Modeling Dengue Immune Responses Mediated by Antibodies: A Qualitative Study

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Modeling Within-Host Dengue Infections
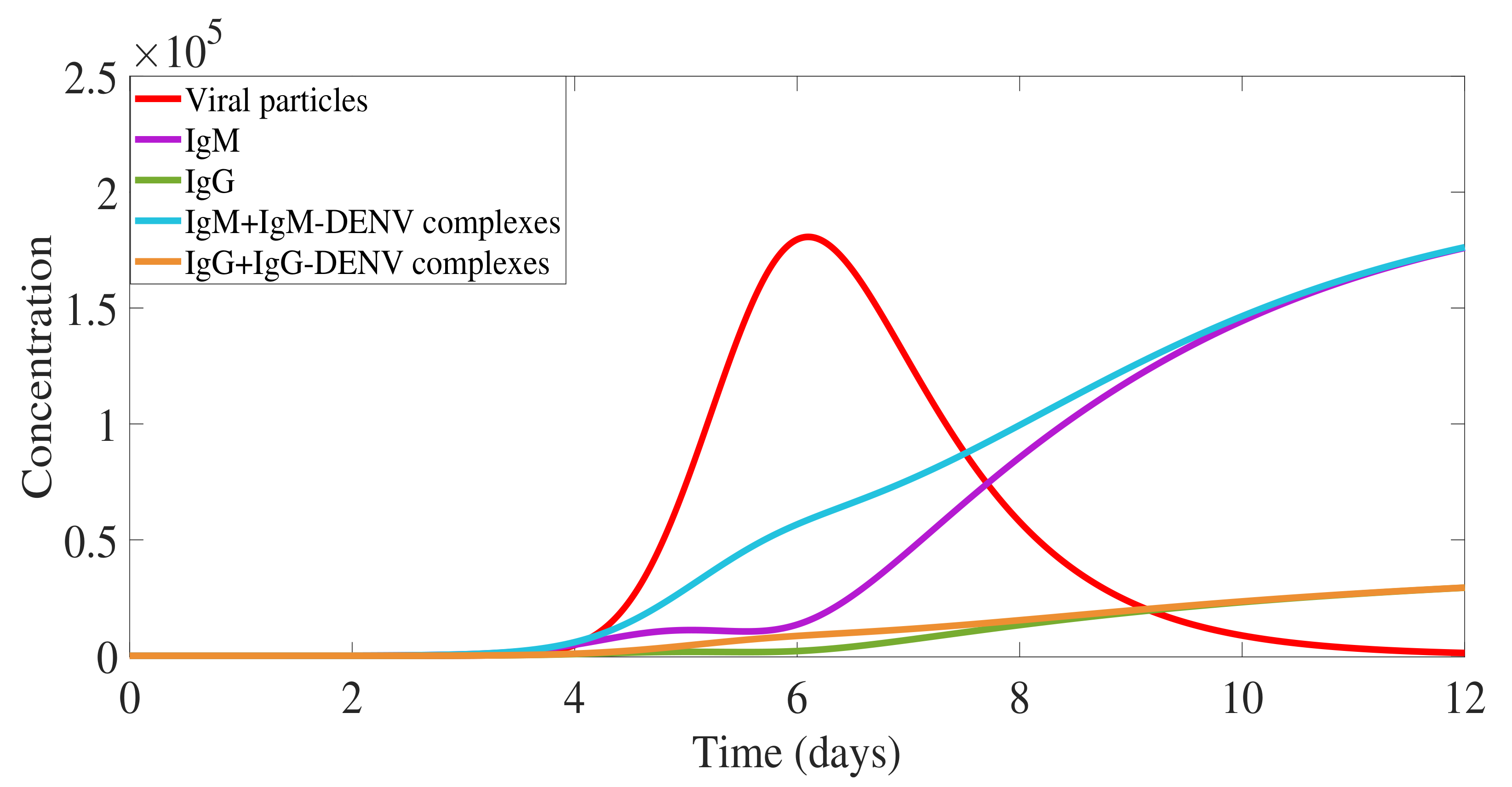
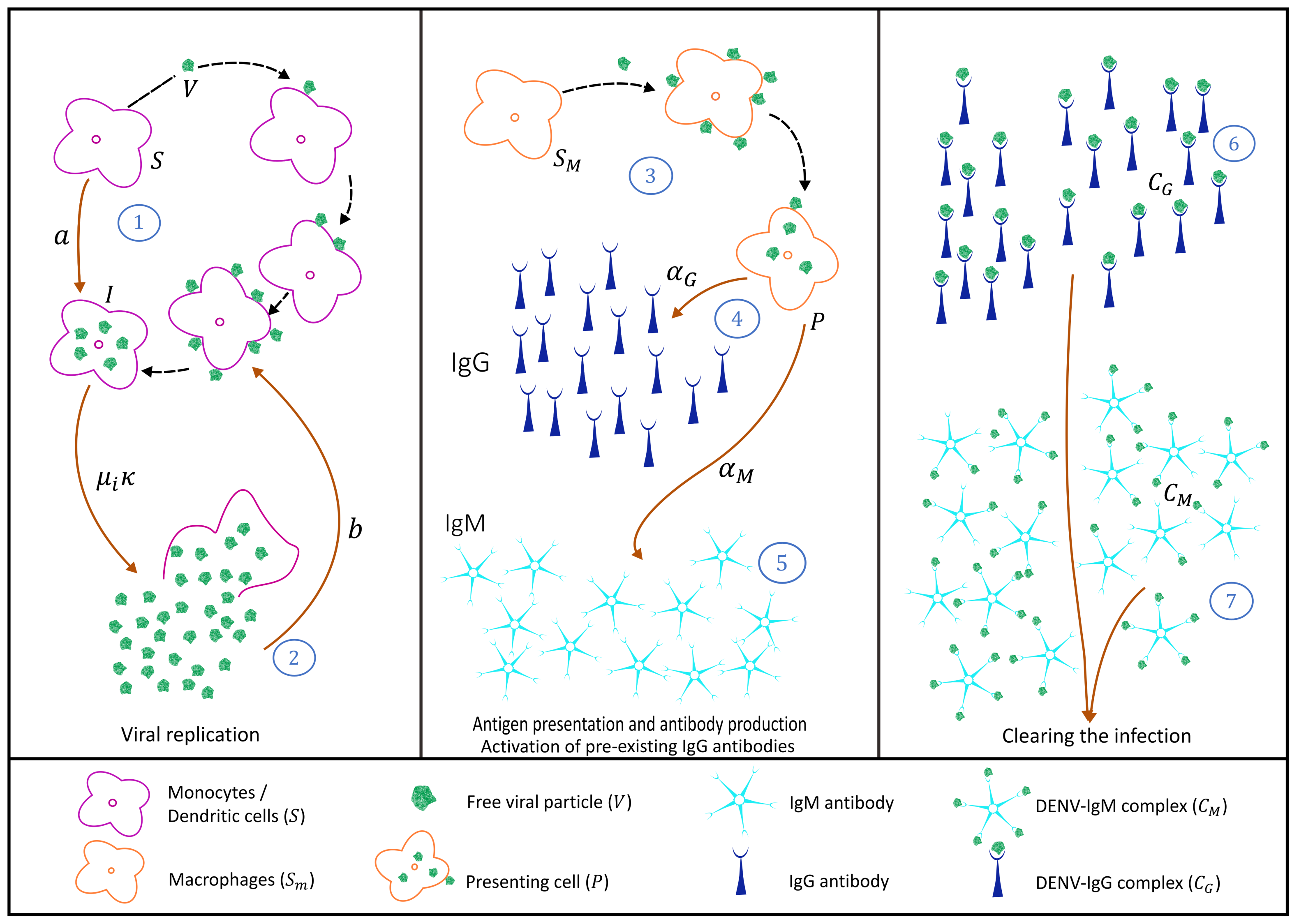
2.1. Primary Dengue Infection Model
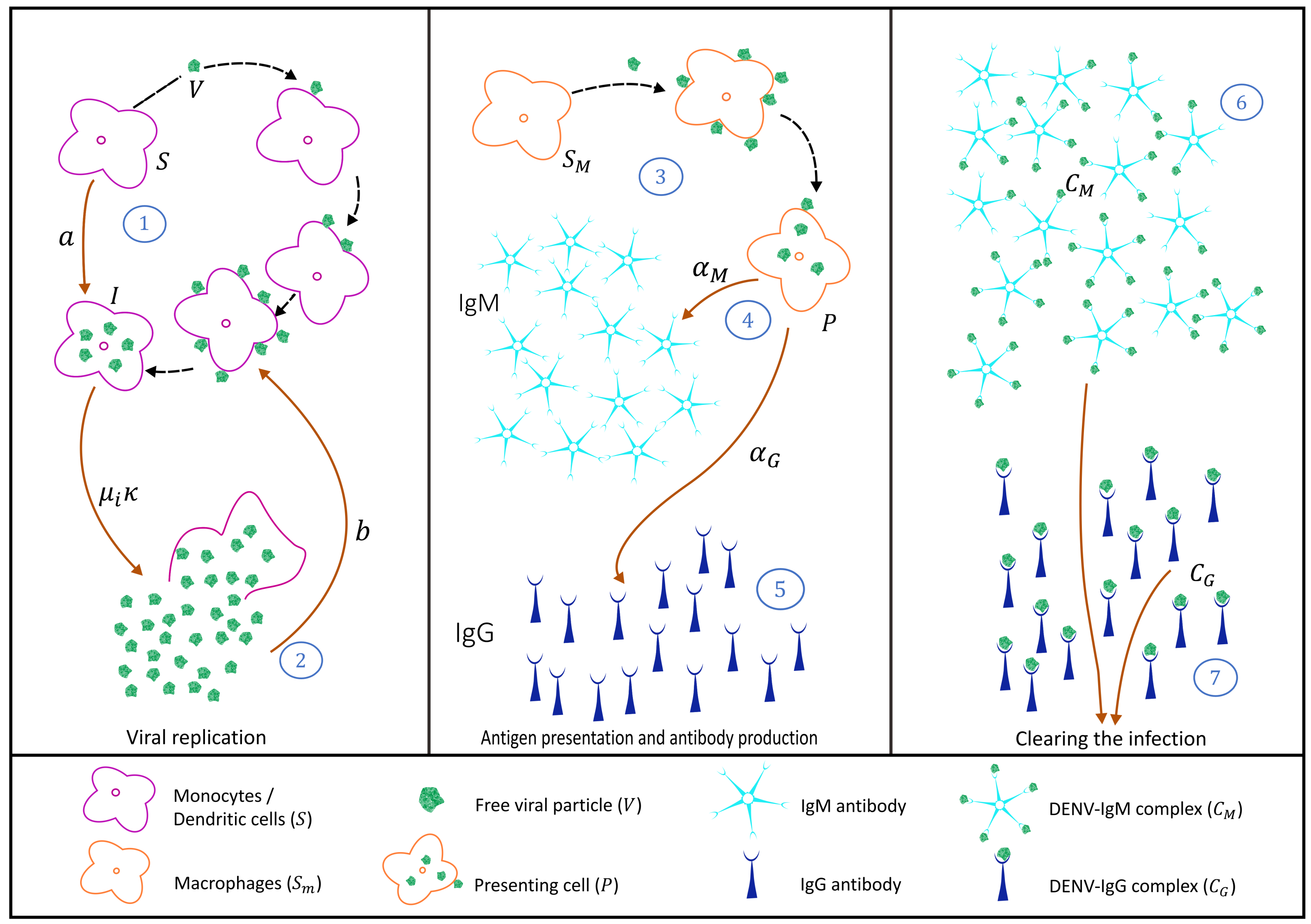
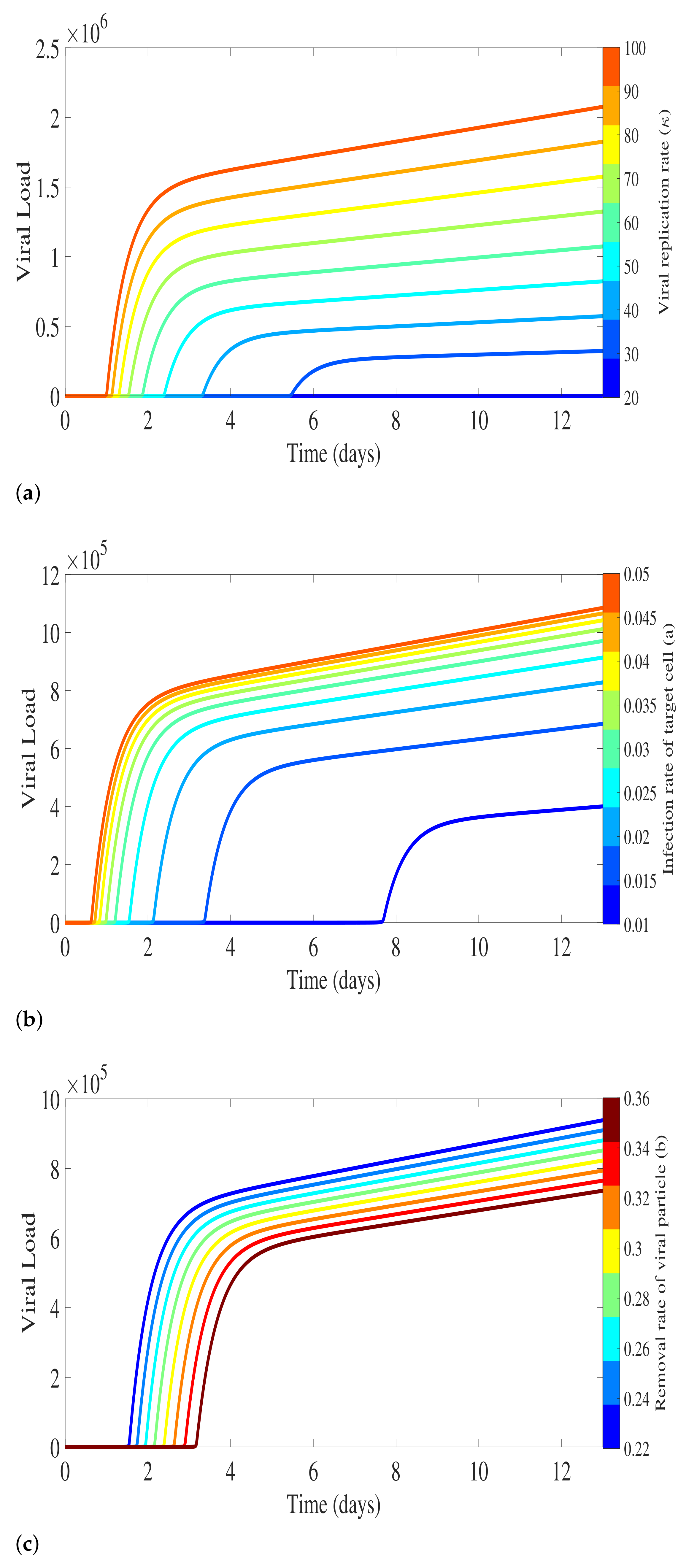
2.1.1. Virus Replication Dynamics
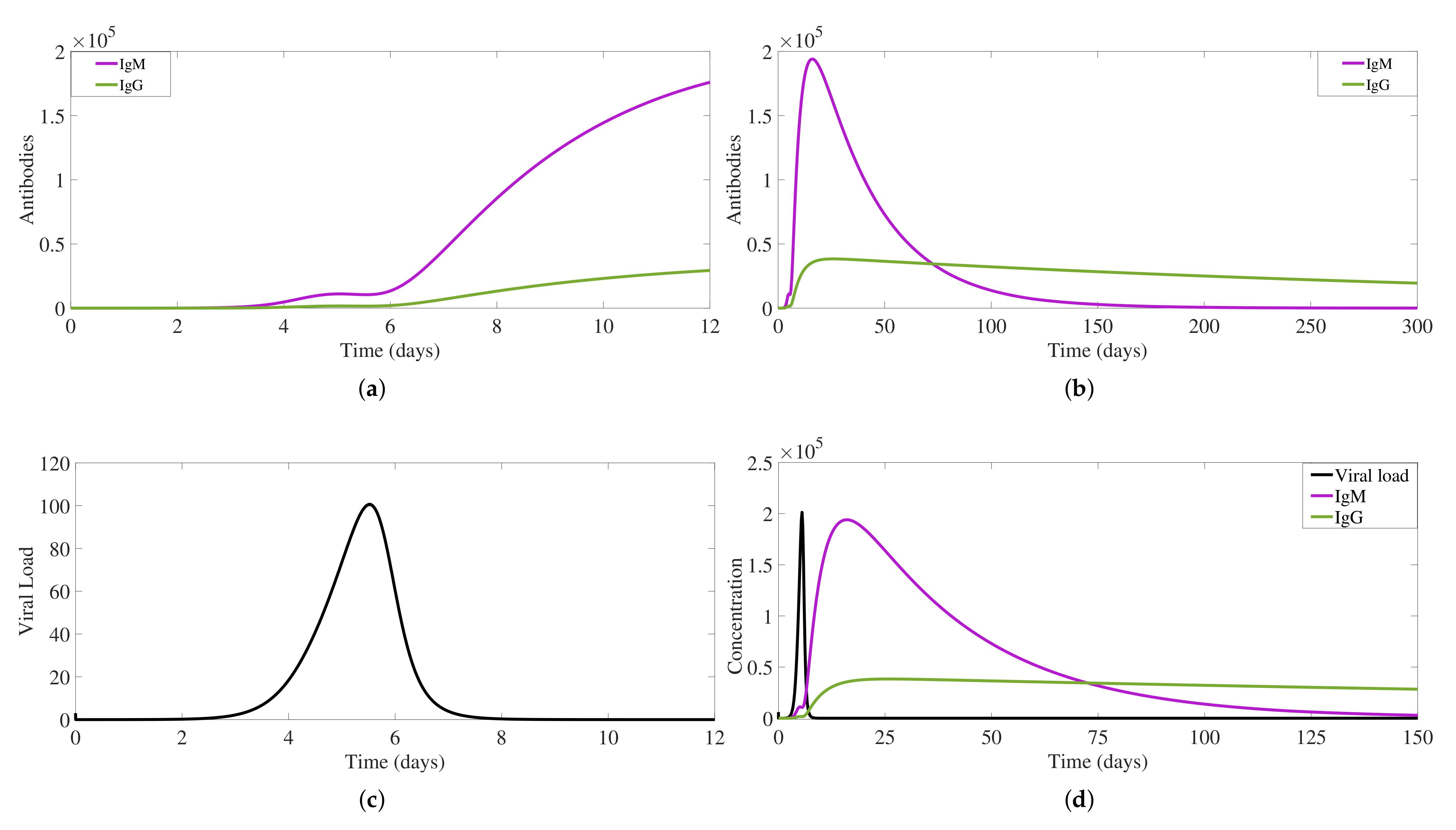
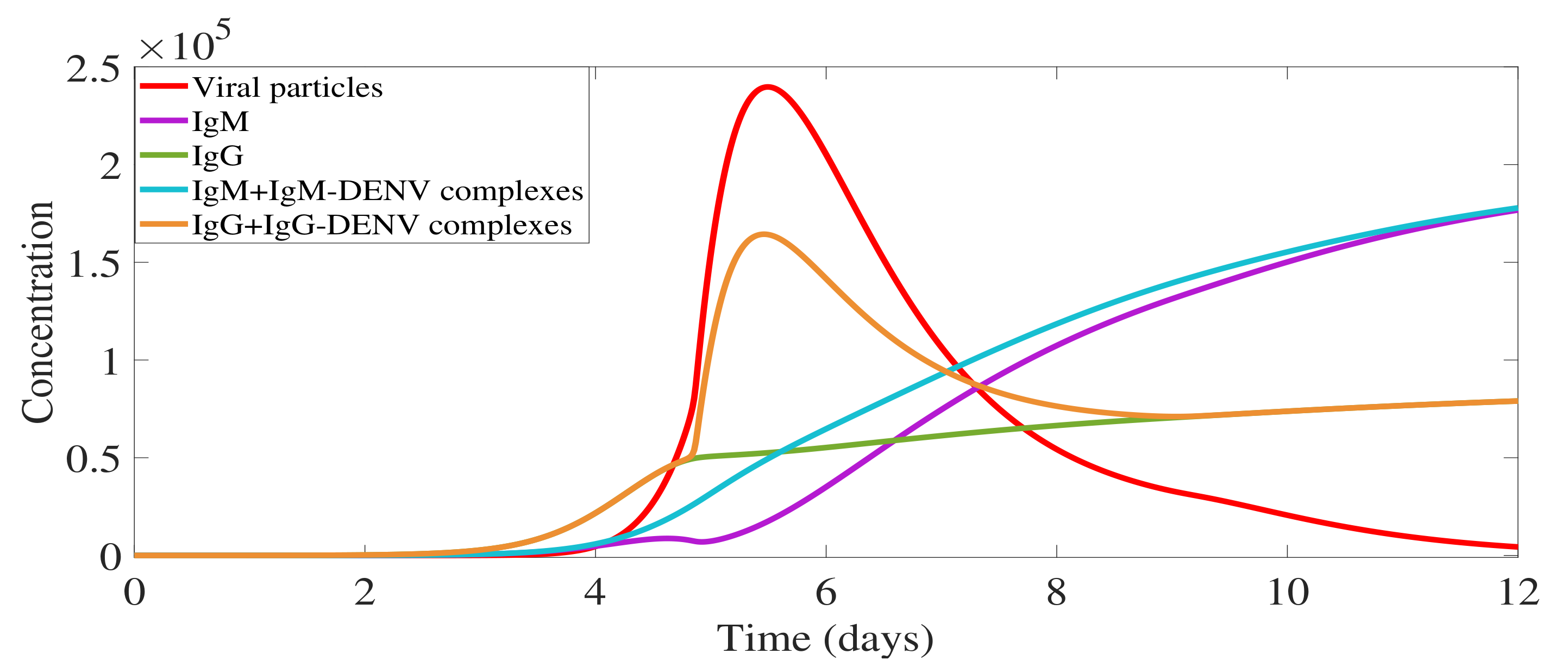
2.1.2. IgM and IgG Antibody Production and Decay and Free Viral Load Dynamics
2.1.3. Antibody-Virus Complexes and Infection Clearance
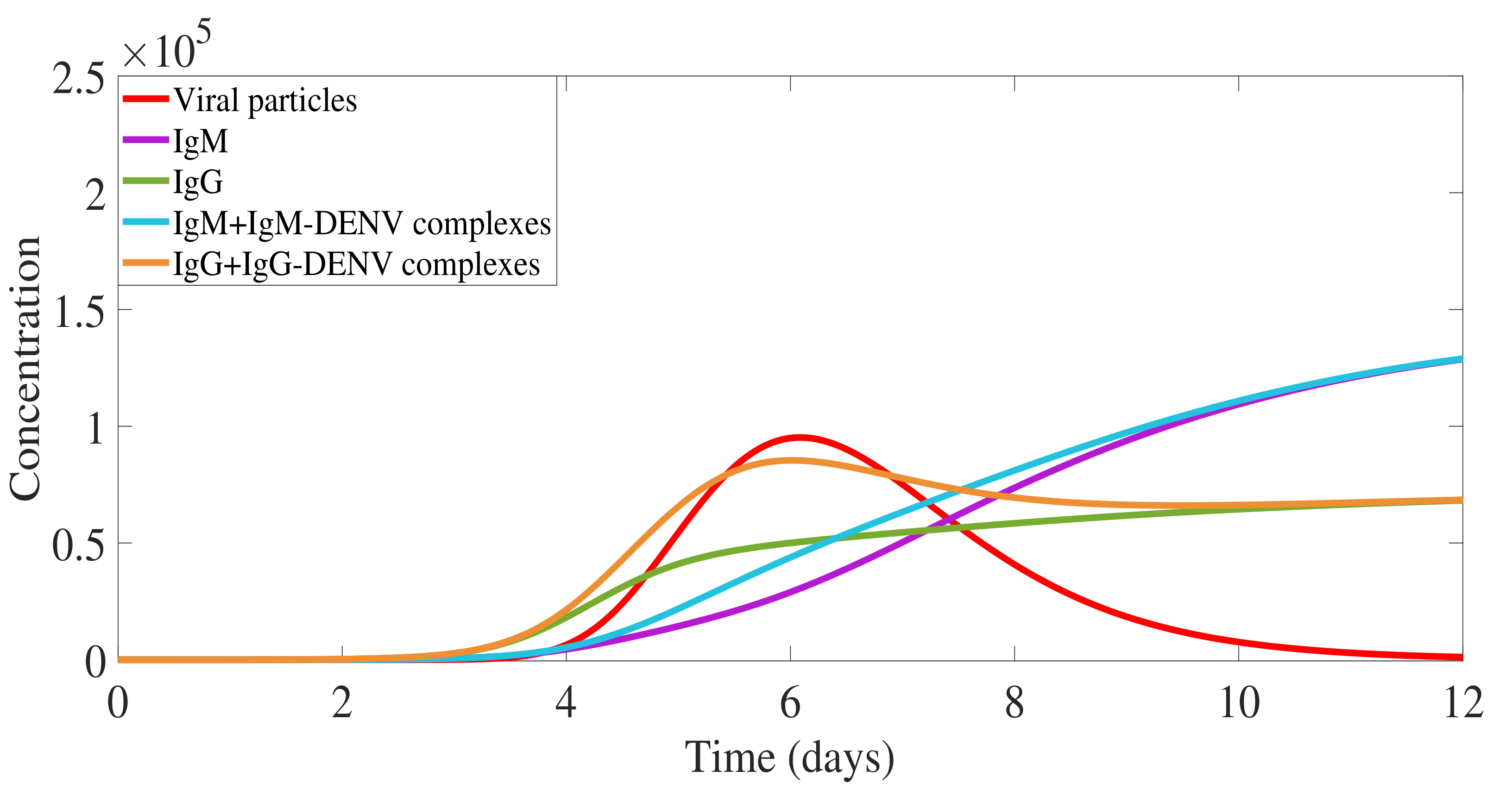
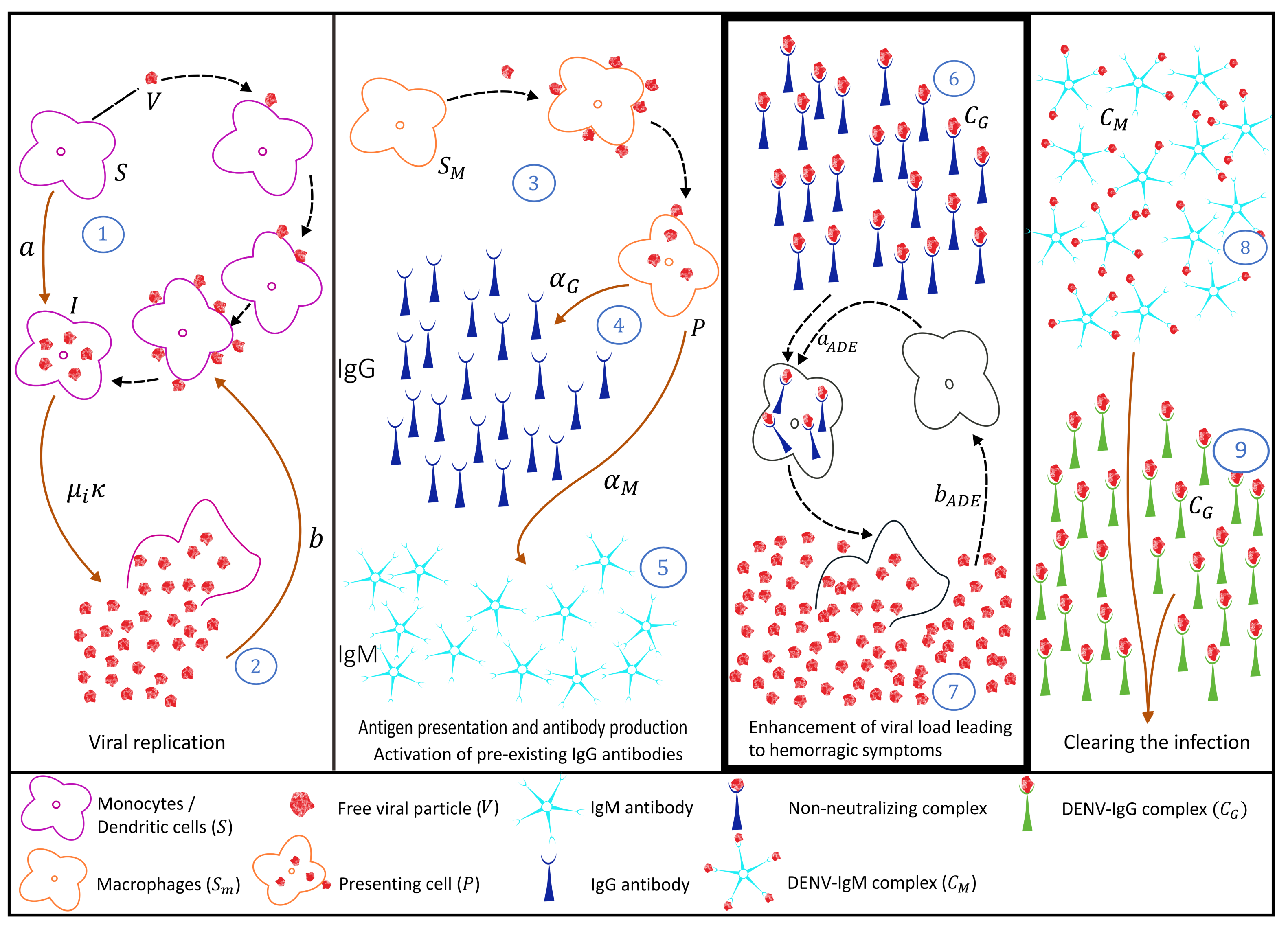
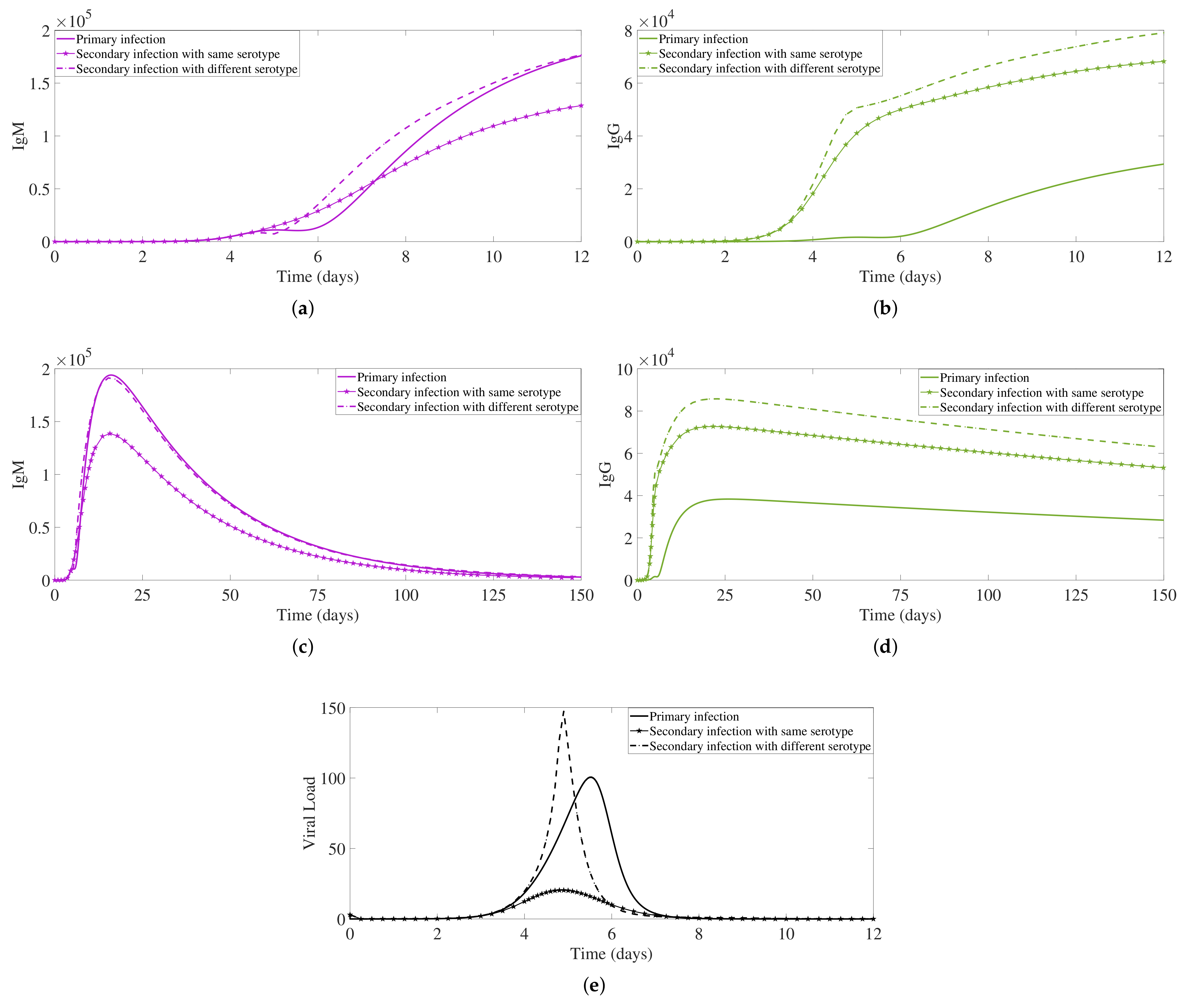
2.2. Secondary Dengue Infection Model with a Homologous Serotype
2.3. Secondary Infection with a Heterologous Serotype
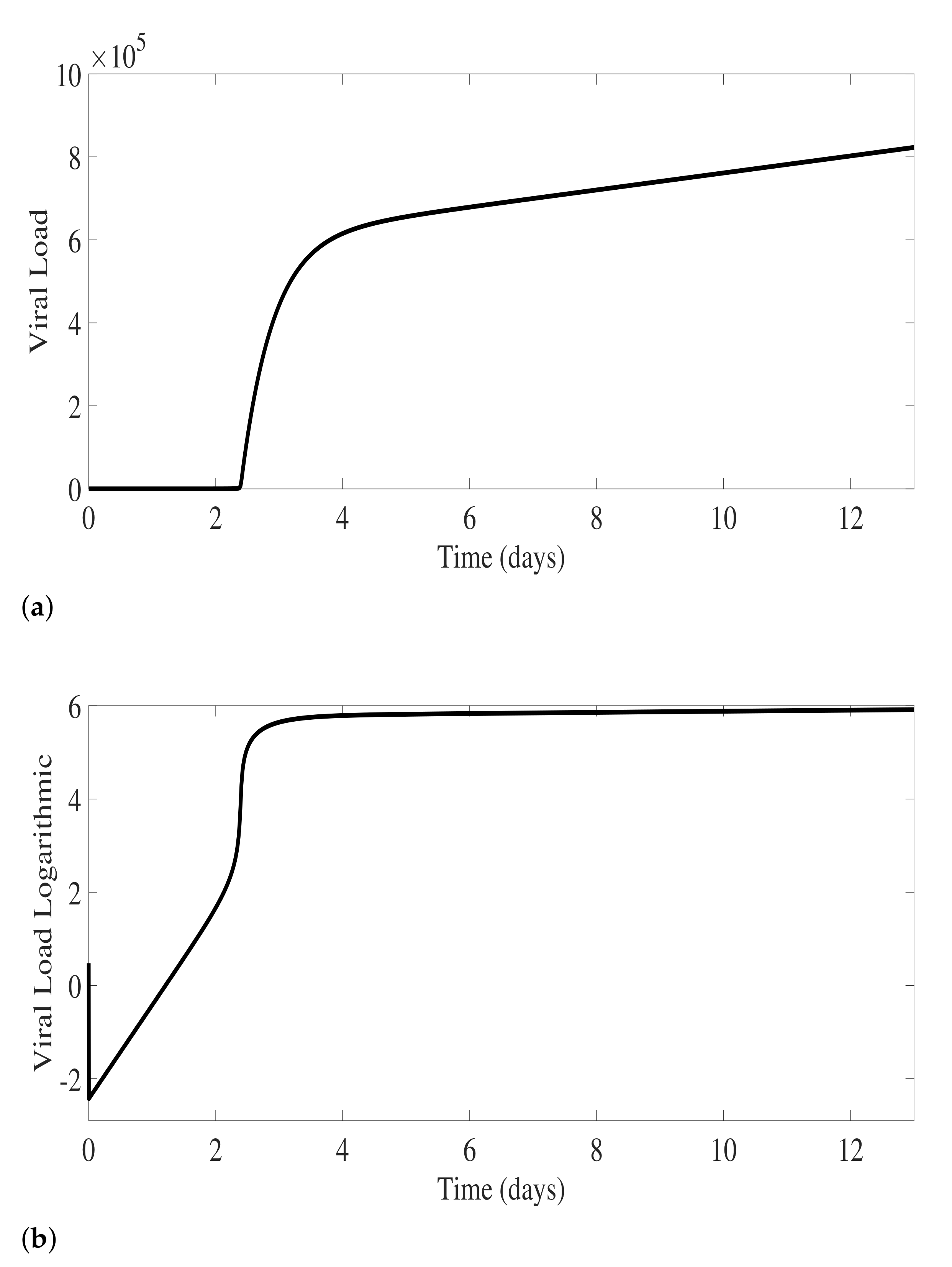
3. Antibody Responses and Viral Load Levels to Explain Disease Symptoms and Severity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue hemorrhagic fever: Two infections and antibody-dependent enhancement, a brief history and personal memoir. Rev. Cuba. Med. Trop. 2002, 54, 3. [Google Scholar]
- Halstead, S.B. Neutralization and antibody-dependent enhancement of dengue viruses. Advances in Virus Research. Adv. Virus Res. 2003, 60, 421–467. [Google Scholar] [PubMed]
- Rothman, A.L. Dengue: Defining protective versus pathologic immunity. J. Clin. Investig. 2004, 113, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Clapham, H.E.; Cummings, D.A.T.; Johansson, M.A. Immune status alters the probability of apparent illness due to dengue virus infection: Evidence from a pooled analysis across multiple cohort and cluster studies. PLoS Negl. Trop. Dis. 2017, 11, e0005926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothman, A.L. Cellular Immunology of Sequential Dengue Virus Infection and Its Role in Disease Pathogenesis; Dengue Virus. Current Topics in Microbiology and Immunology; Rothman, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 338. [Google Scholar] [CrossRef]
- St. John, A.L.; Rathore, A.P.S. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef]
- Malavige, G.N.; Fernando, S.; Fernando, D.J.; Seneviratne, S.L. Dengue Viral Infections. Postgrad Med. J. 2004, 80, 588–601. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, J.E.; Coronel-Ruiz, C. Dengue Disease diagnosis: A puzzle to be solved. Rev. Fac. Med. 2014, 62, 617–629. [Google Scholar] [CrossRef]
- Innis, B.L.; Nisalak, A.; Nimmannitya, S.; Kusalerdchariya, S.V.; Chongswasdi, V.; Suntayakorn, S.; Hoke, C.H. An enzyme-linked immunosorbent assay to characterize dengue infections where dengue and Japanese encephalitis co-circulate. Am. J. Trop. Med. Hyg. 1989, 40, 418–427. [Google Scholar] [CrossRef]
- Pal, S.; Dauner, A.L.; Mitra, I.; Forshey, B.M.; Garcia, P.; Morrison, A.C.; Wu, S.J.L. Evaluation of dengue NS1 antigen rapid tests and ELISA kits using clinical samples. PLoS ONE 2014, 9, e113411. [Google Scholar]
- Changal, K.H.; Raina, A.H.; Raina, A.; Raina, M.; Bashir, R.; Latief, M.; Mir, T.; Changal, Q.H. Differentiating secondary from primary dengue using IgG to IgM ratio in early dengue: An observation hospital bases clinico-serogical study from North India. BMC Infect. Dis. 2016, 16, 715. [Google Scholar] [CrossRef] [Green Version]
- Betts, J.G.; Young, K.A.; Wise, J.A.; Johnson, E.; Poe, B.; Kruse, D.H.; Korol, O.; Johnson, J.E.; Womble, M.; DeSaix, P. Anatomy and Physiology; OpenStax: Houston, TX, USA, 2019; pp. 975–1018. Available online: https://opentextbc.ca/anatomyandphysiologyopenstax/chapter/the-adaptive-immune-response-b-lymphocytes-and-antibodies/ (accessed on 19 August 2021).
- Sathe, A.; Cusick, J.K. Biochemistry, Immunoglobulin M; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Halstead, S.B. Dengue antibody-dependent enhancement: Knowns and unknown. Microbiol. Spectr. 2014, 6, 1–18. [Google Scholar]
- Roehrig, J.T. Antigenic structure of flavirus proteins. Adv. Virus Res. 2003, 59, 141–175. [Google Scholar]
- Billings, L.; Fiorillo, A.; Schwartz, I.B. Vaccination in disease models with antibody-dependent enhancement. Math. Biosci. 2008, 211, 265–281. [Google Scholar] [CrossRef]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martinez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Sirijitt, V.; Wannee, L.; Chunya, P.; Carolyn, E.; Thaneeya, D.; Sunpetchuda, S.; et al. Cross-Reacting Antibodies Enhance Dengue Virus Infection in Humans. Science 2010, 328, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Boonnak, K.; Damback, K.M.; Donofrio, G.C.; Tassaneetrithep, B.; Marovich, M.A. Cell Type and Host Genetic Polymorphisms Influence Antibody-Dependent Enhancement of Dengue Virus Infection. J. Virol. 2011, 85, 1671–1683. [Google Scholar] [CrossRef] [Green Version]
- Bournazos, S.; Gupta, A.; Ravetch, J.V. The role of IgG Fc receptors in antibody-dependent enhancement. Nat. Rev. Immunol. 2020, 20, 633–643. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A.; et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef]
- Thomas, L.; Verlaeten, O.; Cabie, A.; Kaidomar, S.; Moravie, V. Influence of the dengue serotype, previous dengue infection, and plasma viral load on clinical presentation and outcome during a dengue-2 and dengue-4 co-epidemic. Am. J. Trop. Med. Hyg. 2008, 78, 990–998. [Google Scholar] [CrossRef]
- Wang, W.K.; Chao, D.Y.; Kao, C.L.; Wu, H.C.; Liu, Y.C.; Li, C.M.; Lin, S.C.; Ho, S.T.; Huang, J.H.; King, C.C. High levels of plasma dengue viral load during defervescence in patients with dengue hemorrhagic fever: Implications for pathogenesis. Virology 2003, 305, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilarde, A.O.; Turchi, M.D.; Siqueira, J.B., Jr.; Feres, V.C.R.; Rocha, B.; Levi, J.E.; Souza, V.A.U.F.; Boas, L.S.V.; Pannuti, C.S.; Martelli, C.M.T. Dengue and dengue hemorrhagic fever among adults: Clinical outcomes related to viremia, serotypes, and antibody response. J. Infect. Dis. 2008, 197, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadinegoro, S.R.; Arredondo-García, J.L.; Capeding, M.R.; Deseda, C.; Chotpitayasunondh, T.; Dietze, R.; Ismail, H.I.H.J.M.; Reynales, H.; Limkittikul, K.; Rivera-Medina, D.M.; et al. Efficacy and long term safety of a dengue vaccine in regions of endemic disease. N. Engl. J. Med. 2015, 373, 1995–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswal, S.; Reynales, H.; Saez-Llorens, X.; Lopez, P.; Borja-Tabora, C.; Kosalaraksa, P.; Sirivichayakul, C.; Watanaveeradej, V.; Rivera, L.; Espinoza, F.; et al. Efficacy of a Tetravalent Dengue Vaccine in Healthy Children and Adolescents. N. Engl. J. Med. 2019, 381, 2009–2019. [Google Scholar] [CrossRef]
- Biswal, S.; Borja-Tabora, C.; Martinez Vargas, L.; Velásquez, H.; Alera, M.T.; Sierr, V.; Rodriguez-Arenales, E.J.; Yu, D.; Wickramasinghe, V.P.; Moreira, E.D., Jr.; et al. Efficacy of a tetravalent dengue vaccine in healthy children aged 4–16 years: A randomised, placebo-controlled, phase 3 trial. Lancet 2020, 395, 1423–1433. [Google Scholar] [CrossRef]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The risks behind Dengvaxia recommendation. Lancet Infect. Dis. 2016, 16, 882–883. [Google Scholar] [CrossRef]
- Aguiar, M.; Stollenwerk, N. Dengvaxia: Age as surrogate for serostatus. Lancet Infect. 2018, 18, 245. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, M.; Stollenwerk, N. Dengvaxia efficacy dependency on serostatus: A closer look at more recent data. Clin. Infect. Dis. 2018, 66, 641–642. [Google Scholar] [CrossRef] [Green Version]
- Wahala, W.M.P.B.; De Silva, A.M. The human antibody response to dengue virus infection. Viruses 2011, 3, 2374–2395. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, M.; Stollenwerk, N. The impact of serotype cross-protection on vaccine trials: DENVax as a case study. Vaccines 2020, 8, 674. [Google Scholar] [CrossRef]
- White, L.J.; Young, E.F.; Stoops, M.J.; Henein, S.R.; Adams, E.C.; Baric, R.S.; de Silva, A.M. Defining levels of dengue virus serotype-specific neutralizing antibodies induced by a live attenuated tetravalent dengue vaccine (TAK-003). PLoS Negl. Trop. Dis. 2021, 15, e0009258. [Google Scholar] [CrossRef]
- Aguiar, M.; Ballesteros, S.; Kooi, B.W.; Stollenwerk, N. The role of seasonality and import in a minimalistic multi-strain dengue model capturing differences between primary and secondary infections: Complex dynamics and its implications for data analysis. J. Theor. Biol. 2011, 289, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Rocha, R.; Pessanha, J.E.M.; Mateus, L.; Stollenwerk, N. Carnival or football, is there a real risk for acquiring dengue fever in Brazil during holidays seasons. Nat. Sci. Rep. 2015, 5, 8462. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B.; Katzelnick, L.C.; Russell, P.K.; Markoff, L.; Aguiar, M.; Dans, L.R.; Dans, A.L. Ethics of a Partially Effective Dengue Vaccine: Lessons from the Philippines. Vaccine 2020, 38, 5572–5576. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. Modeling the impact of the newly licensed dengue vaccine in endemic countries. PLoS Negl. Trop. Dis. 2016, 10, e0005179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, M.; Stollenwerk, N. Mathematical models of dengue fever epidemiology: Multi-strain dynamics, immunological aspects associated to disease severity and vaccines. Commun. Biomath. Sci. 2017, 1, 1–12. [Google Scholar] [CrossRef]
- Aguiar, M. Dengue vaccination: A more ethical approach is needed. Lancet 2018, 391, 1769–1770. [Google Scholar] [CrossRef]
- Aguiar, M.; Halstead, S.B.; Stollenwerk, N. Consider stopping dengvaxia administration without immunological screening. Expert Rev. Vaccines 2017, 16, 301–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuraini, N.; Tasman, H.; Soewono, E.; Sidarto, K.A. A with-in host dengue infection model with immune response. Math. Comp. Model. 2009, 41, 1148–1155. [Google Scholar] [CrossRef]
- Ansari, N.A.; Hesaaraki, M. A within host dengue infection model with immune response and Beddington-DeAngelis incidence rate. Appl. Math. 2012, 3, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Clapham, H.E.; Tricou, V.; Chau, N.V.V.; Simmons, C.P.; Ferguson, N.M. Within-Host viral dynamics of dengue serotype 1 infection. J. R. Soc. Interface 2014, 11, 20140094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gujarati, T.P.; Ambika, G. Virus antibody dynamics in primary and secondary dengue infections. J. Math. Biol. 2014, 69, 1773–1800. [Google Scholar] [CrossRef] [Green Version]
- Clapham, H.E.; Quyen, T.H.; Kien, D.T.H.; Dorigatti, I.; Simmons, C.P.; Ferguson, N.M. Modelling Virus and Antibody Dynamics during Dengue Virus Infection Suggests a Role for Antibody in Virus Clearance. PLoS Comput. Biol. 2016, 12, e1004951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Shachar, R.; Koelle, K. Minimal within-host dengue models highlight the spesific roles of the immune response in primary and secondary infections. J. R. Soc. Interface 2014, 12. [Google Scholar] [CrossRef]
- Smit, J.M.; Moesker, B.; Rodentiuis-Zybert, I.; Wilschul, J. Flavirus cell entry and membrane fusion. Viruses 2011, 3, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeway, C.A., Jr.; Travers, P.; Walport, M. Immunobiology: The Immune System in Health and Disease, 5th ed.; The Distribution and Functions of Immunoglobulin Isotypes; Garland Science: New York, NY, USA, 2001. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK27162/ (accessed on 19 August 2021).
- Diamond, M.S. Evasion of innate and adaptive immunity by flaviviruses. Immunol. Cell Biol. 2003, 81, 196–206. [Google Scholar] [CrossRef]
- White Cell Count. Available online: http://www.gpnotebook.com/simplepagr.cfm?ID=1013645382 (accessed on 23 August 2021).
- Cologna, R.; Rico-Hesse, R. American Genotype structures Decrease Dengue Virus Output from Human Monocytes and Dendric Cells. J. Virol. 2003, 77, 3929–3938. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhu, B.; Fu, Z.F.; Chen, H.; Cao, S. Immune evasion strategies of flaviviruses. Vacine 2013, 31, 46–471. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dengue Modeling Framework Parameters | |||
|---|---|---|---|
| Parameters | Parameter Values | Biological Meaning | References |
| 600 | constant target cell production (monocytes/dendritic cells) per day | [45,52] | |
| 300 | constant target cell production (macrophages) per day | [45,52] | |
| lifespan of susceptible target cells in days | [48] | ||
| 2 | lifespan of infected cells (monocytes/dendritic cells) per day | modeled | |
| lifespan of presenting cells per day | modeled | ||
| infection rate of susceptible target cells per viral particle per day | modeled | ||
| removal rate of viral particles during the infection of target cells | modeled | ||
| 50 | viral replication factor | [53] | |
| 10 | reproduction rate of IgM antibody per day | modeled | |
| reproduction rate of IgG antibody per day | modeled | ||
| 2000 | activation rate of pre-existing IgG antibody per day | modeled | |
| antibodies binding rate per day | modeled | ||
| binding rate of free virus with IgM antibody per day | modeled | ||
| binding rate of free virus with IgG antibody per day | modeled | ||
| decay rate of IgM per day | [33] | ||
| decay rate of IgG per day | [33] | ||
| 1 | decay rate of antibody-virus complexes per day | modeled | |
| initial value for target cells (monocytes/dendritic cells) | [48] | ||
| initial value for target cells (macrophages) | [48] | ||
| 3 | initial value for free viral particles upon infection (mosquito bite) | modeled | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sebayang, A.A.; Fahlena, H.; Anam, V.; Knopoff, D.; Stollenwerk, N.; Aguiar, M.; Soewono, E. Modeling Dengue Immune Responses Mediated by Antibodies: A Qualitative Study. Biology 2021, 10, 941. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090941
Sebayang AA, Fahlena H, Anam V, Knopoff D, Stollenwerk N, Aguiar M, Soewono E. Modeling Dengue Immune Responses Mediated by Antibodies: A Qualitative Study. Biology. 2021; 10(9):941. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090941
Chicago/Turabian StyleSebayang, Afrina Andriani, Hilda Fahlena, Vizda Anam, Damián Knopoff, Nico Stollenwerk, Maíra Aguiar, and Edy Soewono. 2021. "Modeling Dengue Immune Responses Mediated by Antibodies: A Qualitative Study" Biology 10, no. 9: 941. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10090941