
Molecular Characterization and Expression Analysis of Putative Class C (Glutamate Family) G Protein-Coupled Receptors in Ascidian Styela clava


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Classification of S. clava GPCRs
2.2. Phylogenetic Analysis, Chromosomal Location, and Structural Prediction
2.3. Gene Expression Analysis of S. clava Class C GPCRs
2.4. Adult Animal, Fertilization, and Swimming Larvae Collection
2.5. Whole-Mount In Situ Hybridization
2.6. Tissue Distribution of S. clava Class C GPCRs
3. Results
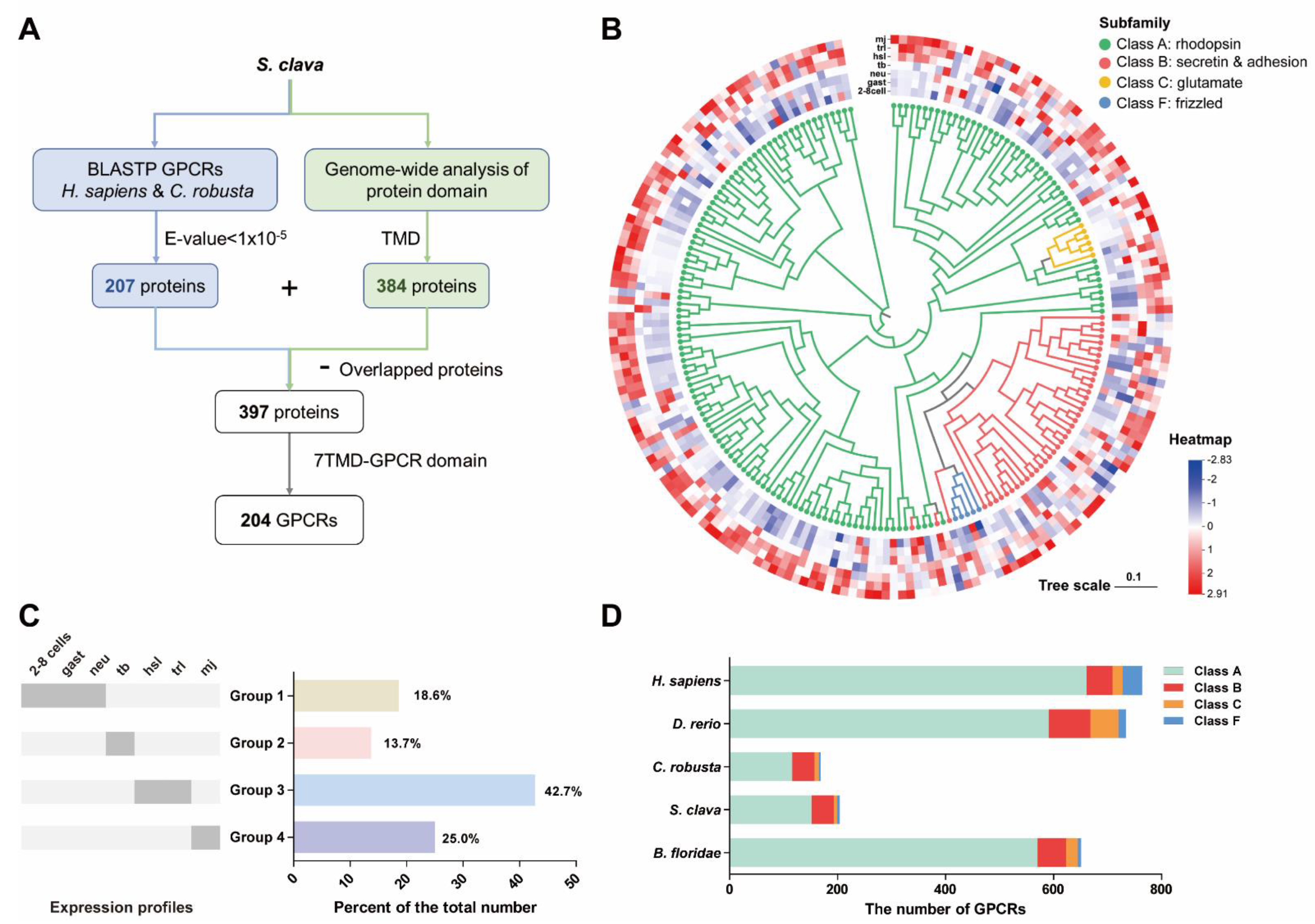
3.1. Prediction of S. clava Putative GPCRs
3.2. Subtypes of Putative S. clava Class C GPCRs
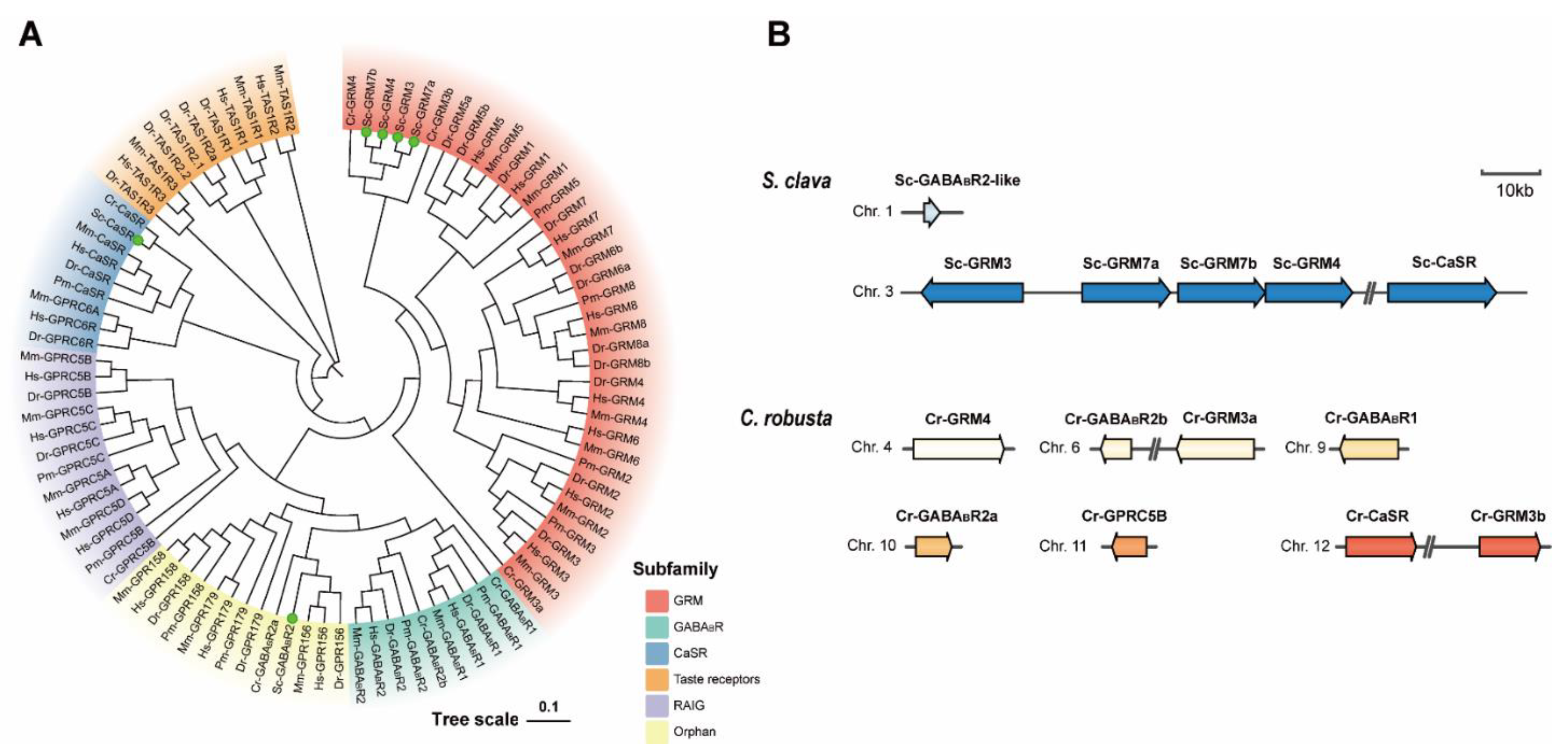
3.3. Phylogenetic and Sequence Analysis of Putative S. clava Class C GPCRs
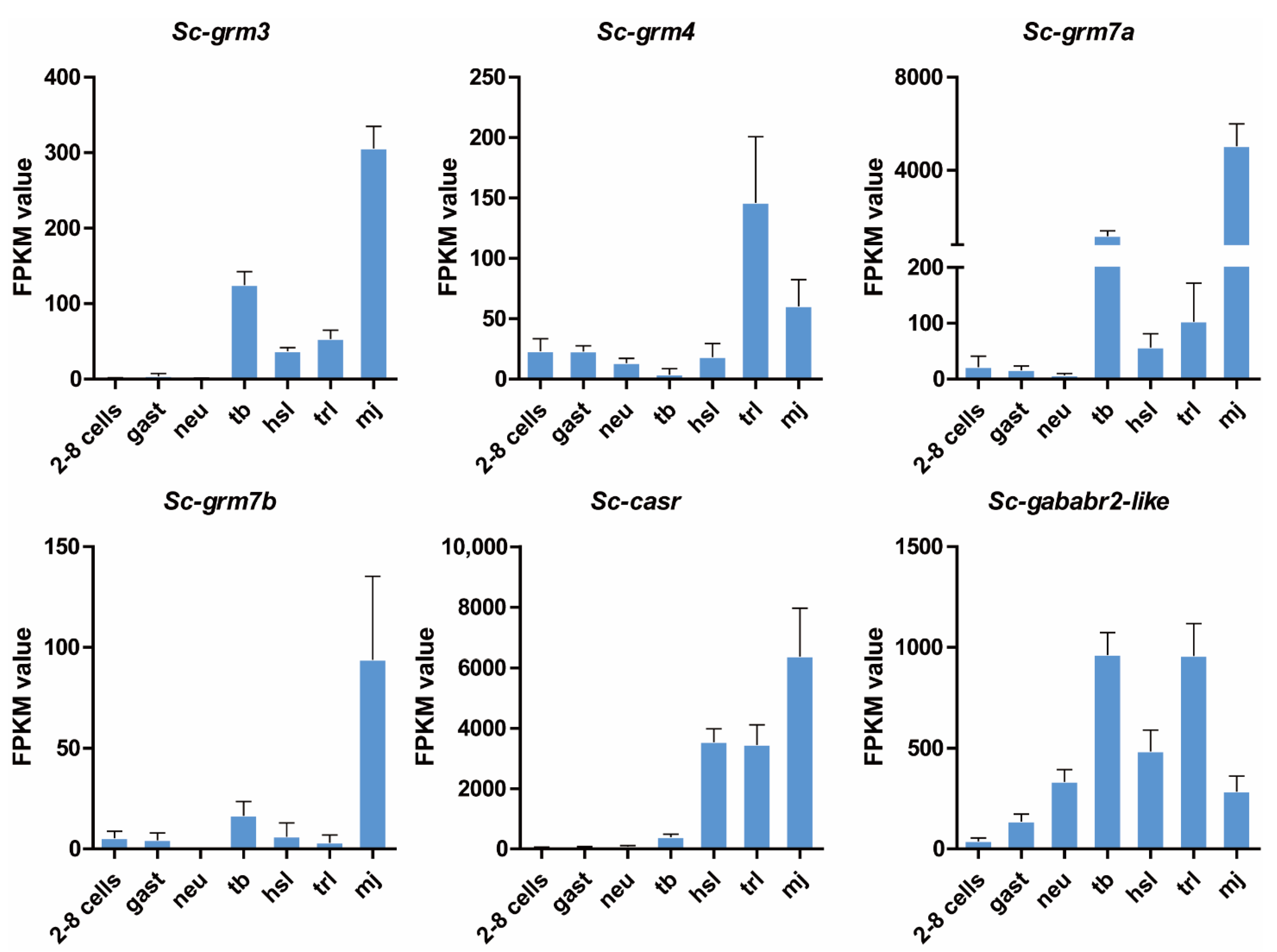
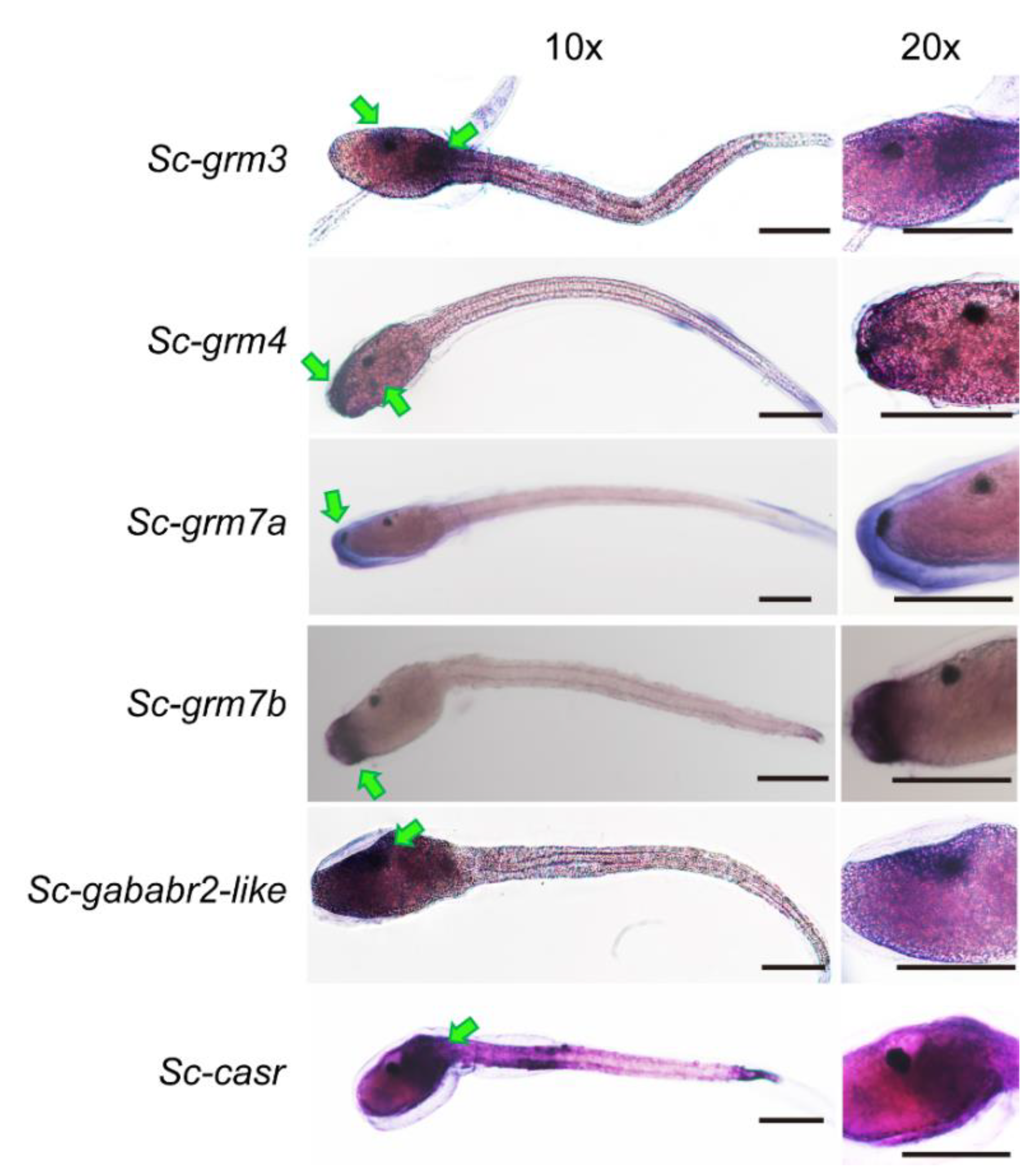
3.4. Expression Pattern of Putative S. clava Class C GPCRs during Development
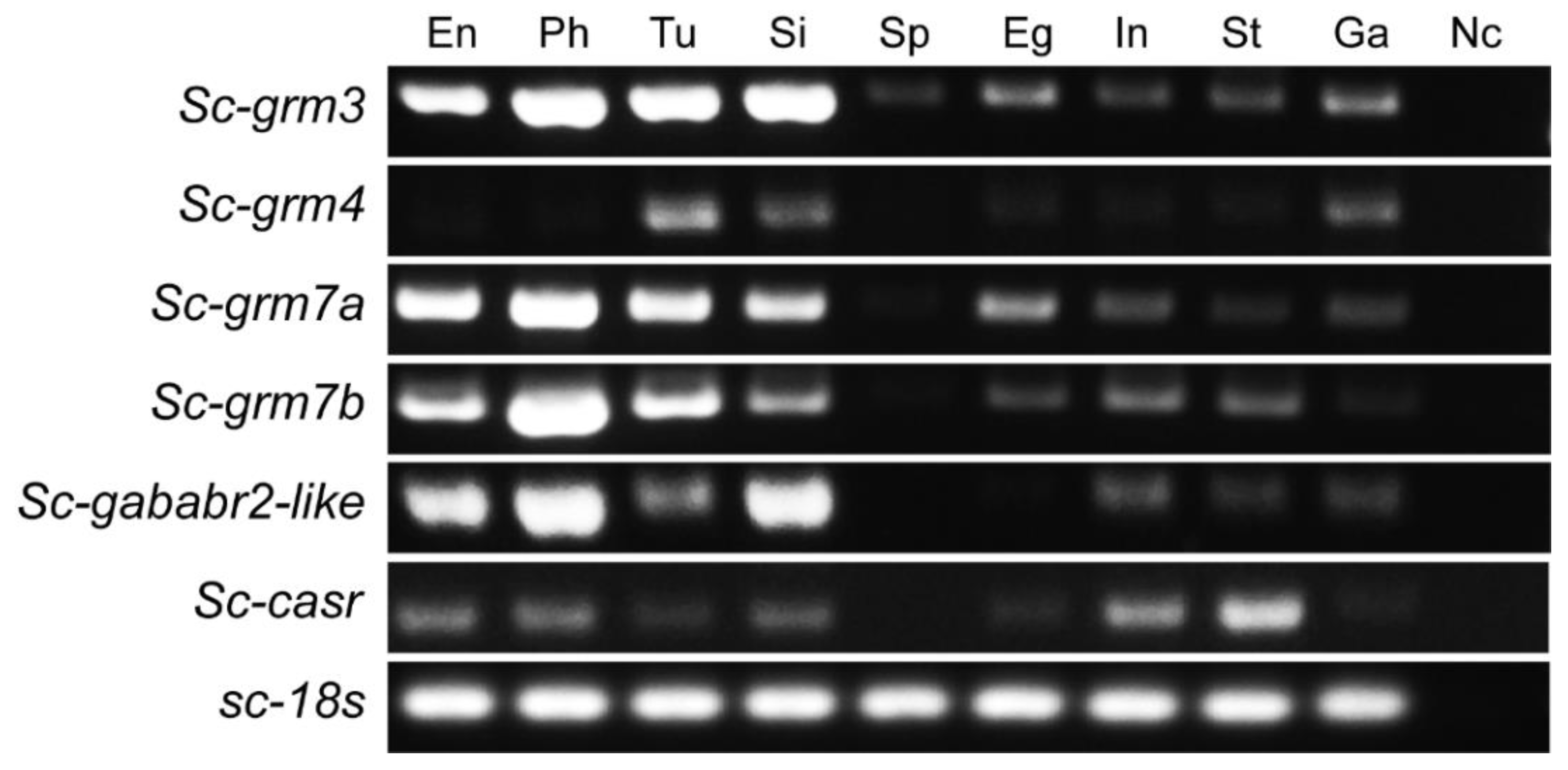
3.5. Expression Pattern of Putative S. clava Class C GPCRs in Swimming Larvae and Different Adult Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Corbo, J.C.; Di Gregorio, A.; Levine, M. The ascidian as a model organism in developmental and evolutionary biology. Cell 2001, 106, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Karaiskou, A.; Swalla, B.J.; Sasakura, Y.; Chambon, J.P. Metamorphosis in solitary ascidians. Genesis 2015, 53, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Aldred, N.; Clare, A.S. BMC evolutionary biologyMini-review: Impact and dynamics of surface fouling by solitary and compound ascidians. Biofouling 2014, 30, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Darbyson, E.A.; Hanson, J.M.; Locke, A.; Willison, J.H.M. Settlement and potential for transport of clubbed tunicate (Styela clava) on boat hulls. Aquat. Invasions 2009, 4, 95–103. [Google Scholar] [CrossRef]
- Olivo, P.; Palladino, A.; Ristoratore, F.; Spagnuolo, A. Brain sensory organs of the ascidian Ciona robusta: Structure, function and developmental mechanisms. Front. Cell Dev. Biol. 2021, 9, 701779. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G.; Tsuda, M. Simple motor system of the ascidian larva: Neuronal complex comprising putative cholinergic and GABAergic/glycinergic neurons. Zool. Sci. 2010, 27, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Yoshida, M.; Morisawa, M. Interaction between noradrenaline or adrenaline and the β1-adrenergic receptor in the nervous system triggers early metamorphosis of larvae in the ascidian, Ciona savignyi. Dev. Biol. 2003, 258, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Zega, G.; Pennati, R.; Groppelli, S.; Sotgia, C.; De Bernardi, F. Dopamine and serotonin modulate the onset of metamorphosis in the ascidian Phallusia mammillata. Dev. Biol. 2005, 282, 246–256. [Google Scholar] [CrossRef] [Green Version]
- Hudson, C. The central nervous system of ascidian larvae. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 538–561. [Google Scholar] [CrossRef] [Green Version]
- Nishino, A. Morphology and Physiology of the Ascidian Nervous Systems and the Effectors. Adv. Exp. Med. Biol. 2018, 1029, 179–196. [Google Scholar]
- Chambon, J.P.; Nakayama, A.; Takamura, K.; McDougall, A.; Satoh, N. ERK-and JNK-signalling regulate gene networks that stimulate metamorphosis and apoptosis in tail tissues of ascidian tadpoles. Development 2007, 134, 1203–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zhang, J.; Lu, Q.; Ren, P.; Guo, X.; Wang, J.; Li, X.; Chang, Y.; Duan, S.; Wang, S.; et al. Genomic basis of environmental adaptation in the leathery sea squirt (Styela clava). Mol. Ecol. Resour. 2020, 20, 1414–1431. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, A.; Matsunobu, S.; Mita, K.; Treen, N.; Sugihara, T.; Horie, T.; Sakuma, T.; Yamamoto, T.; Shiraishi, A.; Hamada, M. GABA-induced GnRH release triggers chordate metamorphosis. Curr. Biol. 2020, 30, 1555–1561.e4. [Google Scholar] [CrossRef] [PubMed]
- Okawa, N.; Shimai, K.; Ohnishi, K.; Ohkura, M.; Nakai, J.; Horie, T.; Kuhara, A.; Kusakabe, T.G. Cellular identity and Ca2+ signaling activity of the non-reproductive GnRH system in the Ciona intestinalis type A (Ciona robusta) larva. Sci. Rep. 2020, 10, 18590. [Google Scholar] [CrossRef] [PubMed]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein-coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.K.; Hou, Z.S.; Tao, Y.X. Biased signaling in naturally occurring mutations of G protein-coupled receptors associated with diverse human diseases. BBA-Mol. Basis Dis. 2021, 1867, 165973. [Google Scholar] [CrossRef]
- Seyedabadi, M.; Ghahremani, M.H.; Albert, P.R. Biased signaling of G protein coupled receptors (GPCRs): Molecular determinants of GPCR/transducer selectivity and therapeutic potential. Pharmacol. Ther. 2019, 200, 148–178. [Google Scholar] [CrossRef]
- Rajagopal, S.; Rajagopal, K.; Lefkowitz, R.J. Teaching old receptors new tricks: Biasing seven-transmembrane receptors. Nat. Rev. Drug Discov. 2010, 9, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, E.; Kumari, P.; Jaiman, D.; Shukla, A.K. Methodological advances: The unsung heroes of the GPCR structural revolution. Nat. Rev. Mol. Cell Biol. 2015, 16, 69–81. [Google Scholar] [CrossRef]
- Møller, T.C.; Moreno-Delgado, D.; Pin, J.P.; Kniazeff, J. Class C G protein-coupled receptors: Reviving old couples with new partners. Biophys. Rep. 2017, 3, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Kunishima, N.; Shimada, Y.; Tsuji, Y.; Sato, T.; Yamamoto, M.; Kumasaka, T.; Nakanishi, S.; Jingami, H.; Morikawa, K. Structural basis of glutamate recognition by a dimeric metabotropic glutamate receptor. Nature 2000, 407, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Margeta-Mitrovic, M.; Jan, Y.N.; Jan, L.Y. Function of GB1 and GB2 subunits in G protein coupling of GABAB receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 14649–14654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruat, M.; Traiffort, E. Roles of the calcium sensing receptor in the central nervous system. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 429–442. [Google Scholar] [CrossRef]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [Green Version]
- Bjarnadóttir, T.K.; Fredriksson, R.; Schiöth, H.B. The gene repertoire and the common evolutionary history of glutamate, pheromone (V2R), taste(1) and other related G protein-coupled receptors. Gene 2005, 362, 70–84. [Google Scholar] [CrossRef]
- Kamesh, N.; Aradhyam, G.K.; Manoj, N. The repertoire of G protein-coupled receptors in the sea squirt Ciona intestinalis. BMC Evol. Biol. 2008, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Zega, G.; Biggiogero, M.; Groppelli, S.; Candiani, S.; Oliveri, D.; Parodi, M.; Pestarino, M.; De Bernardi, F.; Pennati, R. Developmental expression of glutamic acid decarboxylase and of gamma-aminobutyric acid type B receptors in the ascidian Ciona intestinalis. J. Comp. Neurol. 2008, 506, 489–505. [Google Scholar] [CrossRef]
- Brown, E.R.; Nishino, A.; Bone, Q.; Meinertzhagen, I.A.; Okamura, Y. GABAergic synaptic transmission modulates swimming in the ascidian larva. Eur. J. Neurosci. 2005, 22, 2541–2548. [Google Scholar] [CrossRef]
- Goldstien, S.J.; Schiel, D.R.; Gemmell, N.J. Regional connectivity and coastal expansion: Differentiating pre-border and post-border vectors for the invasive tunicate Styela clava. Mol. Ecol. 2010, 19, 874–885. [Google Scholar] [CrossRef]
- Goldstien, S.J.; Dupont, L.; Viard, F.; Hallas, P.J.; Nishikawa, T.; Schiel, D.R.; Gemmell, N.J.; Bishop, J.D. Global phylogeography of the widely introduced North West Pacific ascidian Styela clava. PLoS ONE 2011, 6, e16755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooistra, A.J.; Mordalski, S.; Pándy-Szekeres, G.; Esguerra, M.; Mamyrbekov, A.; Munk, C.; Keserű, G.M.; Gloriam, D.E. GPCRdb in 2021: Integrating GPCR sequence, structure and function. Nucleic Acids Res. 2021, 49, D335–D343. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; He, Q.; Studholme, D.J.; Wu, Q.; Liang, S.; Yu, L. NCD3G: A novel nine-cysteine domain in family 3 GPCRs. Trends Biochem. Sci. 2004, 29, 458–461. [Google Scholar] [CrossRef]
- Muto, T.; Tsuchiya, D.; Morikawa, K.; Jingami, H. Structures of the extracellular regions of the group II/III metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2007, 104, 3759–3764. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiaen, L.; Wagner, E.; Shi, W.; Levine, M. Whole-mount in situ hybridization on sea squirt (Ciona intestinalis) embryos. Cold Spring Harb. Protoc. 2009, 2009, pdb.prot5348. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, R.; Schiöth, H.B. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol. Pharmacol. 2005, 67, 1414–1425. [Google Scholar] [CrossRef] [PubMed]
- Nordström, K.J.; Fredriksson, R.; Schiöth, H.B. The amphioxus (Branchiostoma floridae) genome contains a highly diversified set of G protein-coupled receptors. BMC Evol. Biol. 2008, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.N.; Secker, A.; Halling-Brown, M.; Moss, D.S.; Freitas, A.A.; Timmis, J.; Clark, E.; Flower, D.R. GPCRTree: Online hierarchical classification of GPCR function. BMC Res. Notes 2008, 1, 67. [Google Scholar] [CrossRef] [Green Version]
- Herberger, A.L.; Loretz, C.A. Vertebrate extracellular calcium-sensing receptor evolution: Selection in relation to life history and habitat. Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 86–94. [Google Scholar] [CrossRef]
- Brody, T.; Cravchik, A. Drosophila melanogaster G protein–coupled receptors. J. Cell Biol. 2000, 150, F83–F88. [Google Scholar] [CrossRef]
- Pin, J.P.; Galvez, T.; Prézeau, L. Evolution, structure, and activation mechanism of family 3/C G-protein-coupled receptors. Pharmacol. Ther. 2003, 98, 325–354. [Google Scholar] [CrossRef]
- Kang, X.L.; Li, Y.X.; Li, Y.L.; Wang, J.X.; Zhao, X.F. The homotetramerization of a GPCR transmits the 20-hydroxyecdysone signal and increases its entry into cells for insect metamorphosis. Development 2021, 148, dev196667. [Google Scholar] [CrossRef]
- Bai, H.; Zhu, F.; Shah, K.; Palli, S.R. Large-scale RNAi screen of G protein-coupled receptors involved in larval growth, molting and metamorphosis in the red flour beetle. BMC Genom. 2011, 12, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.H.; Garcia, C.R. Evidence of G-Protein-Coupled Receptors (GPCR) in the Parasitic Protozoa Plasmodium falciparum—Sensing the Host Environment and Coupling within Its Molecular Signaling Toolkit. Int. J. Mol. Sci. 2021, 22, 12381. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, A.; Okuda, T.; Miyasaka, N.; Osugi, T.; Okuno, Y.; Inoue, J.; Satake, H. Repertoires of G protein-coupled receptors for Ciona-specific neuropeptides. Proc. Natl. Acad. Sci. USA 2019, 116, 7847–7856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satake, H.; Ogasawara, M.; Kawada, T.; Masuda, K.; Aoyama, M.; Minakata, H.; Chiba, T.; Metoki, H.; Satou, Y.; Satoh, N. Tachykinin and tachykinin receptor of an ascidian, Ciona intestinalis: Evolutionary origin of the vertebrate tachykinin family. J. Biochem. 2004, 279, 53798–53805. [Google Scholar]
- Sakai, T.; Yamamoto, T.; Matsubara, S.; Kawada, T.; Satake, H. Invertebrate gonadotropin-releasing hormone receptor signaling and its relevant biological actions. Int. J. Mol. Sci. 2020, 21, 8544. [Google Scholar] [CrossRef]
- Dehal, P.; Satou, Y.; Campbell, R.K.; Chapman, J.; Degnan, B.; De Tomaso, A.; Davidson, B.; Di Gregorio, A.; Gelpke, M.; Goodstein, D.M.; et al. The draft genome of Ciona intestinalis: Insights into chordate and vertebrate origins. Science 2002, 298, 2157–2167. [Google Scholar] [CrossRef] [Green Version]
- Satou, Y.; Nakamura, R.; Yu, D.; Yoshida, R.; Hamada, M.; Fujie, M.; Hisata, K.; Takeda, H.; Satoh, N. A Nearly Complete Genome of Ciona intestinalis Type A (C. robusta) Reveals the Contribution of Inversion to Chromosomal Evolution in the Genus Ciona. Genome Biol. Evol. 2019, 11, 3144–3157. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.L.; Li, Y.X.; Wang, X.P.; Kang, X.L.; Guo, K.Q.; Dong, D.J.; Wang, J.X.; Zhao, X.F. Identification and Functional Analysis of G Protein-Coupled Receptors in 20-Hydroxyecdysone Signaling From the Helicoverpa armigera Genome. Front. Cell Dev. Biol. 2021, 9, 753787. [Google Scholar] [CrossRef]
- Mitri, C.; Parmentier, M.L.; Pin, J.P.; Bockaert, J.; Grau, Y. Divergent evolution in metabotropic glutamate receptors: A new receptor activated by an endogenous ligand different from glutamate in insects. J. Biochem. 2004, 279, 9313–9320. [Google Scholar]
- Mitri, C.; Soustelle, L.; Framery, B.; Bockaert, J.; Parmentier, M.L.; Grau, Y. Plant insecticide L-canavanine repels Drosophila via the insect orphan GPCR DmX. PLoS Biol. 2009, 7, e1000147. [Google Scholar] [CrossRef] [Green Version]
- Conn, P.J.; Pin, J.P. Pharmacology and functions of metabotropic glutamate receptors. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 205–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutzeit, V.A.; Thibado, J.; Stor, D.S.; Zhou, Z.; Blanchard, S.C.; Andersen, O.S.; Levitz, J. Conformational dynamics between transmembrane domains and allosteric modulation of a metabotropic glutamate receptor. eLife 2019, 8, e45116. [Google Scholar] [CrossRef] [PubMed]
- Niimura, Y. Evolutionary dynamics of olfactory receptor genes in chordates: Interaction between environments and genomic contents. Hum. Genom. 2009, 4, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddall, M.E. Invertebrates.—R.C. Brusca and G.J. Brusca. 2003. Sinauer Associates, Sunderland, Massachusetts. Syst. Biol. 2004, 53, 664–666. [Google Scholar]
- Jones, K.A.; Tamm, J.A.; Craig, D.A.; Ph, D.; Yao, W.; Panico, R. Signal transduction by GABA(B) receptor heterodimers. Neuropsychopharmacology 2000, 23 (Suppl. S4), S41–S49. [Google Scholar] [CrossRef]
- Satoh, N. Developmental Genomics of Ascidians; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Horie, T.; Nakagawa, M.; Sasakura, Y.; Kusakabe, T.G. Cell type and function of neurons in the ascidian nervous system. Dev. Growth Differ. 2009, 51, 207–220. [Google Scholar] [CrossRef]
- Horie, T.; Kusakabe, T.; Tsuda, M. Glutamatergic networks in the Ciona intestinalis larva. J. Comp. Neurol. 2008, 508, 249–263. [Google Scholar] [CrossRef]
- Sasakura, Y.; Hozumi, A. Formation of adult organs through metamorphosis in ascidians. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e304. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Haug, M.F.; Gesemann, M.; Neuhauss, S.C.F. Novel expression patterns of metabotropic glutamate receptor 6 in the zebrafish nervous system. PLoS ONE 2012, 7, e35256. [Google Scholar] [CrossRef]
- Andrianov, G.N.; Puyal, J.; Raymond, J.; Venteo, S.; Dememes, D.; Ryzhova, I.V. Immunocytochemical and pharmacological characterization of metabotropic glutamate receptors of the vestibular end organs in the frog. Hear. Res. 2005, 204, 200–209. [Google Scholar] [CrossRef]
- Haug, M.F.; Gesemann, M.; Mueller, T.; Neuhauss, S.C.F. Phylogeny and expression divergence of metabotropic glutamate receptor genes in the brain of zebrafish (Danio rerio). J. Comp. Neurol. 2013, 521, 1533–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.H.; Yang, N.; Ma, Y.H.; Jiang, J.; Zhang, J.F.; Fei, J.; Guo, L.H. Identification of glutamate receptors and transporters in mouse and human sperm. J. Androl. 2004, 25, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Storto, M.; Sallese, M.; Salvatore, L.; Poulet, R.; Condorelli, D.F.; Dell Albani, P.; Marcello, M.F.; Romeo, R.; Piomboni, P.; Barone, N. Expression of metabotropic glutamate receptors in the rat and human testis. J. Endocrinol. 2001, 170, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, D.; Kon, S.; Sato, T.; Toyama, F.; Katsura, Y.; Nakauchi, Y.; Takayama-Watanabe, E.; Watanabe, A. NMDA-type glutamate receptors mediate the acrosome reaction and motility initiation in newt sperm. Mol. Reprod. Dev. 2019, 86, 1106–1115. [Google Scholar] [CrossRef]
- Osugi, T.; Sasakura, Y.; Satake, H. The nervous system of the adult ascidian Ciona intestinalis Type A (Ciona robusta): Insights from transgenic animal models. PLoS ONE 2017, 12, e0180227. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Cheng, C.Y.; Sun, X.; Pedicone, A.J.; Mohamadzadeh, M.; Cheng, S.X. The extracellular calcium-sensing receptor in the intestine: Evidence for regulation of colonic absorption, secretion, motility, and immunity. Front. Physiol. 2016, 7, 245. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Purpose |
|---|---|---|---|
| Sc-grm3 | AACAGGAACATGTCAAGCGAT | GAACTACAACAACCAAGAGCG | Probe synthesis RT-PCR |
| Sc-grm4 | AGACTCTTGGACGAGATTGCT | GGTCTCATGCCTTGGTTACAT | Probe synthesis RT-PCR |
| Sc-grm7a | AGTACAGAGAAACGGCGATAGT | TTGTGAACCCCACGTAGAGAG | Probe synthesis RT-PCR |
| Sc-grm7b | GCCAACAGATAATAACGACGA | GACAGCATGAACAAATGAGAGA | Probe synthesis RT-PCR |
| Sc-gababr2-like | TGGCAACAAAGACATGGAGGG | GCTGTGACGACGAATTAGAATCA | Probe synthesis RT-PCR |
| Sc-casr | TTTCTTCATCGTTTGGTTGTCC | CATTTTGTCCTCGCTTTTTGG | Probe synthesis RT-PCR |
| Sc-18s | CTGAGTGAAGCAGCGAGTGTCTAACCTA | CTGAGTGAAGCAGCGAGTGTCTAACCTA | RT-PCR |
| Receptor | Gene ID | Predicted Domains | Homology Search (BLASTP) | ||
|---|---|---|---|---|---|
| Species | Protein ID | E-Value | |||
| Sc-GRM3 | evm.model.000002F_arrow_pilon.86 | VFD + CRD + 7TMD | H. sapiens | NP_000835.1 | 1 × 10−107 |
| C. robusta | XP_018671926.1 | 5 × 10−130 | |||
| Sc-GRM4 | evm.model.000002F_arrow_pilon.88.2 | VFD + CRD + 7TMD | H. sapiens | NP_000832.1 | 6 × 10−132 |
| C. robusta | XP_018671926.1 | 2 × 10−161 | |||
| Sc-GRM7a | evm.model.000002F_arrow_pilon.87 | VFD + CRD + 7TMD | H. sapiens | NP_000835.1 | 2 × 10−157 |
| C. robusta | XP_018671926.1 | 9 × 10−172 | |||
| Sc-GRM7b | evm.model.000002F_arrow_pilon.88.1 | VFD + CRD + 7TMD | H. sapiens | NP_000835.1 | 1 × 10−108 |
| C. robusta | XP_018671926.1 | 3 × 10−112 | |||
| Sc-CaSR | evm.model.000075F_arrow_pilon.22 | VFD + CRD + 7TMD | H. sapiens | NP_000379.3 | 0 |
| C. robusta | XP_026692769.1 | 0 | |||
| Sc-GABABR2-like | evm.model.000022F_arrow_pilon.50 | 7TMD | H. sapiens | XP_005252373.1 | 1 × 10−35 |
| C. robusta | XP_009861983.2 | 5 × 10−33 | |||
| Receptor | H. sapiens | D. rerio | C. robusta | S. clava | B. floridae | D. melanogaster | C. elegans |
|---|---|---|---|---|---|---|---|
| GRM | 8 | 13 | 3 | 4 | 12 | 2 | 3 |
| GABABR | 2 | 3 | 2 | 0 | 4 | 3 | 2 |
| CaSR | 1 | 1 | 1 | 1 | 1 | 0 | 0 |
| TAS1R | 3 | 4 | 0 | 0 | 0 | 0 | 0 |
| VR | 0 | 24 | 0 | 0 | 0 | 0 | 0 |
| Others | 8 | 7 | 2 | 1 | 5 | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Dong, B.; Yang, L. Molecular Characterization and Expression Analysis of Putative Class C (Glutamate Family) G Protein-Coupled Receptors in Ascidian Styela clava. Biology 2022, 11, 782. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050782
Zhang J, Dong B, Yang L. Molecular Characterization and Expression Analysis of Putative Class C (Glutamate Family) G Protein-Coupled Receptors in Ascidian Styela clava. Biology. 2022; 11(5):782. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050782
Chicago/Turabian StyleZhang, Jin, Bo Dong, and Likun Yang. 2022. "Molecular Characterization and Expression Analysis of Putative Class C (Glutamate Family) G Protein-Coupled Receptors in Ascidian Styela clava" Biology 11, no. 5: 782. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050782