
Characterization of Microbial Shifts during the Production and Ripening of Raw Ewe Milk-Derived Idiazabal Cheese by High-Throughput Sequencing



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk and Cheese Sampling
2.2. DNA Extraction
2.3. Library Preparation and Sequencing
2.4. Bioinformatic Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of 16S rRNA Gene Sequencing Data
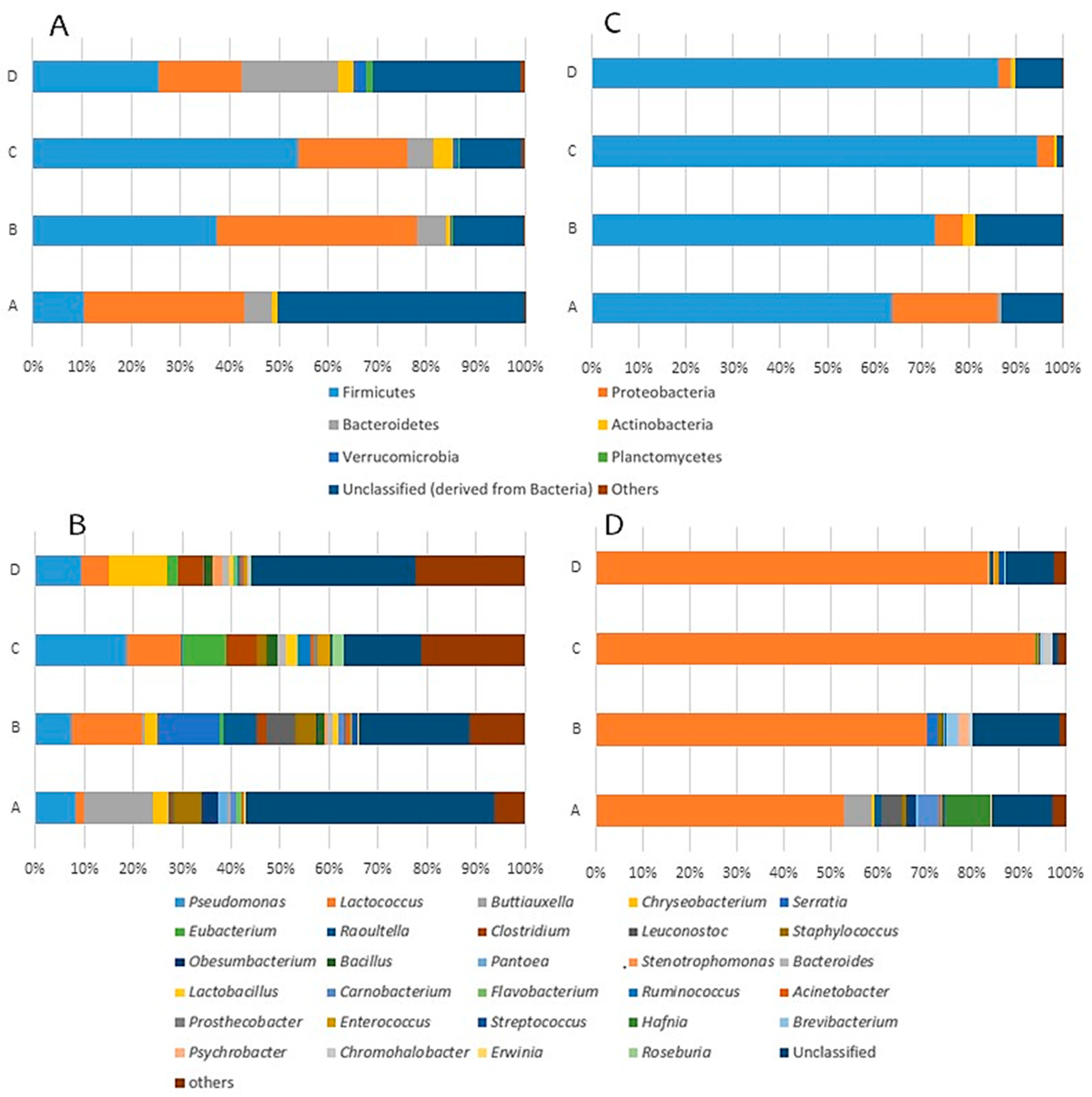
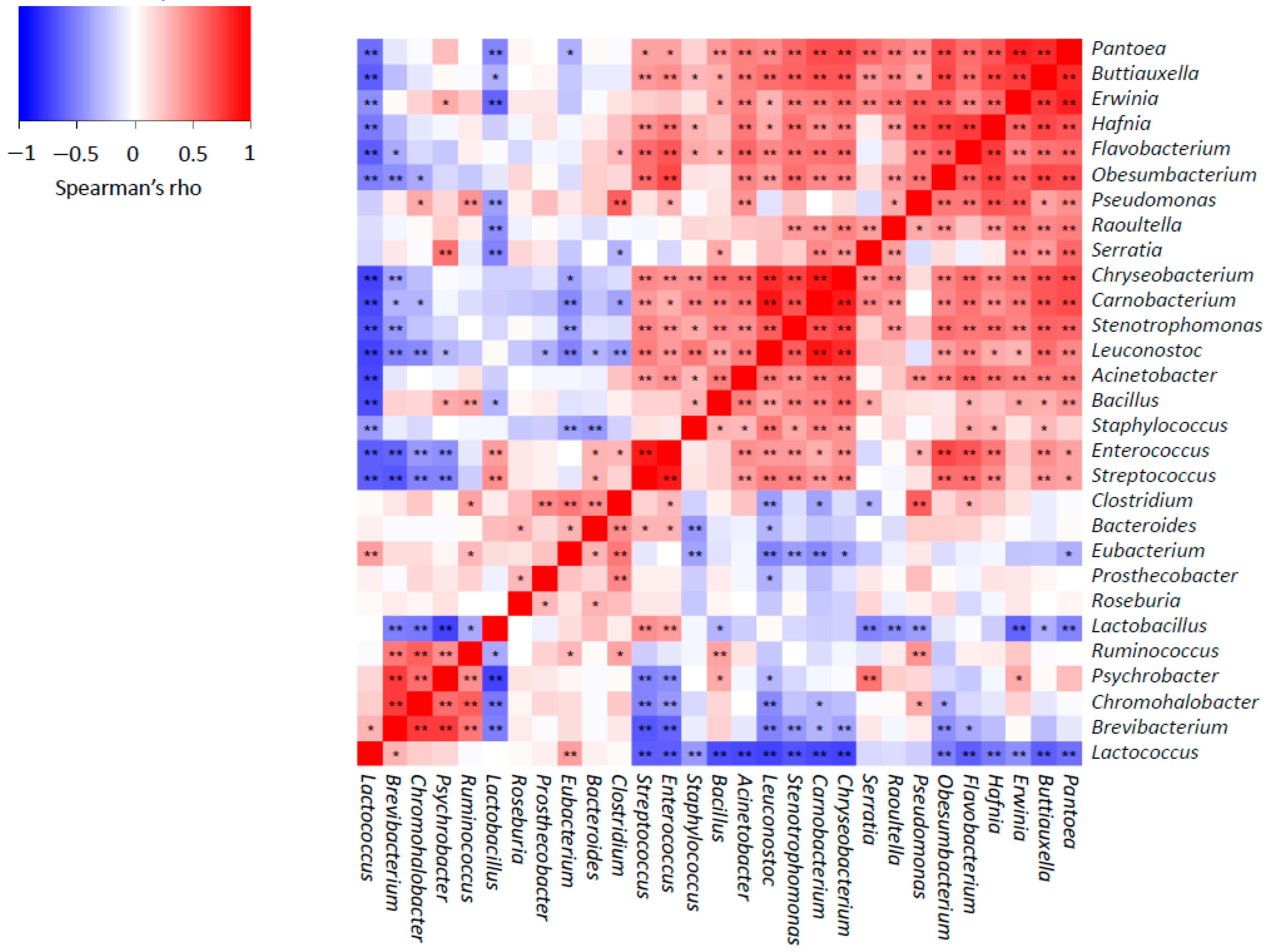
3.2. In-Depth Analysis of Microbial Shifts
3.2.1. Bacterial Composition of the Raw Milk of Ewe
3.2.2. Bacterial Shifts during the Cheese Making Process
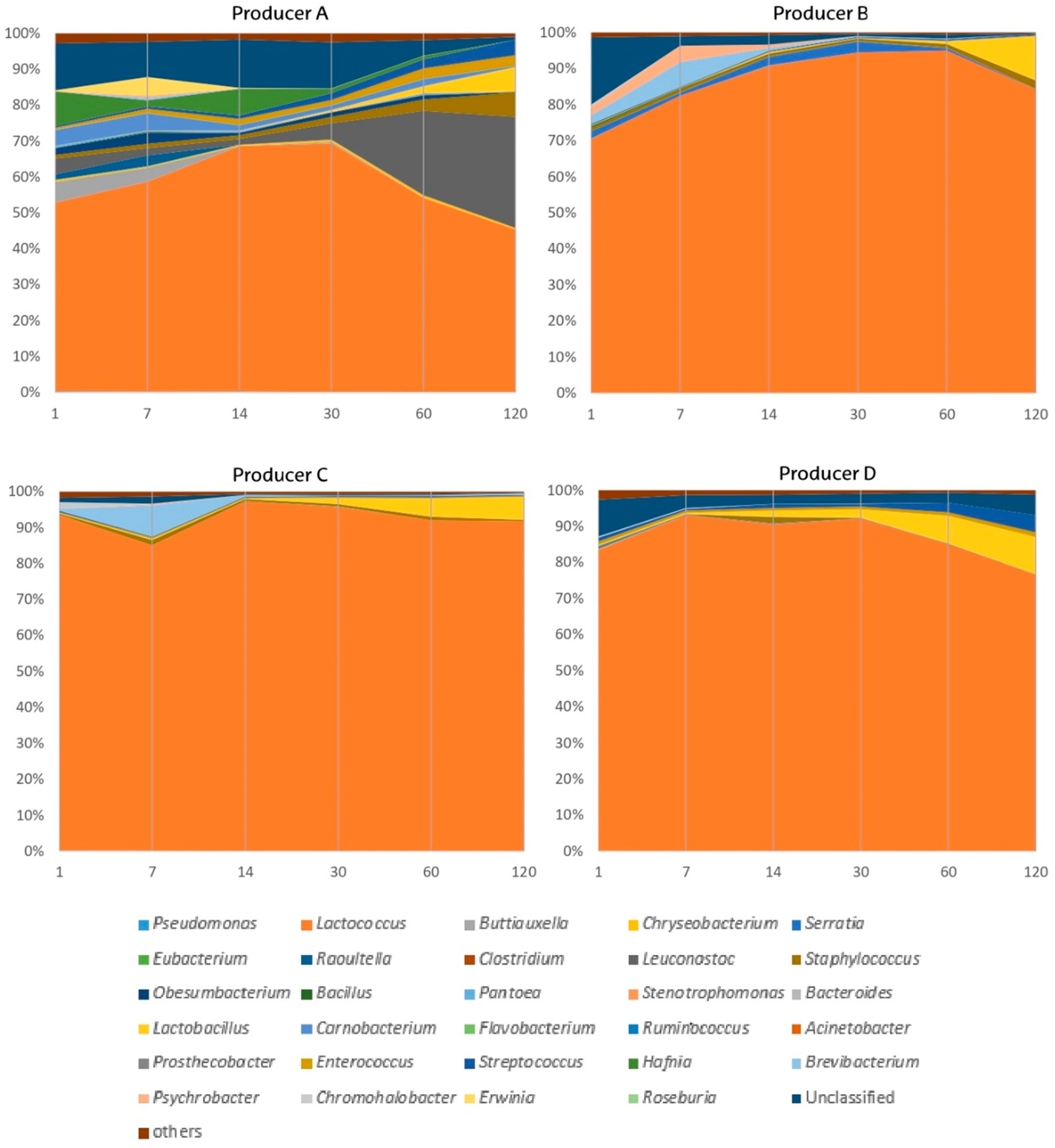
3.2.3. Bacterial Shifts during the Cheese Ripening Process
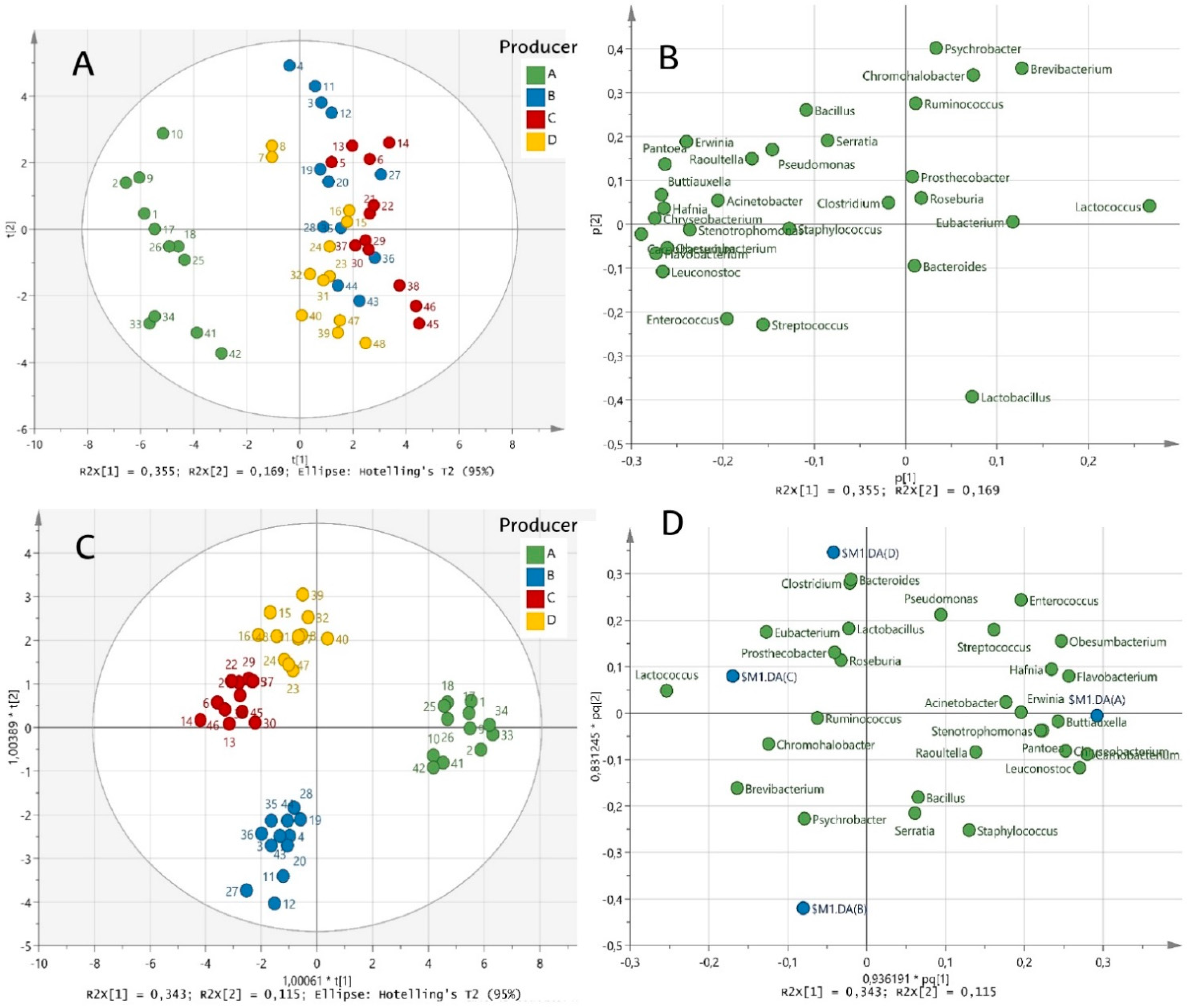
3.3. Overall Effect of Producer and Ripening Time Factors
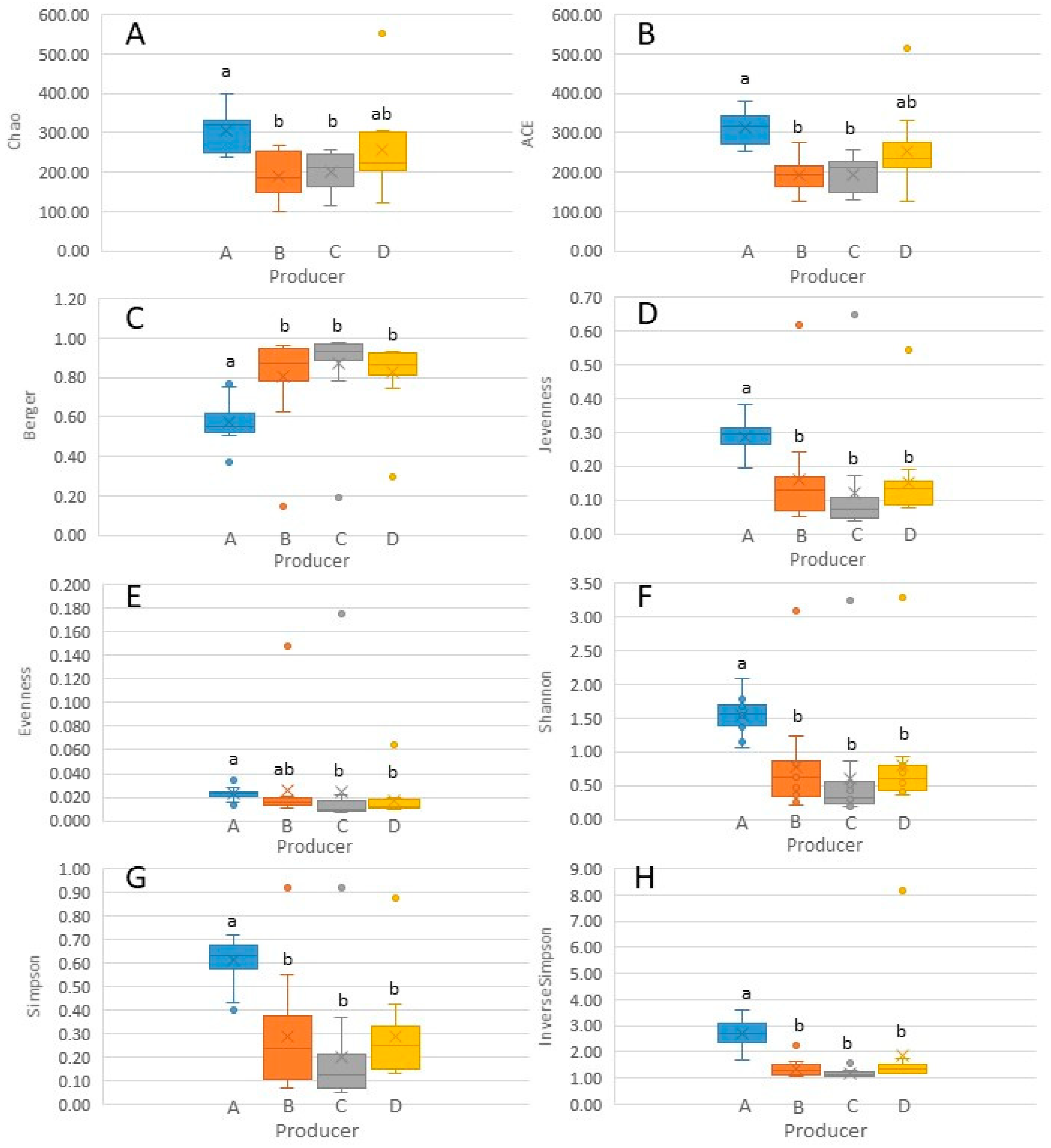
3.4. Alpha and Beta Diversity Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diario Oficial de las Comunidades Europeas. Reglamento (CE) N° 1107/96 de La Comisión de 12 de Junio de 1996 Relativo Al Registro de Las Indicaciones Geográficas y de Las Denominaciones de Origen Con Arreglo Al Procedimiento Establecido En El Artículo 17 Del Reglamento (CEE) No 2081/92 Del Consejo. DOCE 1996, 148, 1–10. [Google Scholar]
- Valdivielso, I.; Bustamante, M.A.; Aldezabal, A.; Amores, G.; Virto, M.; Ruiz De Gordoa, J.C.; De Renobales, M.; Barron, L.J.R. Case Study of a Commercial Sheep Flock under Extensive Mountain Grazing: Pasture Derived Lipid Compounds in Milk and Cheese. Food Chem. 2016, 197, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Boletín Oficial del Estado. Orden del 30 de noviembre por la que se aprueba el Reglamento de la Denominación de Origen «Idiazabal» y su Consejo Regulador. BOE 1993, 289, 34591–34596. [Google Scholar]
- Barron, L.J.R.; Redondo, Y.; Aramburu, M.; Gil, P.; Pérez-Elortondo, F.J.; Albisu, M.; Nájera, A.I.; de Renobales, M.; Fernández-García, E. Volatile Composition and Sensory Properties of Industrially Produced Idiazabal Cheese. Int. Dairy J. 2007, 17, 1401–1414. [Google Scholar] [CrossRef]
- O’Sullivan, O.; Cotter, P.D. Microbiota of Raw Milk and Raw Milk Cheeses. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: San Diego, CA, USA, 2017; Volume 1, pp. 301–316. [Google Scholar] [CrossRef]
- Peláez, C.; Requena, T. Exploiting the Potential of Bacteria in the Cheese Ecosystem. Int. Dairy J. 2005, 15, 831–844. [Google Scholar] [CrossRef]
- Cosentino, S.; Palmas, F. Hygienic Conditions and Microbial Contamination in Six Ewe’s-Milk-Processing Plants in Sardinia, Italy. J. Food Prot. 1997, 60, 283–287. [Google Scholar] [CrossRef]
- Cruciata, M.; Sannino, C.; Ercolini, D.; Scatassa, M.L.; De Filippis, F.; Mancuso, I.; La Storia, A.; Moschetti, G.; Settanni, L. Animal Rennets as Sources of Dairy Lactic Acid Bacteria. Appl. Environ. Microbiol. 2014, 80, 2050–2061. [Google Scholar] [CrossRef] [Green Version]
- Possas, A.; Bonilla-Luque, O.M.; Valero, A. From Cheese-Making to Consumption: Exploring the Microbial Safety of Cheeses through Predictive Microbiology Models. Foods 2021, 10, 355. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The Complex Microbiota of Raw Milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Elortondo, F.J.; Albisu, M.; Barcina, Y. Changes in the Microflora of Idiazabal Cheese with the Addition of Commercial Lactic Starters. Aust. J. Dairy Technol. 1993, 48, 10–14. [Google Scholar]
- Pérez-Elortondo, F.J.; Aldámiz Echobarria, P.; Albisu, M.; Barcina, Y. Indigenous Lactic Acid Bacteria in Idiazabal Ewes’ Milk Cheese. Int. Dairy J. 1998, 8, 725–732. [Google Scholar] [CrossRef]
- Thierry, A.; Pogacic, T.; Weber, M.; Lortal, S. Production of Flavor Compounds by Lactic Acid Bacteria in Fermented Foods. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 314–340. ISBN 9781118868386. [Google Scholar]
- Kamilari, E.; Anagnostopoulos, D.A.; Papademas, P.; Kamilaris, A.; Tsaltas, D. Characterizing Halloumi Cheese’s Bacterial Communities through Metagenomic Analysis. LWT 2020, 126, 109298. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Ning, C.; Chen, L.; Zhao, Y.; Yang, G.; Wang, C.; Chen, N.; Zhang, Z.; Li, S. Impacts of Manufacture Processes and Geographical Regions on the Microbial Profile of Traditional Chinese Cheeses. Food Res. Int. 2021, 148, 110600. [Google Scholar] [CrossRef] [PubMed]
- Niro, S.; Fratianni, A.; Tremonte, P.; Sorrentino, E.; Tipaldi, L.; Panfili, G.; Coppola, R. Innovative Caciocavallo Cheeses Made from a Mixture of Cow Milk with Ewe or Goat Milk. J. Dairy Sci. 2014, 97, 1296–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Liu, F.; Shi, X.; Wang, B.; Li, K.; Li, B.; Zhuge, B. Dynamic Correlations between Microbiota Succession and Flavor Development Involved in the Ripening of Kazak Artisanal Cheese. Food Res. Int. 2018, 105, 733–742. [Google Scholar] [CrossRef]
- Mendia, C.; Ibañez, F.C.; Torre, P.; Barcina, Y. Influence of the Season on Proteolysis and Sensory Characteristics of Idiazabal Cheese. J. Dairy Sci. 2000, 83, 1899–1904. [Google Scholar] [CrossRef]
- Pérez-Elortondo, F.J.; Albisu, M.; Barcina, Y. Physicochemical Properties and Secondary Microflora Variability in the Manufacture and Ripening of Idiazabal Cheese. Lait 1999, 79, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Huang, X.; Yang, B.; Zhang, X.; Han, Y.; Chen, X.-X.; Han, B.-Z. Contrasting the Microbial Community and Metabolic Profile of Three Types of Light-Flavor Daqu. Food Biosci. 2021, 144, 101395. [Google Scholar] [CrossRef]
- Liang, T.; Xie, X.; Zhang, J.; Ding, Y.; Wu, Q. Bacterial Community and Composition of Different Traditional Fermented Dairy Products in China, South Africa, and Sri Lanka by High-Throughput Sequencing of 16S RRNA Genes. LWT 2021, 144, 111209. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Li, D.; Zhang, W.; Jiang, M.; Chen, X.H.; Dong, M.S. Comparative Analysis of the Bacterial Diversity of Chinese Fermented Sausages Using High-Throughput Sequencing. LWT 2021, 150, 111975. [Google Scholar] [CrossRef]
- Biolcati, F.; Ferrocino, I.; Bottero, M.T.; Dalmasso, A. Short Communication: High-Throughput Sequencing Approach to Investigate Italian Artisanal Cheese Production. J. Dairy Sci. 2020, 103, 10015–10021. [Google Scholar] [CrossRef] [PubMed]
- Nero, L.A.; Andretta, M.; Almeida, T.T.; Ferreira, L.R.; Camargo, A.C.; Yamatogi, R.S.; Carvalho, A.F.; Call, D.R. Lactic Microbiota of the Minas Artisanal Cheese Produced in the Serro Region, Minas Gerais, Brazil. LWT 2021, 148, 111698. [Google Scholar] [CrossRef]
- Salazar, J.K.; Carstens, C.K.; Ramachandran, P.; Shazer, A.G.; Narula, S.S.; Reed, E.; Ottesen, A.; Schill, K.M. Metagenomics of Pasteurized and Unpasteurized Gouda Cheese Using Targeted 16S RDNA Sequencing. BMC Microbiol. 2018, 18, 189. [Google Scholar] [CrossRef] [PubMed]
- De Pasquale, I.; Calasso, M.; Mancini, L.; Ercolini, D.; La Storia, A.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Causal Relationship between Microbial Ecology Dynamics and Proteolysis during Manufacture and Ripening of Protected Designation of Origin (PDO) Cheese Canestrato Pugliese. Appl. Environ. Microbiol. 2014, 80, 4085–4094. [Google Scholar] [CrossRef] [Green Version]
- Ercolini, D.; De Filippis, F.; La Storia, A.; Iacono, M. “Remake” by High-Throughput Sequencing of the Microbiota Involved in the Production of Water Buffalo Mozzarella Cheese. Appl. Environ. Microbiol. 2012, 78, 8142–8145. [Google Scholar] [CrossRef] [Green Version]
- Michailidou, S.; Pavlou, E.; Pasentsis, K.; Rhoades, J.; Likotrafiti, E.; Argiriou, A. Microbial Profiles of Greek PDO Cheeses Assessed with Amplicon Metabarcoding. Food Microbiol. 2021, 99, 103836. [Google Scholar] [CrossRef]
- Jiang, Y.; Yu, P.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Shifts in Diversity and Function of Bacterial Community during Manufacture of Rushan. J. Dairy Sci. 2021, 104, 12375–12393. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Z.; Lei, F.; Wang, B.; Jiang, S.; Peng, Q.; Zhang, J.; Shao, Y. Bacterial Diversity in Goat Milk from the Guanzhong Area of China. J. Dairy Sci. 2017, 100, 7812–7824. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Martín-Platero, A.; Maqueda, M.; Valdivia, E.; Martínez-Bueno, M. Biodiversity of the Microbial Community in a Spanish Farmhouse Cheese as Revealed by Culture-Dependent and Culture-Independent Methods. Int. J. Food Microbiol. 2008, 127, 200–208. [Google Scholar] [CrossRef]
- Alegría, Á.; Szczesny, P.; Mayo, B.; Bardowski, J.; Kowalczyk, M. Biodiversity in Oscypek, a Traditional Polish Cheese, Determined by Culture-Dependent and -Independent Approaches. Appl. Environ. Microbiol. 2012, 78, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Masoud, W.; Vogensen, F.K.; Lillevang, S.; Abu Al-Soud, W.; Sørensen, S.J.; Jakobsen, M. The Fate of Indigenous Microbiota, Starter Cultures, Escherichia Coli, Listeria Innocua and Staphylococcus Aureus in Danish Raw Milk and Cheeses Determined by Pyrosequencing and Quantitative Real Time (QRT)-PCR. Int. J. Food Microbiol. 2012, 153, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the Classification of Cultured and Uncultured Bacteria and Archaea Using 16S RRNA Gene Sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Bertani, G.; Levante, A.; Lazzi, C.; Bottari, B.; Gatti, M.; Neviani, E. Dynamics of a Natural Bacterial Community under Technological and Environmental Pressures: The Case of Natural Whey Starter for Parmigiano Reggiano Cheese. Food Res. Int. 2020, 129, 108860. [Google Scholar] [CrossRef] [PubMed]
- De Paula, A.C.L.; Medeiros, J.D.; Fernandes, G.R.; da Silva, V.L.; Diniz, C.G. Microbiome of Industrialized Minas Frescal Cheese Reveals High Prevalence of Putative Bacteria: A Concern in the One Health Context. LWT 2021, 139, 110791. [Google Scholar] [CrossRef]
- Cardinali, F.; Ferrocino, I.; Milanović, V.; Belleggia, L.; Corvaglia, M.R.; Garofalo, C.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Cocolin, L.; et al. Microbial Communities and Volatile Profile of Queijo de Azeitão PDO Cheese, a Traditional Mediterranean Thistle-Curdled Cheese from Portugal. Food Res. Int. 2021, 147, 110537. [Google Scholar] [CrossRef]
- Dimov, S.G.; Gyurova, A.; Zagorchev, L.; Dimitrov, T.; Georgieva-Miteva, D.; Peykov, S. NGS-Based Metagenomic Study of Four Traditional Bulgarian Green Cheeses from Tcherni Vit. LWT 2021, 152, 112278. [Google Scholar] [CrossRef]
- Gaglio, R.; Franciosi, E.; Todaro, A.; Guarcello, R.; Alfeo, V.; Randazzo, C.L.; Settanni, L.; Todaro, M. Addition of Selected Starter/Non-Starter Lactic Acid Bacterial Inoculums to Stabilise PDO Pecorino Siciliano Cheese Production. Food Res. Int. 2020, 136, 109335. [Google Scholar] [CrossRef]
- Endres, C.M.; Castro, Í.M.S.; Trevisol, L.D.; Severo, J.M.; Mann, M.B.; Varela, A.P.M.; Frazzon, A.P.G.; Mayer, F.Q.; Frazzon, J. Molecular Characterization of the Bacterial Communities Present in Sheep’s Milk and Cheese Produced in South Brazilian Region via 16S RRNA Gene Metabarcoding Sequencing. LWT 2021, 147, 111579. [Google Scholar] [CrossRef]
- Esteban-Blanco, C.; Gutiérrez-Gil, B.; Puente-Sánchez, F.; Marina, H.; Tamames, J.; Acedo, A.; Arranz, J.J. Microbiota Characterization of Sheep Milk and Its Association with Somatic Cell Count Using 16s RRNA Gene Sequencing. J. Anim. Breed. Genet. 2020, 137, 73–83. [Google Scholar] [CrossRef]
- Biçer, Y.; Telli, A.E.; Sönmez, G.; Telli, N.; Uçar, G. Comparison of Microbiota and Volatile Organic Compounds in Milk from Different Sheep Breeds. J. Dairy Sci. 2021, 104, 12303–12311. [Google Scholar] [CrossRef]
- Cardinali, F.; Osimani, A.; Taccari, M.; Milanović, V.; Garofalo, C.; Clementi, F.; Polverigiani, S.; Zitti, S.; Raffaelli, N.; Mozzon, M.; et al. Impact of Thistle Rennet from Carlina Acanthifolia All. Subsp. Acanthifolia on Bacterial Diversity and Dynamics of a Specialty Italian Raw Ewes’ Milk Cheese. Int. J. Food Microbiol. 2017, 255, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, M.; Hosseini, S.M.; Ferrocino, I.; Amoozegar, M.A.; Cocolin, L. Molecular Investigation of Bacterial Communities during the Manufacturing and Ripening of Semi-Hard Iranian Liqvan Cheese. Food Microbiol. 2017, 66, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, M.; Chávarri, F.; Santisteban, A.; Ceballos, G.; Hernández, I.; José Miguélez, M.; Aranburu, I.; Barrón, L.J.R.; Virto, M.; De Renobales, M. Coagulating and Lipolytic Activities of Artisanal Lamb Rennet Pastes. J. Dairy Res. 2000, 67, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Erkus, O.; de Jager, V.C.L.; Geene, R.T.C.M.; van Alen-Boerrigter, I.; Hazelwood, L.; van Hijum, S.A.F.T.; Kleerebezem, M.; Smid, E.J. Use of Propidium Monoazide for Selective Profiling of Viable Microbial Cells during Gouda Cheese Ripening. Int. J. Food Microbiol. 2016, 228, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The Metagenomics RAST Server—A Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S RRNA Gene Sequencing for Species and Strain-Level Microbiome Analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 March 2020).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Package “Gplots” Title Various R Programming Tools for Plotting Data [R Package Gplots Version 3.1.1]. Available online: https://cran.r-project.org/package=gplots (accessed on 15 January 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan”. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 15 January 2021).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005; ISBN 92 9059 179 X. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R Language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Fundamentals of Cheese Science, 2nd ed.; Fox, P.F., Guinee, T.P., Cogan, T.M., McSweeney, P.L.H., Eds.; Springer: New York, NY, USA, 2017; ISBN 9781489976819. [Google Scholar]
- Quintana, Á.R.; Seseña, S.; Garzón, A.; Arias, R. Factors Affecting Levels of Airborne Bacteria in Dairy Farms: A Review. Animals 2020, 10, 526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Brom, R.; de Jong, A.; van Engelen, E.; Heuvelink, A.; Vellema, P. Zoonotic Risks of Pathogens from Sheep and Their Milk Borne Transmission. Small Rumin. Res. 2020, 189, 106123. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Wang, N.S. Spoilage of Milk and Dairy Products. In The Microbiological Quality of Food: Foodborne Spoilers; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Duxford, UK, 2017; pp. 151–178. ISBN 9780081005033. [Google Scholar]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Paradh, A.D. Gram-Negative Spoilage Bacteria in Brewing. In Brewing Microbiology: Managing Microbes, Ensuring Quality and Valorising Waste; Hill, A.E., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; pp. 175–194. ISBN 9781782423492. [Google Scholar]
- Staley, J.T. The Genus Prosthecobacter. In The Prokaryotes; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: New York, NY, USA, 1981; pp. 477–479. ISBN 9783662131879. [Google Scholar]
- Suchodolski, J.S. Gastrointestinal Microbiota. In Canine and Feline Gastroenterology; Washabau, R.J., Day, M.J., Eds.; W.B. Saunders: St. Louis, MO, USA, 2012; pp. 32–41. ISBN 9781416036616. [Google Scholar]
- Vataščinová, T.; Pipová, M.; Fraqueza, M.J.R.; Maľa, P.; Dudriková, E.; Drážovská, M.; Lauková, A. Short Communication: Antimicrobial Potential of Lactobacillus plantarum Strains Isolated from Slovak Raw Sheep Milk Cheeses. J. Dairy Sci. 2020, 103, 6900–6903. [Google Scholar] [CrossRef]
- Falardeau, J.; Keeney, K.; Trmčić, A.; Kitts, D.; Wang, S. Farm-to-Fork Profiling of Bacterial Communities Associated with an Artisan Cheese Production Facility. Food Microbiol. 2019, 83, 48–58. [Google Scholar] [CrossRef]
- Bellassi, P.; Rocchetti, G.; Nocetti, M.; Lucini, L.; Masoero, F.; Morelli, L. A Combined Metabolomic and Metagenomic Approach to Discriminate Raw Milk for the Production of Hard Cheese. Foods 2021, 10, 109. [Google Scholar] [CrossRef]
- Yeluri Jonnala, B.R.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Sequencing of the Cheese Microbiome and Its Relevance to Industry. Front. Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef] [Green Version]
- Actor, J.K. Clinical Bacteriology. In Elsevier’s Integrated Review Immunology and Microbiology; Actor, J.K., Ed.; W.B. Saunders: Philadelphia, PA, USA, 2012; pp. 105–120. ISBN 978-0-323-07447-6. [Google Scholar]
- Antonello, V.S.; Dallé, J.; Domingues, G.C.; Ferreira, J.A.S.; Fontoura, M.d.C.Q.; Knapp, F.B. Post-Cesarean Surgical Site Infection Due to Buttiauxella agrestis. Int. J. Infect. Dis. 2014, 22, 65–66. [Google Scholar] [CrossRef] [Green Version]
- Bergogne-Bérézin, E. Bacteria: Acinetobacter. In Encyclopedia of Food Safety; Motarjemi, Y., Ed.; Academic Press: San Diego, CA, USA, 2014; Volume 1, pp. 337–341. ISBN 9780123786128. [Google Scholar]
- Fusco, V.; Abriouel, H.; Benomar, N.; Kabisch, J.; Chieffi, D.; Cho, G.-S.; Franz, C.M.A.P. Opportunistic Food-Borne Pathogens. In Food Safety and Preservation; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 269–306. ISBN 9780128149560. [Google Scholar]
- Guiu, A.; Buendía, B.; Llorca, L.; Gómez Punter, R.M.; Girón, R. Chryseobacterium spp., a New Opportunistic Pathogen Associated with Cystic Fibrosis? Enferm. Infecc. Microbiol. Clin. 2014, 32, 497–501. [Google Scholar] [CrossRef]
- Li, N.; Wang, Y.; You, C.; Ren, J.; Chen, W.; Zheng, H.; Liu, Z. Variation in Raw Milk Microbiota throughout 12 Months and the Impact of Weather Conditions. Sci. Rep. 2018, 8, 2371. [Google Scholar] [CrossRef] [Green Version]
- Morin, A. Pantoea. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 1028–1032. ISBN 9780123847331. [Google Scholar]
- Murphy, E.C.; Mörgelin, M.; Cooney, J.C.; Frick, I.M. Interaction of Bacteroides fragilis and Bacteroides thetaiotaomicron with the Kallikrein-Kinin System. Microbiology 2011, 157, 2094–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titécat, M.; Wallet, F.; Vieillard, M.H.; Courcol, R.J.; Loïez, C. Ruminococcus gnavus: An Unusual Pathogen in Septic Arthritis. Anaerobe 2014, 30, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Irzykowska, L. Flavobacterium spp.—Characteristics, Occurrence, and Toxicity. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 938–942. ISBN 9780123847331. [Google Scholar]
- Appel, T.M.; Quijano-Martínez, N.; De La Cadena, E.; Mojica, M.F.; Villegas, M.V. Microbiological and Clinical Aspects of Raoultella spp. Front. Public Health 2021, 9, 686789. [Google Scholar] [CrossRef] [PubMed]
- Lo, R.; Xue, T.; Weeks, M.; Turner, M.S.; Bansal, N. Inhibition of Bacterial Growth in Sweet Cheese Whey by Carbon Dioxide as Determined by Culture-Independent Community Profiling. Int. J. Food Microbiol. 2016, 217, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Morandi, S.; Battelli, G.; Silvetti, T.; Tringali, S.; Nunziata, L.; Villa, A.; Acquistapace, A.; Brasca, M. Impact of Salting and Ripening Temperatures on Late Blowing Defect in Valtellina Casera PDO Cheese. Food Control 2021, 120, 107508. [Google Scholar] [CrossRef]
- Fotou, K.; Tzora, A.; Voidarou, C.; Alexopoulos, A.; Plessas, S.; Avgeris, I.; Bezirtzoglou, E.; Akrida-Demertzi, K.; Demertzis, P.G. Isolation of Microbial Pathogens of Subclinical Mastitis from Raw Sheep’s Milk of Epirus (Greece) and Their Role in Its Hygiene. Anaerobe 2011, 17, 315–319. [Google Scholar] [CrossRef]
- Fuka, M.M.; Engel, M.; Skelin, A.; Redžepović, S.; Schloter, M. Bacterial Communities Associated with the Production of Artisanal Istrian Cheese. Int. J. Food Microbiol. 2010, 142, 19–24. [Google Scholar] [CrossRef]
- Ruvalcaba-Gómez, J.M.; Delgado-Macuil, R.J.; Zelaya-Molina, L.X.; Maya-Lucas, O.; Ruesga-Gutiérrez, E.; Anaya-Esparza, L.M.; de la Mora, Z.V.; López-De la Mora, D.A.; Arteaga-Garibay, R.I. Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput Dna Sequencing. Microorganisms 2021, 9, 24. [Google Scholar] [CrossRef]
- Bowman, J.P. The Genus Psychrobacter. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 920–930. ISBN 9780387307466. [Google Scholar]
- Shweta, F.N.U.; Gurram, P.R.; O’Horo, J.C.; Khalil, S. Brevibacterium Species: An Emerging Opportunistic Cause of Bloodstream Infections. Mayo Clin. Proc. 2021, 96, 1093–1094. [Google Scholar] [CrossRef]
- Yin, Z.; Yuan, C.; Du, Y.; Yang, P.; Qian, C.; Wei, Y.; Zhang, S.; Huang, D.; Liu, B. Comparative Genomic Analysis of the Hafnia Genus Reveals an Explicit Evolutionary Relationship between the Species Alvei and Paralvei and Provides Insights into Pathogenicity. BMC Genom. 2019, 20, 768. [Google Scholar] [CrossRef] [Green Version]
- Anast, J.M.; Dzieciol, M.; Schultz, D.L.; Wagner, M.; Mann, E.; Schmitz-Esser, S. Brevibacterium from Austrian Hard Cheese Harbor a Putative Histamine Catabolism Pathway and a Plasmid for Adaptation to the Cheese Environment. Sci. Rep. 2019, 9, 6164. [Google Scholar] [CrossRef] [PubMed]
- Ozturkoglu-Budak, S.; Wiebenga, A.; Bron, P.A.; de Vries, R.P. Protease and Lipase Activities of Fungal and Bacterial Strains Derived from an Artisanal Raw Ewe’s Milk Cheese. Int. J. Food Microbiol. 2016, 237, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Palmer, J.; Teh, K.H.; Flint, S. Identification and Selection of Heat-Stable Protease and Lipase-Producing Psychrotrophic Bacteria from Fresh and Chilled Raw Milk during up to Five Days Storage. LWT 2020, 134, 110165. [Google Scholar] [CrossRef]
- De Pasquale, I.; Di Cagno, R.; Buchin, S.; De Angelis, M.; Gobbetti, M. Spatial Distribution of the Metabolically Active Microbiota within Italian PDO Ewes’ Milk Cheeses. PLoS ONE 2016, 11, e0153213. [Google Scholar] [CrossRef] [Green Version]
- Deetae, P.; Bonnarme, P.; Spinnler, H.E.; Helinck, S. Production of Volatile Aroma Compounds by Bacterial Strains Isolated from Different Surface-Ripened French Cheeses. Appl. Microbiol. Biotechnol. 2007, 76, 1161–1171. [Google Scholar] [CrossRef]
- Irlinger, F.; In Yung, S.A.Y.; Sarthou, A.S.; Delbès-Paus, C.; Montel, M.C.; Coton, E.; Coton, M.; Helinck, S. Ecological and Aromatic Impact of Two Gram-Negative Bacteria (Psychrobacter celer and Hafnia alvei) Inoculated as Part of the Whole Microbial Community of an Experimental Smear Soft Cheese. Int. J. Food Microbiol. 2012, 153, 332–338. [Google Scholar] [CrossRef]
- Quillaguamán, J.; Delgado, O.; Mattiasson, B.; Hatti-Kaul, R. Chromohalobacter sarecensis sp. Nov., a Psychrotolerant Moderate Halophile Isolated from the Saline Andean Region of Bolivia. Int. J. Syst. Evol. Microbiol. 2004, 54, 1921–1926. [Google Scholar] [CrossRef]
- Fuka, M.M.; Wallisch, S.; Engel, M.; Welzl, G.; Havranek, J.; Schloter, M. Dynamics of Bacterial Communities during the Ripening Process of Different Croatian Cheese Types Derived from Raw Ewe’s Milk Cheeses. PLoS ONE. 2013, 8, e80734. [Google Scholar] [CrossRef] [Green Version]
- García-López, M.L.; Santos, J.A.; Otero, A.; Rodríguez-Calleja, J.M. Psychrobacter. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 261–268. ISBN 9780123847331. [Google Scholar]
- Hernández, I.; Barrón, L.J.R.; Virto, M.; Pérez-Elortondo, F.J.; Flanagan, C.; Rozas, U.; Nájera, A.I.; Albisu, M.; Vicente, M.S.; de Renobales, M. Lipolysis, Proteolysis and Sensory Properties of Ewe’s Raw Milk Cheese (Idiazabal) Made with Lipase Addition. Food Chem. 2009, 116, 158–166. [Google Scholar] [CrossRef]
- Gil, P.F.; Conde, S.; Albisu, M.; Pérez-Elortondo, F.J.; Etayo, I.; Virto, M.; De Renobales, M. Hygienic Quality of Ewes’ Milk Cheeses Manufactured with Artisan-Produced Lamb Rennet Pastes. J. Dairy Res. 2007, 74, 329–335. [Google Scholar] [CrossRef]
- Etayo, I.; Pérez Elortondo, F.J.; Gil, P.F.; Albisu, M.; Virto, M.; Conde, S.; Rodriguez Barron, L.J.; Nájera, A.I.; Gómez-Hidalgo, M.E.; Delgado, C.; et al. Hygienic Quality, Lipolysis and Sensory Properties of Spanish Protected Designation of Origin Ewe’s Milk Cheeses Manufactured with Lamb Rennet Paste. Lait 2006, 86, 415–434. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Facility-Specific “House” Microbiome Drives Microbial Landscapes of Artisan Cheesemaking Plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium Review: Interaction of Starter Cultures and Nonstarter Lactic Acid Bacteria in the Cheese Environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef] [PubMed]
- Frantzen, C.A.; Kot, W.; Pedersen, T.B.; Ardö, Y.M.; Broadbent, J.R.; Neve, H.; Hansen, L.H.; Dal Bello, F.; Østlie, H.M.; Kleppen, H.P.; et al. Genomic Characterization of Dairy Associated Leuconostoc Species and Diversity of Leuconostocs in Undefined Mixed Mesophilic Starter Cultures. Front. Microbiol. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and Cons for Using Non-Starter Lactic Acid Bacteria (NSLAB) as Secondary/Adjunct Starters for Cheese Ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Li, S.; Li, Y.; Du, Z.; Li, B.; Liu, Y.; Gao, Y.; Zhang, Y.; Zhang, K.; Wang, Q.; Lu, S.; et al. Impact of NSLAB on Kazakh Cheese Flavor. Food Res. Int. 2021, 144, 110315. [Google Scholar] [CrossRef]
- Mohamaden, W.I.; Zhen-fen, Z.; Hegab, I.M.; Shang-li, S. Experimental Infection in Mice with Erwinia persicina. Microb. Pathog. 2019, 130, 38–43. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-Throughput Sequencing for Detection of Subpopulations of Bacteria Not Previously Associated with Artisanal Cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef] [Green Version]
- Sádecká, J.; Šaková, N.; Pangallo, D.; Koreňová, J.; Kolek, E.; Puškárová, A.; Bučková, M.; Valík, L.; Kuchta, T. Microbial Diversity and Volatile Odour-Active Compounds of Barrelled Ewes’ Cheese as an Intermediate Product That Determines the Quality of Winter Bryndza Cheese. LWT 2016, 70, 237–244. [Google Scholar] [CrossRef]
- D’Amico, D.J.; Donnelly, C.W. Growth and Survival of Microbial Pathogens in Cheese. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: San Diego, CA, USA, 2017; Volume 1, pp. 573–594. ISBN 9780122636530. [Google Scholar]
- Gontijo, M.T.P.; Silva, J.d.S.; Vidigal, P.M.P.; Martin, J.G.P. Phylogenetic Distribution of the Bacteriocin Repertoire of Lactic Acid Bacteria Species Associated with Artisanal Cheese. Food Res. Int. 2020, 128, 108783. [Google Scholar] [CrossRef]
- Kaya, H.I.; Simsek, O. Characterization of Pathogen-Specific Bacteriocins from Lactic Acid Bacteria and Their Application within Cocktail against Pathogens in Milk. LWT 2019, 115, 108464. [Google Scholar] [CrossRef]
- Macedo, A.C.; Malcata, F.X.; Oliveira, J.C. Effect of Production Factors and Ripening Conditions on the Characteristics of Serra Cheese. Int. J. Food Sci. Technol. 1997, 32, 501–511. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Producer | Milk/Cheese Ripening Time (Day) | Sample ID | Bacterial Diversity | |||||
|---|---|---|---|---|---|---|---|---|
| Sequences | OTUs | Mbp Count | Phyla | Families | Genera | |||
| A | Milk | MA | 66,989 | 8450 | 2.70 | 8 | 103 | 221 |
| 1 | 1 | 321,148 | 21,102 | 6.84 | 10 | 102 | 244 | |
| 2 | 331,816 | 17,948 | 5.96 | 10 | 107 | 242 | ||
| 7 | 9 | 238,804 | 12,412 | 4.21 | 10 | 82 | 182 | |
| 10 | 251,225 | 13,472 | 4.55 | 10 | 92 | 203 | ||
| 14 | 17 | 263,231 | 13,305 | 4.51 | 10 | 81 | 176 | |
| 18 | 334,305 | 16,504 | 5.95 | 11 | 90 | 209 | ||
| 30 | 25 | 269,207 | 14,266 | 5.18 | 8 | 89 | 195 | |
| 26 | 280,285 | 13,701 | 4.65 | 8 | 84 | 191 | ||
| 60 | 33 | 330,729 | 15,810 | 5.50 | 10 | 79 | 179 | |
| 34 | 341,419 | 16,324 | 5.92 | 10 | 79 | 183 | ||
| 120 | 41 | 370,181 | 22,557 | 7.61 | 14 | 83 | 187 | |
| 42 | 332,471 | 16,882 | 6.15 | 12 | 78 | 170 | ||
| B | Milk | MB | 6092 | 2504 | 0.769 | 10 | 66 | 136 |
| 1 | 3 | 257,407 | 12,967 | 4.39 | 9 | 90 | 178 | |
| 4 | 184,670 | 10,745 | 3.66 | 7 | 80 | 158 | ||
| 7 | 11 | 160,436 | 9174 | 3.15 | 7 | 70 | 124 | |
| 12 | 148,700 | 8429 | 3.09 | 9 | 67 | 117 | ||
| 14 | 19 | 193,164 | 8672 | 2.94 | 9 | 57 | 107 | |
| 20 | 134,695 | 7270 | 2.48 | 7 | 55 | 107 | ||
| 30 | 27 | 140,484 | 2919 | 1.31 | 7 | 40 | 60 | |
| 28 | 129,491 | 6422 | 2.33 | 6 | 43 | 88 | ||
| 60 | 35 | 295,377 | 13,046 | 4.38 | 10 | 64 | 125 | |
| 36 | 157,505 | 7103 | 2.46 | 6 | 47 | 91 | ||
| 120 | 43 | 294,909 | 12,781 | 4.61 | 10 | 54 | 107 | |
| 44 | 175,689 | 8629 | 3.13 | 10 | 53 | 95 | ||
| C | Milk | MC | 10,632 | 4889 | 1.41 | 10 | 66 | 135 |
| 1 | 5 | 237,563 | 11,167 | 3.76 | 10 | 83 | 165 | |
| 6 | 200,211 | 9565 | 3.24 | 10 | 73 | 156 | ||
| 7 | 13 | 172,573 | 9232 | 3.16 | 9 | 72 | 136 | |
| 14 | 105,377 | 2947 | 1.32 | 5 | 39 | 65 | ||
| 14 | 21 | 281,503 | 11,879 | 4.03 | 10 | 76 | 152 | |
| 22 | 162,633 | 7438 | 2.54 | 8 | 57 | 112 | ||
| 30 | 29 | 374,652 | 14,534 | 5.20 | 8 | 75 | 164 | |
| 30 | 178,935 | 7641 | 2.65 | 9 | 66 | 125 | ||
| 60 | 37 | 390,610 | 14,658 | 5.06 | 12 | 83 | 170 | |
| 38 | 218,602 | 8265 | 2.84 | 8 | 57 | 105 | ||
| 120 | 45 | 116,160 | 5909 | 2.05 | 5 | 47 | 76 | |
| 46 | 188,779 | 8633 | 2.99 | 7 | 51 | 83 | ||
| D | Milk | MD | 52,040 | 11,547 | 3.80 | 21 | 151 | 378 |
| 1 | 7 | 210,147 | 11,128 | 3.71 | 10 | 97 | 217 | |
| 8 | 175,444 | 8825 | 3.03 | 11 | 91 | 197 | ||
| 7 | 15 | 136,796 | 6512 | 2.38 | 7 | 65 | 120 | |
| 16 | 126,129 | 6062 | 2.11 | 8 | 56 | 113 | ||
| 14 | 23 | 194,767 | 8284 | 2.99 | 11 | 66 | 131 | |
| 24 | 201,259 | 9138 | 3.09 | 14 | 73 | 139 | ||
| 30 | 31 | 161,519 | 7050 | 2.39 | 13 | 68 | 119 | |
| 32 | 187,889 | 8164 | 2.78 | 10 | 66 | 136 | ||
| 60 | 39 | 86,466 | 4525 | 1.68 | 8 | 45 | 84 | |
| 40 | 202,656 | 8933 | 3.28 | 7 | 65 | 127 | ||
| 120 | 47 | 205,505 | 9667 | 3.36 | 11 | 67 | 119 | |
| 48 | 209,716 | 10,185 | 3.51 | 15 | 63 | 123 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamarina-García, G.; Hernández, I.; Amores, G.; Virto, M. Characterization of Microbial Shifts during the Production and Ripening of Raw Ewe Milk-Derived Idiazabal Cheese by High-Throughput Sequencing. Biology 2022, 11, 769. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050769
Santamarina-García G, Hernández I, Amores G, Virto M. Characterization of Microbial Shifts during the Production and Ripening of Raw Ewe Milk-Derived Idiazabal Cheese by High-Throughput Sequencing. Biology. 2022; 11(5):769. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050769
Chicago/Turabian StyleSantamarina-García, Gorka, Igor Hernández, Gustavo Amores, and Mailo Virto. 2022. "Characterization of Microbial Shifts during the Production and Ripening of Raw Ewe Milk-Derived Idiazabal Cheese by High-Throughput Sequencing" Biology 11, no. 5: 769. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050769