
Seven Sycoryctine Fig Wasp Species (Chalcidoidea: Pteromalidae) Associated with Dioecious Ficus hirta Inhabiting South China and Southeast Asia


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
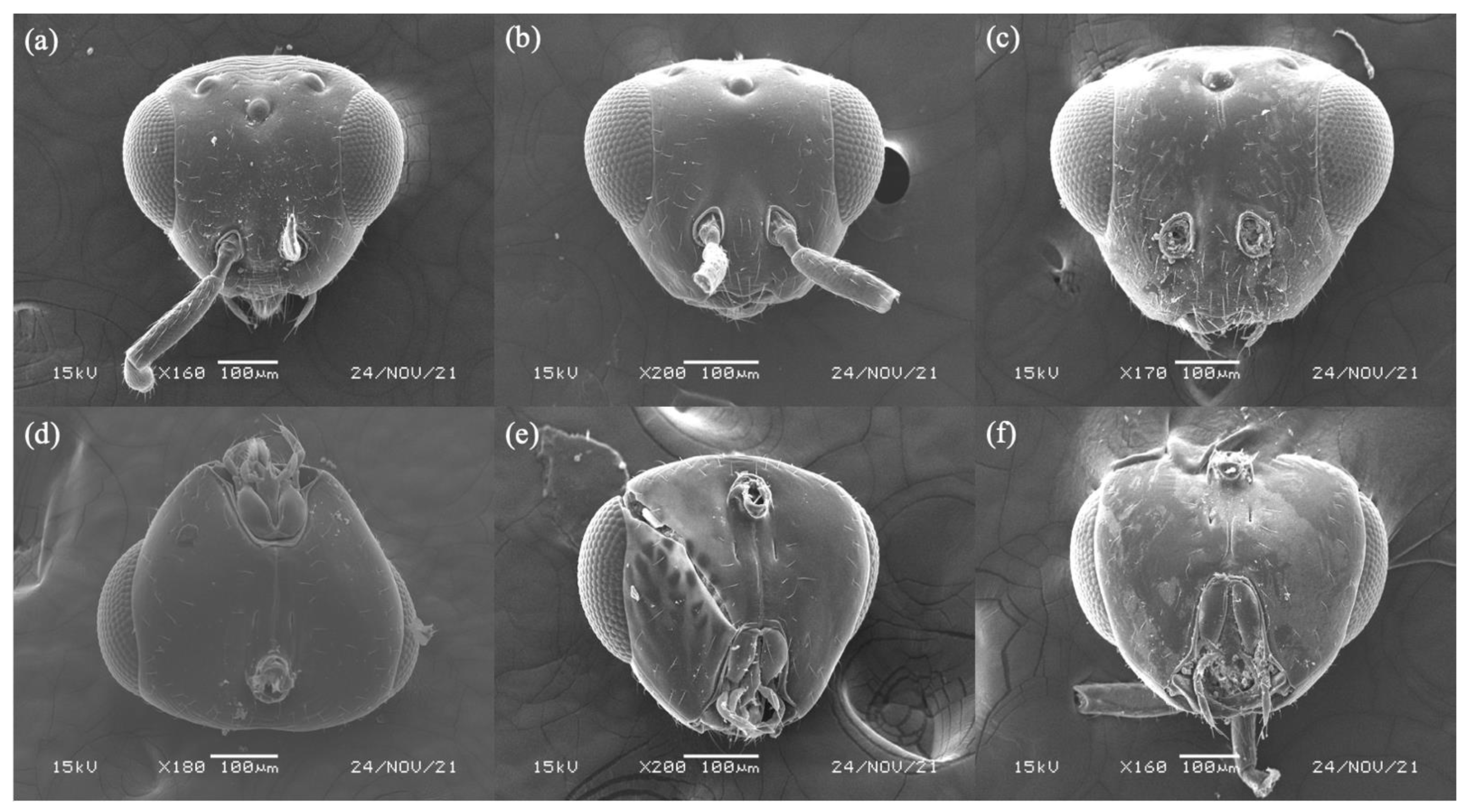
3.1. Philotrypesis Forster, 1878
3.1.1. Philotrypesis Forster, 1878: 153–187. Type Species: Philotrypesis Longicaudata Mayr, 1906
Philotrypesis guangdongensis Yu sp. n.
Philotrypesis yunnanensis Yu sp. n.
Philotrypesis fujianensis Yu sp. n.
3.2. Sycoryctes Mayr, 1885
3.2.1. Sycoryctes Mayr, 1885: 153–187. Type Species: Sycoryctes Patellaris Mayr, 1885
Sycoryctes javaensis Yu sp. n.
3.3. Sycoscapter Saunders, 1883
3.3.1. Sycoscapter Saunders, 1883: 29–47. Type Species: Sycoscapter Insignis Saunders, 1883
Sycoscapter chinensis Yu sp. n.
Sycoscapter thaiensis Yu sp. n.
Sycoscapter singaporensis Yu sp. n.
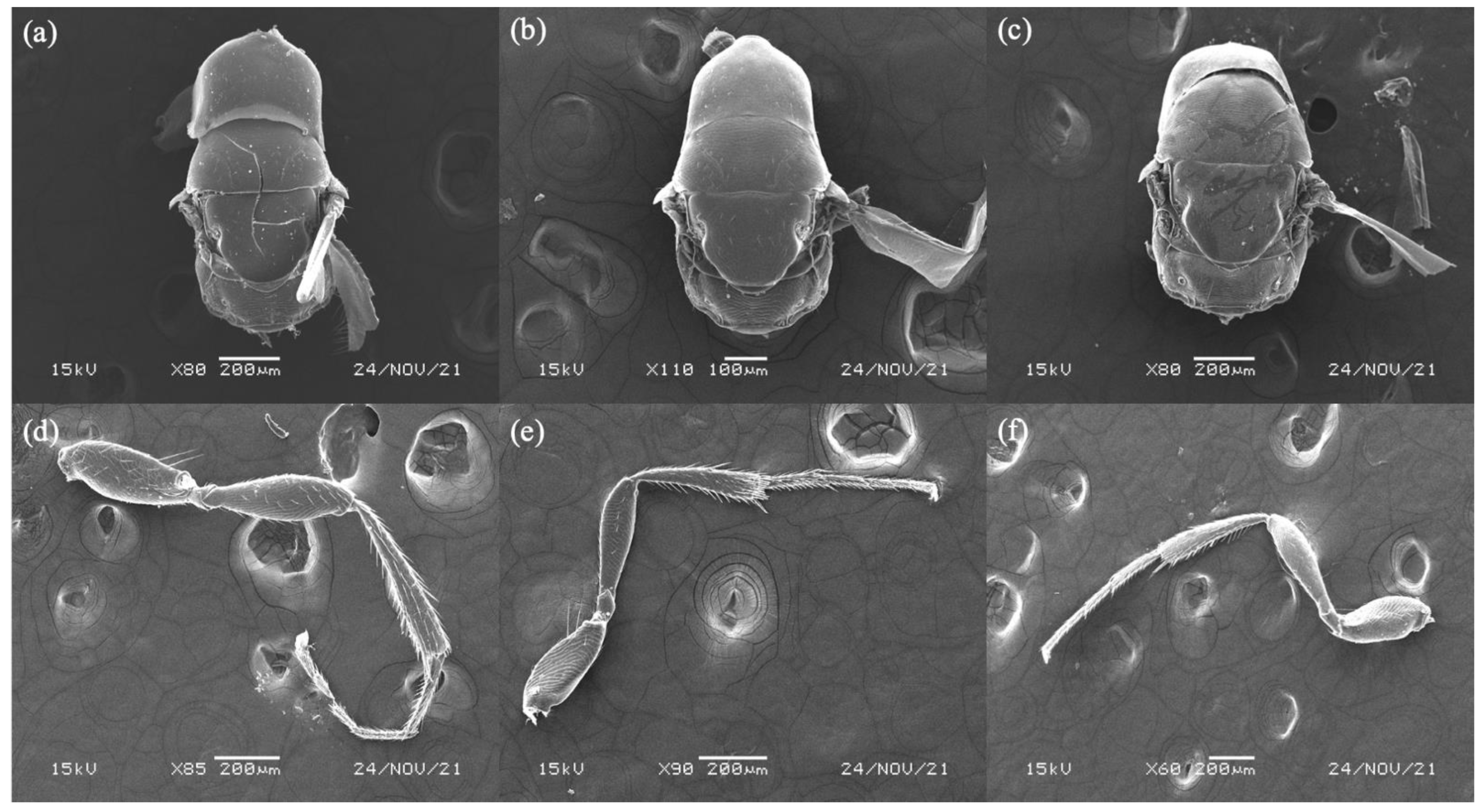
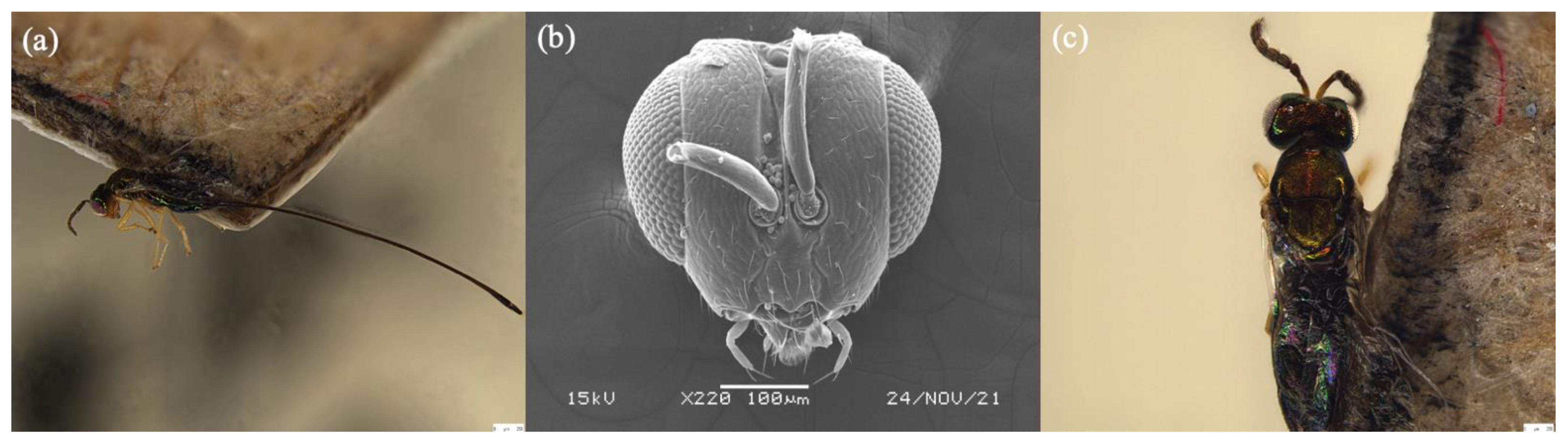
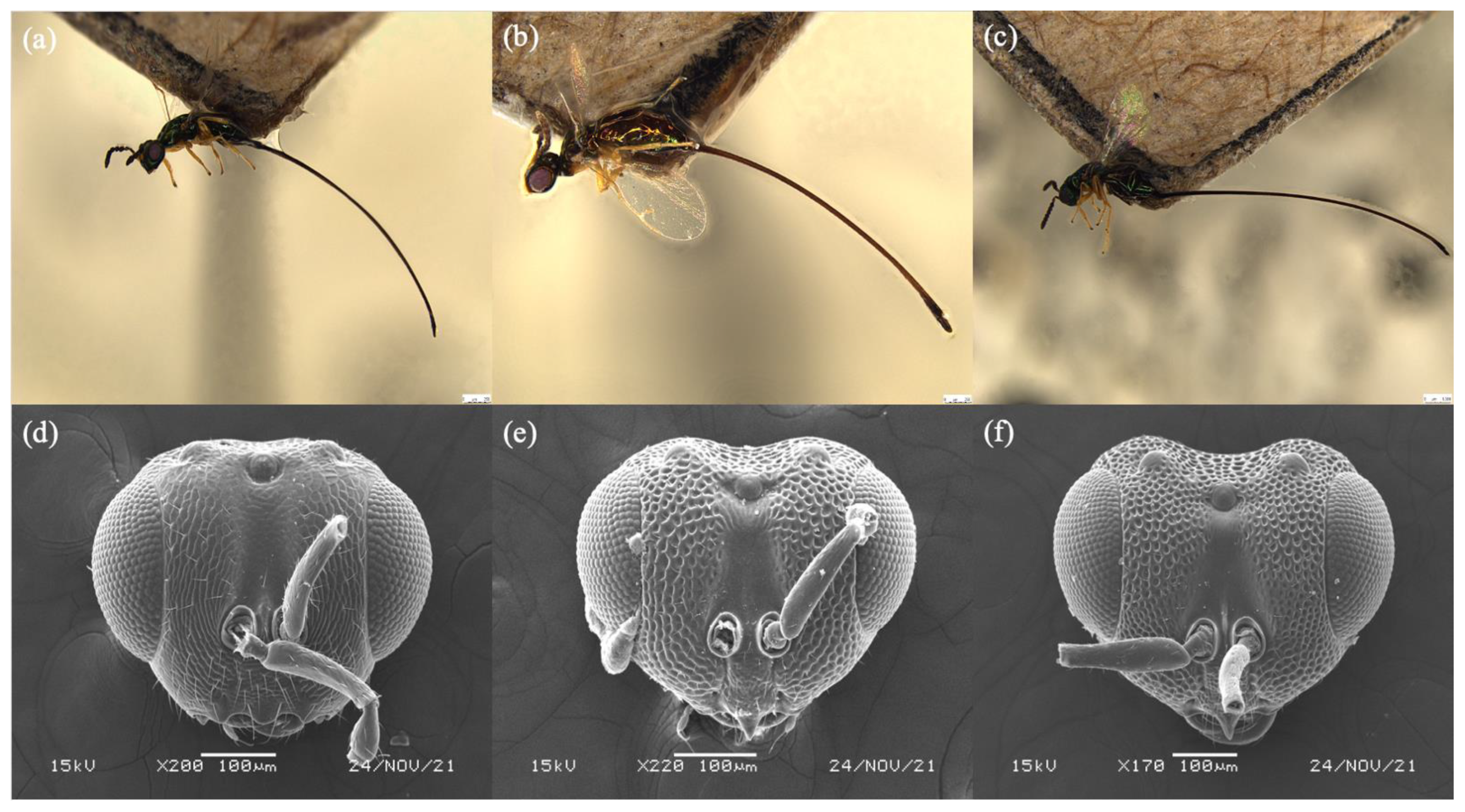
3.4. Diagnoses of Female Sycoryctine Species Associated with Ficus Hirta
3.5. Key to Female Sycoryctine Species Associated with Ficus Hirta
- 1a.
- Toruli apart; gastral tail consists of two last tergites, ovipositor and its sheaths; stigmal vein without knob; body non-metallic gloss (Genus Philotrypesis Forster) …………. 2
- 1b.
- Toruli approach and located above the bottom line of compound eyes; gastral tail consists of a last tergites, ovipositor and its sheaths; stigmal vein with a knob; body with metallic gloss ………………………………………………………………………………………………4
- 2a.
- Black band on scutellum indistinct (Figure 1b) …………………. P. yunnanensis sp. n.
- 2b.
- Black band on scutellum distinct ……………………………………………………… 3
- 3a.
- 3b.
- Mouthpart extended below the central of compound eyes (Figure 2f); black band on mesoscutum does not bifurcate ………………………………………… P. fujianensis sp. n.
- 4a.
- Epistomal margin without acute projection (Figure 4b); knob on stigmal vein does not elongate; wing pilosity strongly reduced ……………………. Sycoryctes javaensis sp. n.
- 4b.
- Epistomal margin with an acute projection; fore wing with some long robust hairs below the marginal vein (Genus Sycoscapter Saunders) ……………………………………………………………………………………………… 5
- 5a.
- Face without deep sculpture (Figure 5d) ……………………………… S. chinensis sp. n.
- 5b.
- Face with deep sculpture ………………………………………………………………. 6
- 6a.
- Metallic green body color with brownish reflection (Figure 6b) …… S. thaiensis sp. n.
- 6b.
- Metallic green body color with blue reflection (Figure 6c) ……. S. singaporensis sp. n.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cook, J.M.; Rasplus, J.-Y. Mutualists with attitude: Coevolving fig wasps and figs. Trends Ecol. Evol. 2003, 18, 241–248. [Google Scholar] [CrossRef]
- Jousselin, E.; Van Noort, S.; Greeff, J.M. Labile male morphology and intraspecific male polymorphism in the Philotrypesis fig wasps. Mol. Phylogenetics Evol. 2004, 33, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Compton, S.G. African fig wasp communities: Undersaturation and latitudinal gradients in species richness. J. Anim. Ecol. 1992, 61, 361–372. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/5328 (accessed on 9 July 2021). [CrossRef]
- Van Noort, S.; Rasplus, J. Revision of the otiteselline fig wasps (Hymenoptera: Chalcidoidea: Agaonidae), I: The Otitesella digitata species-group of the Afrotropical region, with a key to Afrotropical species of Otitesella Westwood. Afr. Entomol. 1997, 5, 125–147. Available online: https://hdl.handle.net/10520/AJA10213589_161 (accessed on 9 July 2021).
- Segar, S.T.; Lopez-Vaamonde, C.; Rasplus, J.-Y.; Cook, J.M. The global phylogeny of the subfamily Sycoryctinae (Pteromalidae): Parasites of an obligate mutualism. Mol. Phylogenetics Evol. 2012, 65, 116–125. [Google Scholar] [CrossRef]
- Weiblen, G.D. How to be a fig wasp. Annu. Rev. Entomol. 2002, 47, 299–330. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.F.; Huang, D.W.; Chen, L.L.; Zhen, W.Q.; Fu, Y.G.; Peng, Z.Q. Rampant host switching and multiple female body colour transitions in Philotrypesis (Hymenoptera: Chalcidoidea: Agaonidae). J. Evol. Biol. 2006, 19, 1157–1166. [Google Scholar] [CrossRef]
- Wong, D.-M.; Bain, A.; Chou, L.-S.; Shiao, S.-F. Description of two new species of fig wasps (Chalcidoidea: Pteromalidae: Sycoryctinae) associated with Ficus benguetensis. Taiwania 2018, 63, 155–162. [Google Scholar] [CrossRef]
- Pramanik, A.; Dey, D. Two new fig wasp species of genus Sycoscapter Saunders, 1883 (Hymenoptera: Chalcidoidea: Pteromalidae) with a key to species of the genus from India. Taiwania 2019, 64, 139–148. [Google Scholar] [CrossRef]
- McLeish, M.J.; Beukman, G.; van Noort, S.; Wossler, T.C. Host-plant species conservatism and ecology of a parasitoid fig wasp genus (Chalcidoidea; Sycoryctinae; Arachonia). PLoS ONE 2012, 7, e44804. [Google Scholar] [CrossRef]
- Tzeng, H.-Y.; Tseng, L.-J.; Ou, C.-H.; Lu, K.-C.; Lu, F.-Y.; Chou, L.-S. Confirmation of the parasitoid feeding habit in Sycoscapter, and their impact on pollinator abundance in Ficus formosana. Symbiosis 2008, 45, 129–134. [Google Scholar]
- Kuttamathiathu, J. The biology of Philotrypesis caricae (L.), parasite of Blastophaga psenes (L.) (Chalcidoidea: Parasitic Hymenoptera). Proc. Int. Congr. Zool 1959, 15, 662–664. [Google Scholar]
- Segar, S.T.; Cook, J.M. The dominant exploiters of the fig/pollinator mutualism vary across continents, but their costs fall consistently on the male reproductive function of figs. Ecol. Entomol. 2012, 37, 342–349. [Google Scholar] [CrossRef]
- Deng, X.; Chen, L.; Tian, E.; Zhang, D.; Wattana, T.; Yu, H.; Kjellberg, F.; Segar, S.T. Low host specificity and broad geographical ranges in a community of parasitic non-pollinating fig wasps (Sycoryctinae; Chalcidoidea). J. Anim. Ecol. 2021, 90, 1678–1690. [Google Scholar] [CrossRef]
- Noyes, J.S. Collecting and preserving chalcid wasps (Hymenoptera: Chalcidoidea). J. Nat. Hist. 1982, 16, 315–334. [Google Scholar] [CrossRef]
- Kawada, R.; Buffington, M.L. A scalable and modular dome illumination system for scientific microphotography on a budget. PLoS ONE 2016, 11, e0153426. [Google Scholar] [CrossRef]
- Gibson, G.; Huber, J.; Woolley, J. Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997. [Google Scholar]
- Yoder, M.J.; Mikó, I.; Seltmann, K.C.; Bertone, M.A.; Deans, A.R. A gross anatomy ontology for Hymenoptera. PLoS ONE 2010, 5, e15991. [Google Scholar] [CrossRef]
- Berg, C.; Corner, E. Moraceae (Ficus); Flora Malesiana, Series I (Seed plants); Ational Herbarium of The Netherlands: Leiden, The Netherlands, 2005; Volume 17. [Google Scholar]
- Yu, H.; Zhao, N.-X.; Chen, Y.-Z.; Deng, Y.; Yao, J.-Y.; Ye, H.-G. Phenology and reproductive strategy of a common fig in Guangzhou. Bot. Stud. 2006, 47, 435–441. [Google Scholar]
- Chen, Y.; Compton, S.G.; Liu, M.; Chen, X.Y. Fig trees at the northern limit of their range: The distributions of cryptic pollinators indicate multiple glacial refugia. Mol. Ecol. 2012, 21, 1687–1701. [Google Scholar] [CrossRef]
- Bain, A.; Borges, R.M.; Chevallier, M.-H.; Vignes, H.; Kobmoo, N.; Peng, Y.; Cruaud, A.; Rasplus, J.-Y.; Kjellberg, F.; Hossaert-Mckey, M. Geographic structuring into vicariant species-pairs in a wide-ranging, high-dispersal plant–insect mutualism: The case of Ficus racemosa and its pollinating wasps. Evol. Ecol. 2016, 30, 663–684. [Google Scholar] [CrossRef]
- Rodriguez, L.J.; Bain, A.; Chou, L.-S.; Conchou, L.; Cruaud, A.; Gonzales, R.; Hossaert-McKey, M.; Rasplus, J.-Y.; Tzeng, H.-Y.; Kjellberg, F. Diversification and spatial structuring in the mutualism between Ficus septica and its pollinating wasps in insular South East Asia. BMC Evol. Biol. 2017, 17, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Tian, E.; Zheng, L.; Deng, X.; Cheng, Y.; Chen, L.; Wu, W.; Tanming, W.; Zhang, D.; Compton, S.G. Multiple parapatric pollinators have radiated across a continental fig tree displaying clinal genetic variation. Mol. Ecol. 2019, 28, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liang, D.; Tian, E.; Zheng, L.; Kjellberg, F. Plant geographic phenotypic variation drives diversification in its associated community of a phytophagous insect and its parasitoids. BMC Evol. Biol. 2018, 18, 134. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.; Cruaud, A.; Rasplus, J.-Y. Low sampling effort and high genetic isolation contribute to under-documented diversity in Philippine fig wasps. Philipp. J. Sci. 2020, 150, 173–180. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wasp Species | Country | Site | Latitude, Longitude |
|---|---|---|---|
| Philotrypesis guangdongensis | China | Gui | 25.077, 110.306 |
| Huo | 23.170, 113.373 | ||
| DHS | 23.166, 112.543 | ||
| Xiang | 22.424, 114.306 | ||
| Nan | 22.787, 108.389 | ||
| Ding | 19.697, 110.328 | ||
| Wan | 18.795, 110.391 | ||
| Thailand | QMS | 18.809, 98.914 | |
| CH | 12.774, 102.096 | ||
| Wu | 14.443, 105.273 | ||
| HB | 12.999, 108.230 | ||
| ST | 7.467, 99.639 | ||
| P. yunnanensis | China | Sand | 25.984, 107.874 |
| XI | 21.913, 101.264 | ||
| Thailand | QMS | 18.809, 98.914 | |
| Tai | 18.894, 98.858 | ||
| CS | 18.84, 99.47 | ||
| P. fujianensis | China | Ning | 26.664, 119.549 |
| Sha | 26.419, 117.818 | ||
| Xia | 24.742, 118.072 | ||
| Sui | 26.476, 114.239 | ||
| Da | 24.258, 116.806 | ||
| Sycoryctes javaensis | Indonesia | CI | −6.566, 106.706 |
| JA | −6.368, 106.830 | ||
| Sycoscapter chinensis | China | Gui | 25.077, 110.306 |
| Huo | 23.170, 113.373 | ||
| DHS | 23.166, 112.543 | ||
| Xiang | 22.424, 114.306 | ||
| Sand | 25.984, 107.874 | ||
| Nan | 22.787, 108.389 | ||
| Ding | 19.697, 110.328 | ||
| Wan | 18.795, 110.391 | ||
| XI | 21.913, 101.264 | ||
| S. thaiensis | Thailand | CH | 12.774, 102.096 |
| Wu | 14.443, 105.273 | ||
| Tai | 18.894, 98.858 | ||
| CS | 18.84, 99.47 | ||
| QMS | 18.809, 98.914 | ||
| S. singaporensis | Singapore | SNP | 1.312, 103.816 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, D.-M.; Fan, S.; Yu, H. Seven Sycoryctine Fig Wasp Species (Chalcidoidea: Pteromalidae) Associated with Dioecious Ficus hirta Inhabiting South China and Southeast Asia. Biology 2022, 11, 801. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060801
Wong D-M, Fan S, Yu H. Seven Sycoryctine Fig Wasp Species (Chalcidoidea: Pteromalidae) Associated with Dioecious Ficus hirta Inhabiting South China and Southeast Asia. Biology. 2022; 11(6):801. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060801
Chicago/Turabian StyleWong, Da-Mien, Songle Fan, and Hui Yu. 2022. "Seven Sycoryctine Fig Wasp Species (Chalcidoidea: Pteromalidae) Associated with Dioecious Ficus hirta Inhabiting South China and Southeast Asia" Biology 11, no. 6: 801. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060801