Mol. Sci. 2021, 22, 4414. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, X.; Ban, Q.; Zhu, X.; Jiang, C.; Wei, C.; Bennetzen, J.L. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genom. 2019, 20, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarina, L.S.; Bobrovskikh, A.V.; Doroshkov, A.V.; Malyukova, L.S.; Matskiv, A.O.; Rakhmangulov, R.S.; Koninskaya, N.G.; Malyarovskaya, V.I.; Tong, W.; Xia, E.; et al. Comparative expression analysis of stress-inducible candidate genes in response to cold and drought in tea plant [Camellia sinensis (L.) Kuntze]. Front. Genet. 2020, 11, 611283. [Google Scholar] [CrossRef]
- Lin, S.R.; Yu, S.Y.; Chang, T.D.; Lin, Y.J.; Wen, C.J.; Lin, Y.H. First report of anthracnose caused by Colletotrichum fructicola on tea in Taiwan. Plant. Dis. 2020, 3, 105. [Google Scholar] [CrossRef]
- Yao, L.; Ding, C.; Hao, X.; Zeng, J.; Yang, Y.; Wang, X.; Wang, L. CsSWEET1a and CsSWEET17 mediate growth and freezing tolerance by promoting sugar transport across the plasma membrane. Plant. Cell Physiol. 2020, 61, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Passow, C.N.; Kono, T.J.Y.; Stahl, B.A.; Jaggard, J.B.; Keene, A.C.; McGaugh, S.E. Nonrandom RNAseq gene expression associated with RNAlater and flash freezing storage methods. Mol. Ecol. Resour. 2019, 19, 456–464. [Google Scholar] [CrossRef]
- Palomares, M.A.; Dalmasso, C.; Bonnet, E.; Derbois, C.; Brohard-Julien, S.; Ambroise, C.; Battail, C.; Deleuze, J.F.; Olaso, R. Systematic analysis of TruSeq, SMARTer and SMARTer Ultra-Low RNA-seq kits for standard, low and ultra-low quantity samples. Sci. Rep. 2019, 9, 7550. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Li, H.; Zhang, W.; Tang, H.; Yang, L. Transcriptional regulation and functional analysis of Nicotiana tabacum under salt and ABA stress. Biochem. Biophys. Res. Commun. 2021, 570, 110–116. [Google Scholar] [CrossRef]
- Soler-García, A.A.; De Jesús, A.J.; Taylor, K.; Brown, E.W. Differentiation of Salmonella strains from the SARA, SARB and SARC reference collections by using three genes PCR-RFLP and the 2100 Agilent Bioanalyzer. Front. Microbiol. 2014, 5, 417. [Google Scholar]
- Holm, J.B.; Humphrys, M.S.; Robinson, C.K.; Settles, M.L.; Ott, S.; Fu, L.; Yang, H.; Gajer, P.; He, X.; McComb, E.; et al. Ultrahigh-throughput multiplexing and sequencing of > 500-base-pair amplicon regions on the Illumina HiSeq 2500 platform. mSystems 2019, 4, e00029-19. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, J.A.; Al-Harbi, K.M.; Ramzan, K.; Albalawi, A.M.; Mehmood, A.; Samman, M.I.; Basit, S. A novel splice-site mutation in the ASPM gene underlies autosomal recessive primary microcephaly. Ann. Saudi Med. 2016, 36, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Ward, C.M.; To, T.H.; Pederson, S.M. ngsReports: A Bioconductor package for managing FastQC reports and other NGS related log files. Bioinformatics 2020, 36, 2587–2588. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, P.; Su, H.; Cai, L.; Zhao, L.; Zhou, H. Bioinformatics analysis of neuroblastoma miRNA based on GEO data. Pharmgenomics Pers. Med. 2021, 14, 849–858. [Google Scholar] [PubMed]
- Harris, M.A.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]
- Angeloni, S.; Navarini, L.; Khamitova, G.; Maggi, F.; Sagratini, G.; Vittori, S.; Caprioli, G. A new analytical method for the simultaneous quantification of isoflavones and lignans in 25 green coffee samples by HPLC-MS/MS. Food Chem. 2020, 325, 126924. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Sano, M.; Kuroda, H.; Ohara, H.; Ando, H.; Matsumoto, K. DISCOVER: A facile structure-based screening method for vinyl compound producing microbes. Sci. Rep. 2019, 9, 16007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.R.; Chen, R.J.; Zhao, F.; Zhang, H.H.; Bi, Q.Y.; Zhang, Y.N.; Zhang, Y.Q.; Wu, Z.C.; Ji, X.M. Effect of Wenxia Changfu formula combined with cisplatin reversing non-small cell lung cancer cell adhesion-mediated drug resistance. Front. Pharmacol. 2020, 11, 500137. [Google Scholar] [CrossRef]
- Ng, B.; Quinete, N.; Gardinali, P.R. Assessing accuracy, precision and selectivity using quality controls for non-targeted analysis. Sci. Total Environ. 2020, 713, 136568. [Google Scholar] [CrossRef]
- Skedgel, C.; Regier, D.A. Constant-sum paired comparisons for eliciting stated preferences: A tutorial. Patient 2015, 8, 155–163. [Google Scholar] [CrossRef]
- Webb-Robertson, B.J.; Lowry, D.F.; Jarman, K.H.; Harbo, S.J.; Meng, Q.R.; Fuciarelli, A.F.; Pounds, J.G.; Lee, K.M. A study of spectral integration and normalization in NMR-based metabonomic analyses. J. Pharm. Biomed. Anal. 2005, 39, 830–836. [Google Scholar] [CrossRef]
- Mi, J.X.; Zhang, Y.N.; Lai, Z.; Li, W.; Zhou, L.; Zhong, F. Principal component analysis based on nuclear norm minimization. Neural Netw. 2019, 118, 1–16. [Google Scholar] [CrossRef]
- Yang, Q.; Tian, G.L.; Qin, J.W.; Wu, B.Q.; Tan, L.; Xu, L.; Wu, S.Z.; Yang, J.T.; Jiang, J.H.; Yu, R.Q. Coupling bootstrap with synergy self-organizing map-based orthogonal partial least squares discriminant analysis: Stable metabolic biomarker selection for inherited metabolic diseases. Talanta 2020, 219, 121370. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Ambigaipalan, P.; Oh, W.Y.; Shahidi, F. Epigallocatechin (EGC) esters as potential sources of antioxidants. Food Chem. 2020, 309, 125609. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Alseekh, S.; Fernie, A.R. Conservation and diversification of flavonoid metabolism in the plant kingdom. Curr. Opin. Plant. Biol. 2020, 55, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Rubio, S.; Noriega, X.; Pérez, F.J. Abscisic acid (ABA) and low temperatures synergistically increase the expression of CBF/DREB1 transcription factors and cold-hardiness in grapevine dormant buds. Ann. Bot. 2019, 123, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, F.; Nong, S.; Liao, J.; Xing, A.; Shen, Q.; Ma, Y.; Fang, W.; Zhu, X. Effects of nitric oxide on the GABA, polyamines, and proline in tea (Camellia sinensis) roots under cold stress. Sci. Rep. 2020, 10, 12240. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, Y.; Dang, P.; Zhao, S.; Lai, D.; Zhou, L. Rice secondary metabolites: Structures, roles, biosynthesis, and metabolic regulation. Molecules. 2018, 23, 3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttner, L.; Grabe, V.; Gablenz, S.; Böhme, N.; Appenroth, K.J.; Gershenzon, J.; Huber, M. Differential localization of flavonoid glucosides in an aquatic plant implicates different functions under abiotic stress. Plant Cell Environ. 2021, 44, 900–914. [Google Scholar] [CrossRef]
- Knez Hrnčič, M.; Ivanovski, M.; Cör, D.; Knez, Ž. Chia Seeds (Salvia hispanica L.): An overview-phytochemical profile, isolation methods, and application. Molecules 2019, 25, 11. [Google Scholar] [CrossRef] [Green Version]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Flavonoids are determinants of freezing tolerance and cold acclimation in Arabidopsis thaliana. Sci. Rep. 2016, 6, 34027. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jin, J.; Gao, T.; Zhang, N.; Jing, T.; Wang, J.; Ban, Q.; Schwab, W.; Song, C. Glucosyltransferase CsUGT78A14 regulates flavonols accumulation and reactive oxygen species scavenging in response to cold stress in Camellia sinensis. Front. Plant. Sci. 2019, 10, 1675. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Wu, Z.; Wang, Y.; Ding, J.; Zheng, Y.; Tang, H.; Yang, L. Transcriptome and Metabolome Analysis Revealed the Freezing Resistance Mechanism in 60-Year-Old Overwintering Camellia sinensis. Biology 2021, 10, 996. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100996
Wu H, Wu Z, Wang Y, Ding J, Zheng Y, Tang H, Yang L. Transcriptome and Metabolome Analysis Revealed the Freezing Resistance Mechanism in 60-Year-Old Overwintering Camellia sinensis. Biology. 2021; 10(10):996. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100996
Chicago/Turabian StyleWu, Hui, Zixian Wu, Yuanheng Wang, Jie Ding, Yalin Zheng, Heng Tang, and Long Yang. 2021. "Transcriptome and Metabolome Analysis Revealed the Freezing Resistance Mechanism in 60-Year-Old Overwintering Camellia sinensis" Biology 10, no. 10: 996. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10100996