
Early Life History and Recruitment Processes of a Tropical Anguillid Eel Anguilla marmorata to the Pacific Coast, as Revealed by Otolith Sr:Ca Ratios and Microstructure


Abstract
:Simple Summary
Abstract
1. Introduction
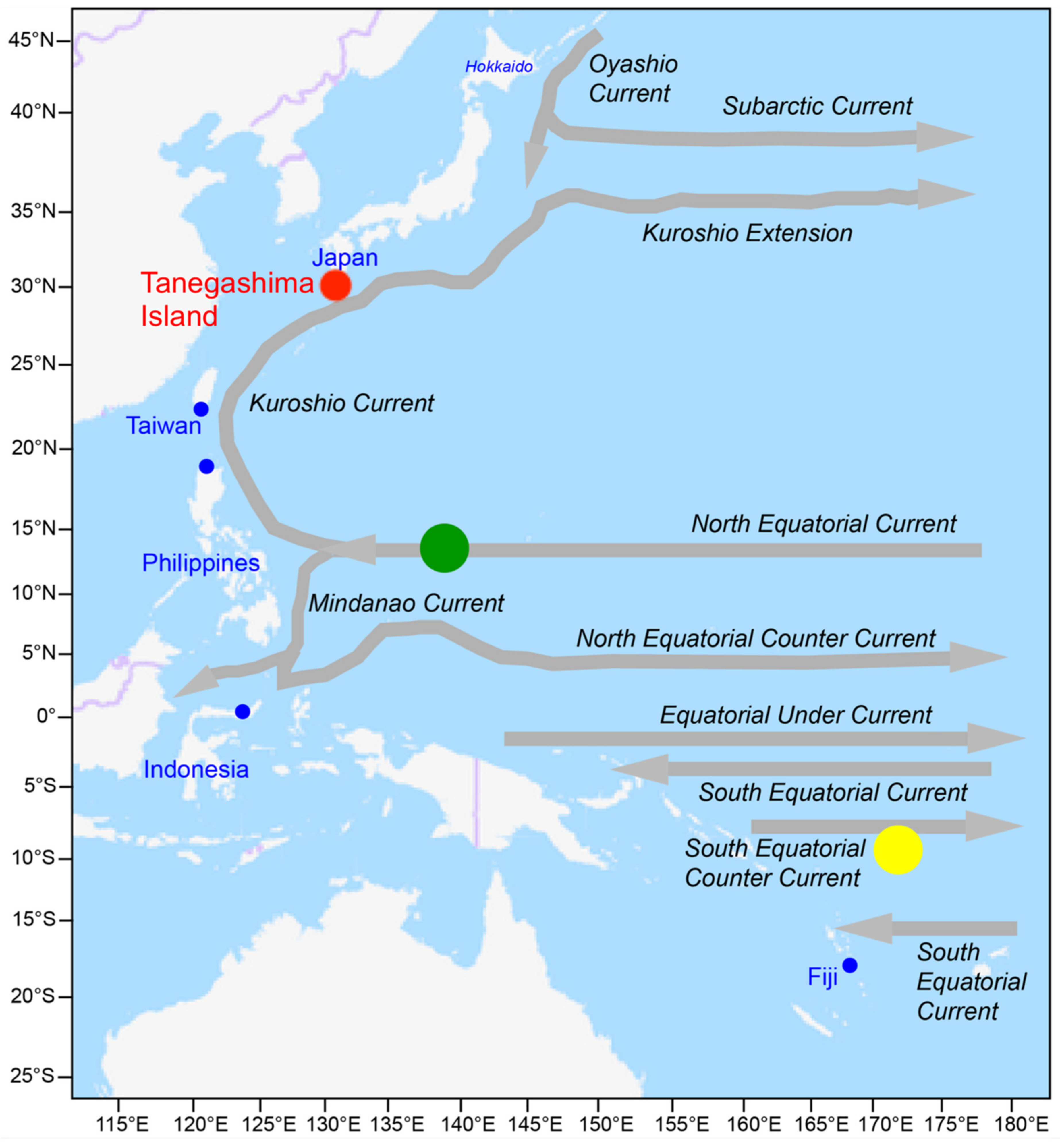
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
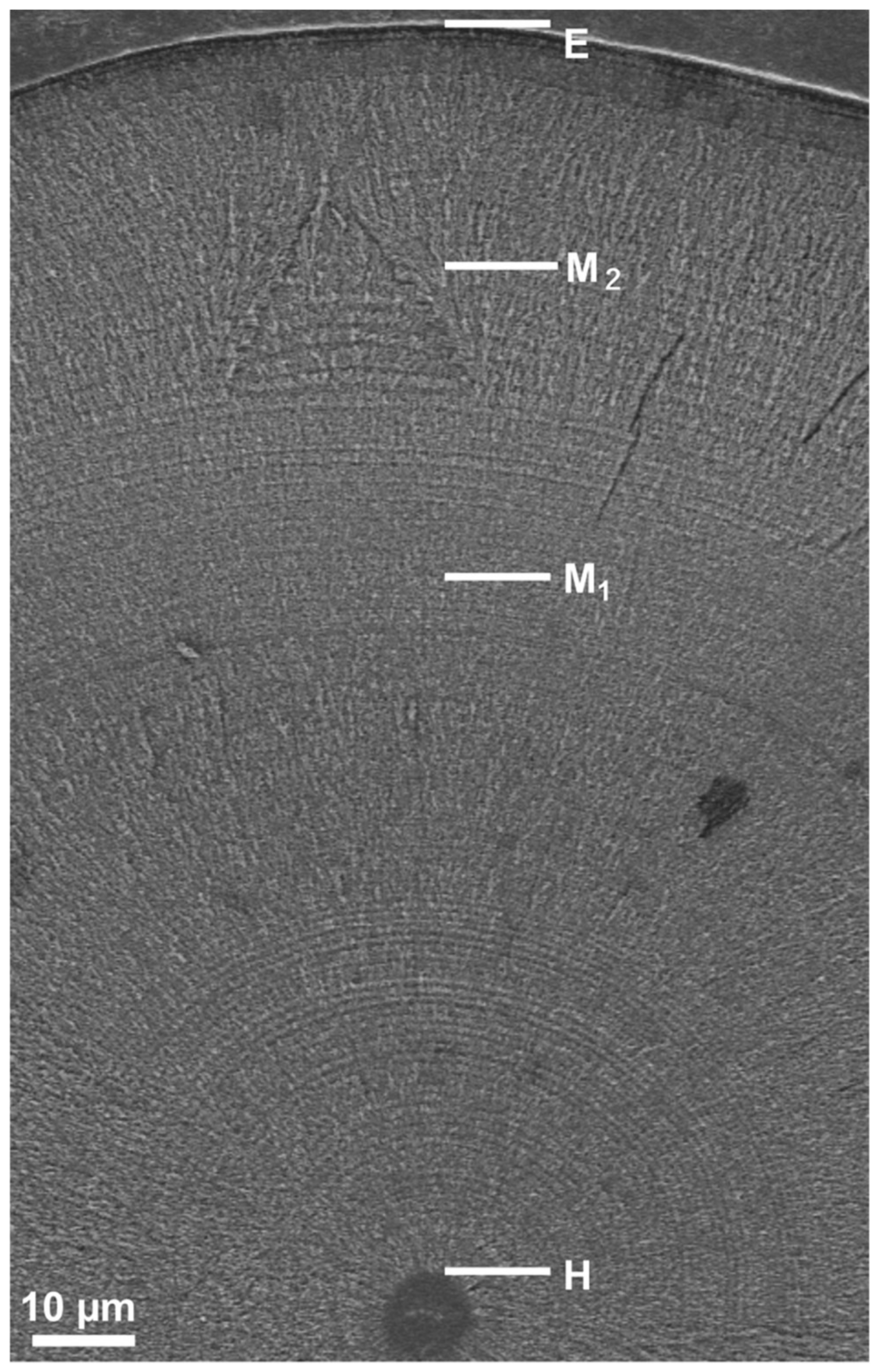
2.3. Otolith Preparation
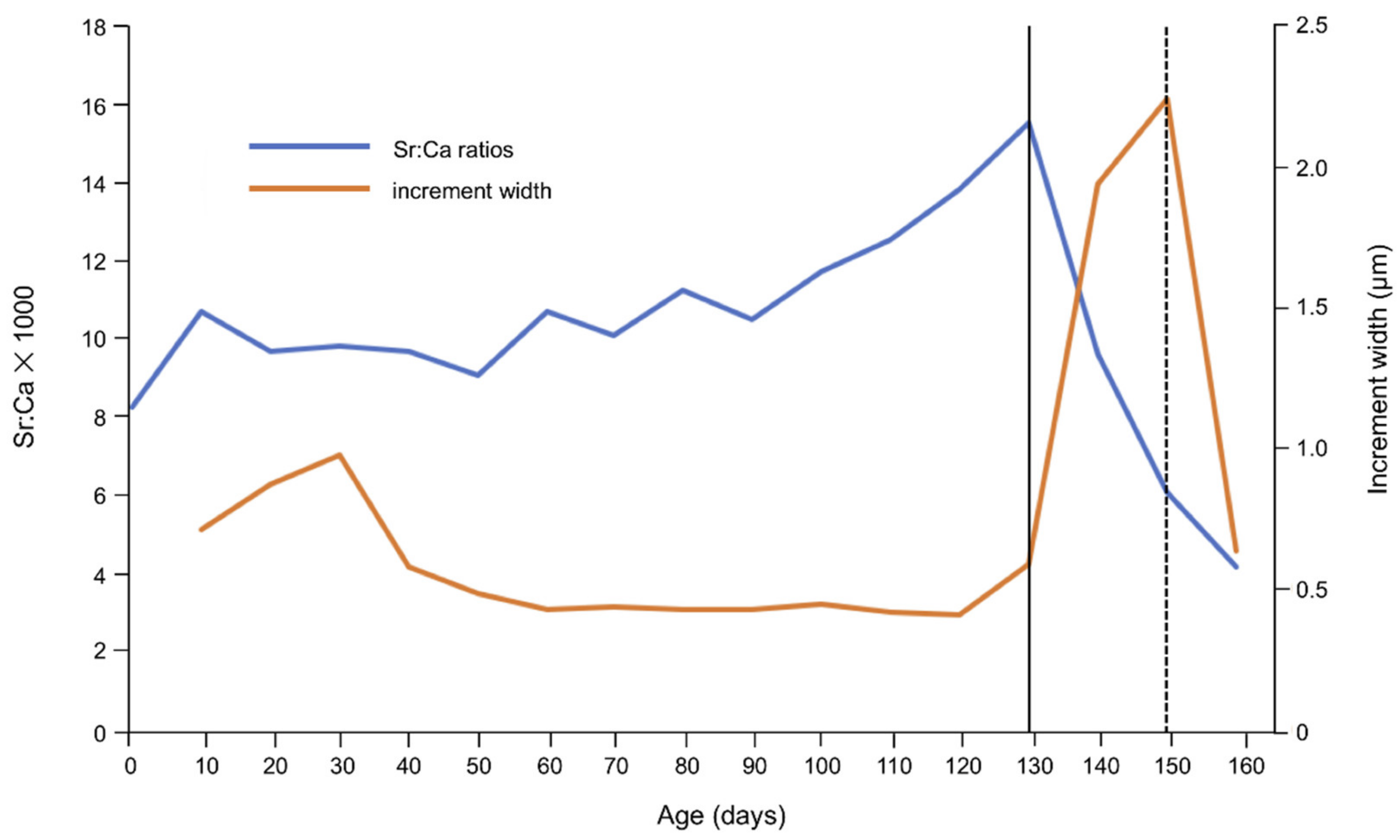
2.4. Otolith Sr and Ca Analyses
2.5. Otolith Increment Analysis
2.6. Interpretation of Early Life History
2.7. Statistics
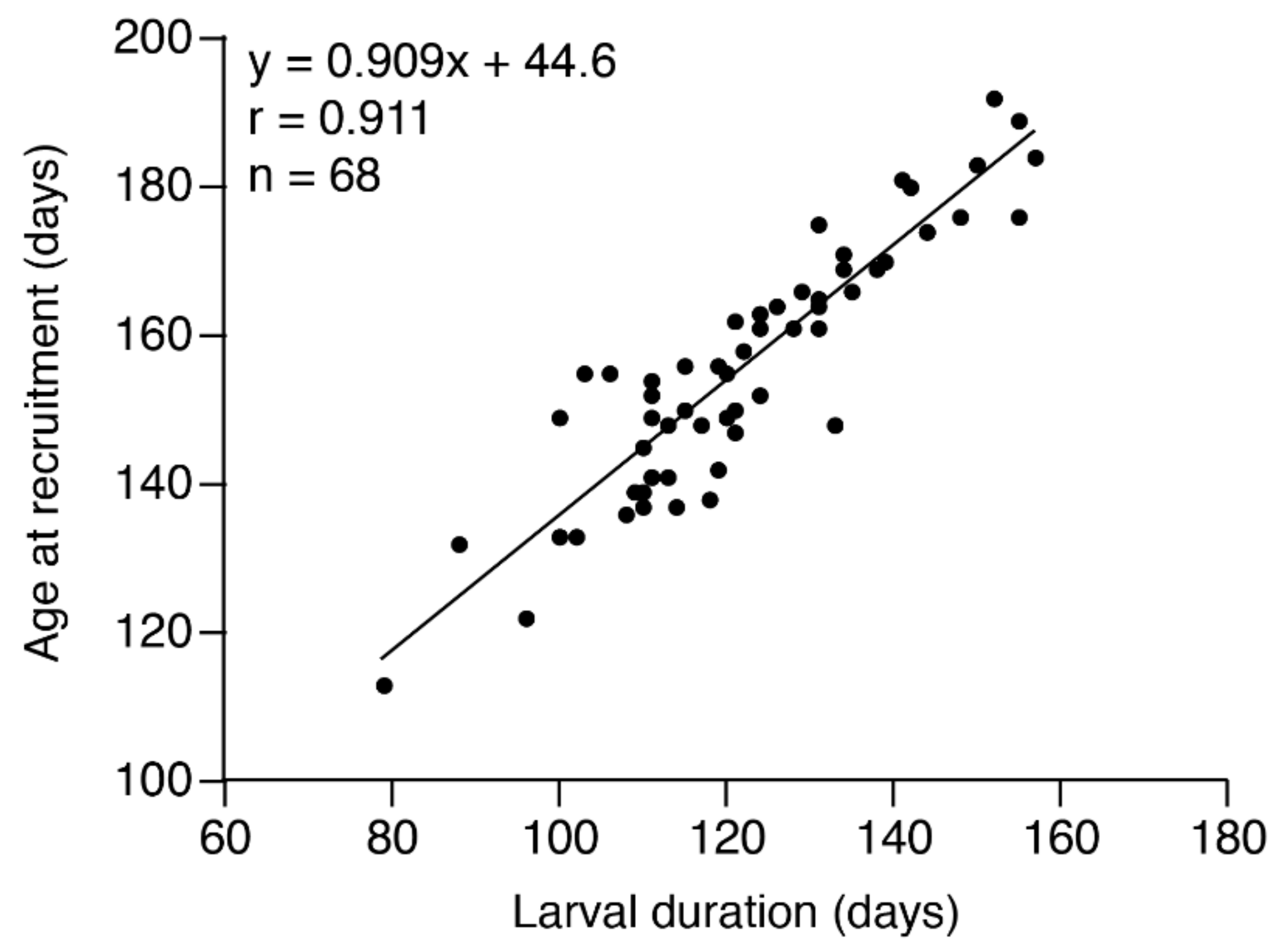
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tesch, F.W. The Eel, 3rd ed.; Blackwell Science: Oxford, UK, 2003. [Google Scholar]
- Arai, T. Taxonomy and Distribution. In Biology and Ecology of Anguillid Eels; Arai, T., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1–20. [Google Scholar]
- Minegishi, Y.; Aoyama, J.; Tsukamoto, K. Multiple population structure of the giant mottled eel, Anguilla marmorata. Mol. Ecol. 2008, 17, 3109–3122. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, P.A.; Minegishi, Y.; Zenboudji, S.; Valade, P.; Aoyama, J.; Berrebi, P. Within-population structure highlighted by differential introgression across semipermeable barriers to gene flow in Anguilla marmorata. Evolution 2011, 65, 3413–3427. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Taha, H.; Mohd-Riduan, M.N.; Mokti, S.S.A. Molecular and morphological evidence for the identity of the giant mottled eel, Anguilla marmorata in Southeast Asia. Trop. Ecol. 2020, 61, 429–436. [Google Scholar] [CrossRef]
- Zan, N.D.; Sarbini, A.; Taha, H.; Tan, I.V.; Azri, A.; Kahar, R.; Metali, F.; Ahmad, N.; Arai, T. Occurrence and ecological implication of a tropical anguillid eel, Anguilla marmorata, in Brunei Darussalam, Borneo Island. Zoologia 2020, 37, e39468. [Google Scholar] [CrossRef]
- Arai, T. Migration ecology in the freshwater eels of the genus Anguilla Schrank, 1798. Trop. Ecol. 2022, 63, 155–170. [Google Scholar] [CrossRef]
- Arai, T. Ecology and evolution of migration in the freshwater eels of the genus Anguilla Schrank, 1798. Heliyon 2020, 6, e05176. [Google Scholar] [CrossRef]
- Campana, S.E. Chemistry and composition of fish otoliths: Pathway, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Sato, H.; Ishii, T.; Tsukamoto, K. Alkaline earth metal and Mn distribution in otoliths of Anguilla spp. glass eels and elvers. Fish. Sci. 2003, 69, 421–423. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N. Diverse migration strategy between freshwater and seawater habitats in the freshwater eels genus Anguilla. J. Fish Biol. 2012, 81, 442–455. [Google Scholar] [CrossRef]
- Arai, T. Effect of salinity on strontium:calcium ratios in the otoliths of Sakhalin taimen, Hucho perryi. Fish. Sci. 2010, 76, 451–455. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N. Influence of water salinity on the strontium:calcium ratios in otoliths of the giant mottled eel, Anguilla marmorata. Environ. Biol. Fish. 2017, 100, 281–286. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N. Contribution of migratory types to the reproduction of migrating silver eels in a tropical eel, Anguilla bicolor bicolor. Heliyon 2022, 8, e09491. [Google Scholar] [CrossRef]
- Arai, T.; Kotake, A.; McCarthy, T.K. Habitat use by the European eel Anguilla anguilla in Irish waters. Estuar. Coast. Shelf Sci. 2006, 67, 569–578. [Google Scholar] [CrossRef]
- Arai, T.; Kotake, A.; Harrod, C.; Morrissey, M.; McCarthy, T.K. Ecological plasticity of the European eel Anguilla anguilla in a tidal Atlantic lake system in Ireland. J. Mar. Biol. Assoc. 2019, 99, 1189–1195. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Arai, T. Facultative catadromy of the eel, Anguilla japonica, between freshwater and seawater habitats. Mar. Ecol. Prog. Ser. 2001, 220, 365–376. [Google Scholar] [CrossRef]
- Arai, T.; Kotake, A.; Ohji, M.; Miller, M.J.; Tsukamoto, K.; Miyazaki, N. Occurrence of sea eels of Anguilla japonica along the Sanriku Coast of Japan. Ichthyol. Res. 2003, 50, 78–81. [Google Scholar] [CrossRef]
- Arai, T.; Kotake, A.; Ohji, M.; Miyazaki, N.; Tsukamoto, K. Migratory history and habitat use of Japanese eel Anguilla japonica in the Sanriku Coast of Japan. Fish. Sci. 2003, 69, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Kotake, A.; Ohji, M. Variation in migratory history of Japanese eels, Anguilla japonica, collected in the northernmost part of its distribution. J. Mar. Biol. Assoc. 2008, 88, 1075–1080. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N.; Kotake, A. Occurrence of estuarine and sea eels Anguilla japonica and a migrating silver eel Anguilla anguilla in Tokyo Bay area, Japan. Fish. Sci. 2009, 75, 1197–1203. [Google Scholar] [CrossRef]
- Kotake, A.; Arai, T.; Ozawa, T.; Nojima, S.; Miller, M.J.; Tsukamoto, K. Variation in migratory history of Japanese eels, Anguilla japonica, collected in coastal waters of the Amakusa Islands, Japan, inferred from otolith Sr/Ca ratios. Mar. Biol. 2003, 142, 849–854. [Google Scholar] [CrossRef]
- Kotake, A.; Okamura, A.; Yamada, Y.; Utoh, T.; Arai, T.; Miller, M.J.; Oka, H.P.; Tsukamoto, K. Seasonal variation in migratory history of the Japanese eel, Anguilla japonica, in Mikawa Bay, Japan. Mar. Ecol. Prog. Ser. 2005, 293, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Chino, N.; Arai, T. Relative contribution of migratory type on the reproduction of migrating silver eels Anguilla japonica, collected off Shikoku Island, Japan. Mar. Biol. 2009, 156, 661–668. [Google Scholar] [CrossRef]
- Lamson, H.M.; Shiao, J.C.; Iizuka, Y.; Tzeng, W.N.; Cairns, D.K. Movement patterns of American eels (Anguilla rostrata) between salt- and freshwater in a coasta watershed, based on otolith microchemistry. Mar. Biol. 2006, 149, 1567–1576. [Google Scholar] [CrossRef]
- Jessop, B.M.; Cairns, D.K.; Thibault, I.; Tzeng, W.N. Life history of American eel Anguilla rostrata: New insights from otolith microchemistry. Aquat. Biol. 2008, 1, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Kotake, A.; Lokman, M.; Miller, M.J.; Tsukamoto, K. Evidence of different habitat use by New Zealand freshwater eel Anguilla australis and A. dieffenbachii as revealed by otolith microchemistry. Mar. Ecol. Prog. Ser. 2004, 266, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Shiao, J.C.; Iizuka, Y.; Chang, C.W.; Tzeng, W.N. Disparities in habitat use and migratory behavior between tropical eel Anguilla marmorata and temperate eel A. japonica in four Taiwanese rivers. Mar. Ecol. Prog. Ser. 2003, 261, 233–242. [Google Scholar] [CrossRef]
- Chino, N.; Arai, T. Migratory history of the giant mottled eel (Anguilla marmorata) in the Bonin Islands of Japan. Ecol. Freshw. Fish. 2010, 19, 19–25. [Google Scholar] [CrossRef]
- Lin, Y.J.; Jessop, B.M.; Weyl, O.L.; Iizuka, Y.; Lin., S.H.; Tzeng, W.N.; Sun, C.L. Regional variation in otolith Sr:Ca ratios of African longfinned eel Anguilla mossambica and mottled eel Anguilla marmorata: A challenge to the classic tool for reconstructing migratory histories of fishes. J. Fish Biol. 2012, 81, 427–441. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N.; Le, D.Q. Migration and habitat use of the tropical eels Anguilla marmorata and A. bicolor pacifica in Vietnam. Aquat. Ecol. 2013, 47, 57–65. [Google Scholar] [CrossRef]
- Arai, T.; Chino, N. Opportunistic migration and habitat use of the giant mottled eel Anguilla marmorata (Teleostei: Elopomorpha). Sci. Rep. 2018, 8, 5666. [Google Scholar] [CrossRef] [Green Version]
- Chino, N.; Arai, T. Occurrence of marine resident tropical eel Anguilla bicolor bicolor in Indonesia. Mar. Biol. 2010, 157, 1075–1081. [Google Scholar] [CrossRef]
- Chino, N.; Arai, T. Habitat use and habitat transitions in the tropical eel, Anguilla bicolor bicolor. Environ. Biol. Fish. 2010, 89, 571–578. [Google Scholar] [CrossRef]
- Arai, T.; Chai, I.J.; Iizuka, Y.; Chang, C.W. Habitat segregation and migration in tropical anguillid eels, Anguilla bengalensis bengalensis and A. bicolor bicolor. Sci. Rep. 2020, 10, 16890. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Limbong, D.; Otake, T.; Tsukamoto, K. Metamorphosis and inshore migration of tropical eels Anguilla spp. in the Indo-Pacific. Mar. Ecol. Prog. Ser. 1999, 182, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Limbong, D.; Otake, T.; Tsukamoto, K. Recruitment mechanisms of tropical eels, Anguilla spp., and implications for the evolution of oceanic migration in the genus Anguilla. Mar. Ecol. Prog. Ser. 2001, 216, 253–264. [Google Scholar] [CrossRef]
- Arai, T.; Otake, T.; Tsukamoto, K. Drastic changes in otolith microstructure and microchemistry accompanying the onset of metamorphosis in the Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 1997, 161, 17–22. [Google Scholar] [CrossRef]
- Arai, T.; Marui, M.; Miller, M.J.; Tsukamoto, K. Growth history and inshore migration of the tropical eel, Anguilla marmorata in the Pacific. Mar. Biol. 2002, 140, 309–316. [Google Scholar]
- Marui, M.; Arai, T.; Miller, M.J.; Jellyman, D.J.; Tsukamoto, K. Comparison of early life history between New Zealand temperate eels and Pacific tropical eels revealed by otolith microstructure and microchemistry. Mar. Ecol. Prog. Ser. 2001, 213, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Marui, M.; Otake, T.; Tsukamoto, K. Inshore migration of a tropical eel, Anguilla marmorata from Taiwanese and Japanese coasts. Fish. Sci. 2002, 68, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Réveillac, É.; Feunteun, E.; Berrebi, P.; Gagnaire, P.A.; Lecomte-Finiger, R.; Bosc, P.; Robinet, T. Anguilla marmorata larval migration plasticity as revealed by otolith microstructural analysis. Can. J. Fish. Aquat. Sci. 2008, 65, 2127–2137. [Google Scholar] [CrossRef]
- Leander, N.; Tzeng, W.N.; Yeh, N.T.; Shen, K.N.; Han, Y.S. Effects of metamorphosis timing and the larval growth rate on the latitudinal distribution of sympatric freshwater eels, Anguilla japonica and A. marmorata, in the western North Pacific. Zool. Stud. 2013, 52, 30. [Google Scholar] [CrossRef] [Green Version]
- Hewavitharane, C.A.; Pickering, T.D.; Rico, C.; Mochioka, N. Early life history of tropical freshwater eels (Anguilla spp.) recruiting to Viti Levu, Fiji Islands, in the western South Pacific. Mar. Freshw. Res. 2020, 71, 452–460. [Google Scholar] [CrossRef]
- Hedges, K.J.; Ludsin, S.A.; Fryer, B.J. Effects of ethanol preservation on otolith microchemistry. J. Fish Biol. 2004, 64, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Bertin, L. Eels, A Biological Study; Cleaver-Hume Press Ltd.: London, UK, 1956. [Google Scholar]
- Miyama, T.; Awaji, T.; Akitomo, K.; Imasato, N. Study of seasonal transport variations in the Indonesian seas. J. Geophys. Res. 1995, 100, 20517–20541. [Google Scholar] [CrossRef]
- Godfrey, J.S. The effect of the Indonesian throughflow on ocean circulation and heat exchange with the atmosphere: A review. J. Geophys. Res. 1996, 101, 12217–12237. [Google Scholar] [CrossRef]
- Oui, G.; Lukas, R. Seasonal and interannual variability of the North Equatorial Current, the Mindanao Current, and the Kuroshio along the Pacific western boundary. J. Geophys. Res. 1996, 101, 12315–12330. [Google Scholar]
- Tsukamoto, K.; Chow, S.; Otake, T.; Kurogi, H.; Mochioka, N.; Miller, M.J.; Aoyama, J.; Kimura, S.; Watanabe, S.; Yoshinaga, T.; et al. Oceanic spawning ecology of freshwater eels in the western North Pacific. Nat. Commun. 2011, 2, 179. [Google Scholar] [CrossRef] [Green Version]
- Schabetsberger, R.; Føkland, F.; Kalfatak, D.; Sichrowsky, U.; Meelis, T.; Aarestrup, K.; Gubili, C.; Sarginson, J.; Boufana, B.; Jehle, R.; et al. Genetic and migratory evidence for sympatric spawning of tropical Pacific eels from Vanuatu. Mar. Ecol. Prog. Ser. 2015, 521, 171–187. [Google Scholar] [CrossRef]
- Sugeha, H.Y.; Shinoda, A.; Marui, M.; Arai, T.; Tsukamoto, K. Validation of otolith daily increments in the tropical eel Anguilla marmorata. Mar. Ecol. Prog. Ser. 2001, 220, 291–294. [Google Scholar] [CrossRef]
- Umezawa, A.; Tsukamoto, K.; Tabeta, O.; Yamakawa, H. Daily growth increments in the larval otolith of the Japanese eel, Anguilla japonica. Jpn. J. Ichthyol. 1989, 35, 440–444. [Google Scholar] [CrossRef]
- Tsukamoto, K. Otolith daily growth increments in the Japanese eel. Bull. Jpn. Soc. Sci. Fish. 1989, 55, 1017–1021. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, A.; Tsukamoto, K. Factors influencing otolith increment formation in Japanese eel, Anguilla japonica T. and S., elvers. J. Fish Biol. 1991, 39, 211–223. [Google Scholar] [CrossRef]
- Martin, M.H. Validation of daily growth increments in otoliths of Anguilla rostrata (Lesueur) elvers. Can. J. Zool. 1995, 73, 208–211. [Google Scholar] [CrossRef]
- Arai, T.; Daniel, L.; Tsukamoto, K. Validation of otolith daily increments in the tropical eel, Anguilla celebesensis. Can. J. Zool. 2000, 78, 1078–1084. [Google Scholar] [CrossRef]
- Arai, T.; Otake, T.; Tsukamoto, K. Timing of metamorphosis and larval segregation of the Atlantic eels Anguilla rostrata and A. anguilla, as revealed by otolith microstructure and microchemistry. Mar. Biol. 2000, 137, 39–45. [Google Scholar] [CrossRef]
- Otake, T.; Ishii, T.; Nakahara, M.; Nakamura, R. Drastic changes in otolith strontium/calcium ratios in leptocephali and glass eels of Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 1994, 112, 189–193. [Google Scholar] [CrossRef]
- Arai, T.; Otake, T.; Jellyman, D.J.; Tsukamoto, K. Differences in the early life history of the Australasian shortfinned eel, Anguilla australis from Australia and New Zealand, as revealed by otolith microstructure and microchemistry. Mar. Biol. 1999, 135, 381–389. [Google Scholar] [CrossRef]
- Shiao, J.C.; Tzeng, W.N.; Collins, A.; Iizuka, Y. Role of marine larval duration and growth rate of glass eels in determining the distribution of Anguilla reinhardtii and A. australis on Australian eastern coasts. Mar. Freshw. Res. 2002, 53, 687–695. [Google Scholar] [CrossRef]
- Arai, T.; Aoyama, J.; Ishikawa, S.; Miller, M.J.; Otake, T.; Inagaki, T.; Tsukamoto, K. Early life history of tropical Anguilla leptocephali in the western Pacific Ocean. Mar. Biol. 2001, 138, 887–895. [Google Scholar]
- Arai, T.; Miller, M.J.; Tsukamoto, K. Larval duration of the tropical eel, Anguilla celebesensis, from the Indonesian and Philippine coasts. Mar. Ecol. Prog. Ser. 2003, 251, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Otake, T.; Limbong, D.; Tsukamoto, K. Early life history and recruitment of the tropical eel Anguilla bicolor pacifica, as revealed by otolith microstructure and microchemistry. Mar. Biol. 1999, 133, 319–326. [Google Scholar] [CrossRef]
- Pfeiler, E. Glycosaminoglycan breakdown during metamorphosis of larval bonefish Albla. Mar. Biol. Lett. 1984, 5, 241–249. [Google Scholar]
- Arai, T.; Abdul Kadir, S.R.; Chino, N. Year-round spawning by a tropical catadromous eel Anguilla bicolor bicolor. Mar. Biol. 2016, 163, 37. [Google Scholar] [CrossRef]
- Arai, T.; Abdul Kadir, S.R. Opportunistic spawning of tropical anguillid eels Anguilla bicolor bicolor and A. bengalensis bengalensis. Sci. Rep. 2017, 7, 41649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinet, T.; Lecomte-Finiger, R.; Escoubeyrou, K.; Feunteun, E. Tropical eels Anguilla spp. recruiting to Réunion Island in the Indian Ocean: Taxonomy, patterns of recruitment and early life histories. Mar. Ecol. Prog. Ser. 2003, 259, 263–272. [Google Scholar] [CrossRef]
- Réveillac, É.; Robinet, T.; Rabenevanana, M.W.; Valade, P.; Feunteun, É. Clues to the location of the spawning area and larval migration characteristics of Anguilla mossambica as inferred from otolith microstructural analyses. J. Fish Biol. 2009, 74, 1866–1877. [Google Scholar] [CrossRef]
- Tabeta, O.; Konishi, Y. An anguillid leptocephalus Anguilla japonica from the waters of Miyako Island, Okinawa, Japan. Bull. Jpn. Soc. Sci. Fish. 1986, 52, 1935–1937. [Google Scholar] [CrossRef] [Green Version]
- Kleckner, R.C.; McCleave, J.D. Spatial and temporal distribution of American eel larvae in relation to North Atlantic Ocean current systems. Dana 1985, 4, 67–92. [Google Scholar]
- Umezawa, A.; Tsukamoto, K. Age and birth date of the glass eel, Anguilla japonica, collected in Taiwan. Bull. Jpn. Soc. Sci. Fish. 1990, 56, 1199–1202. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, K. Recruitment mechanism of the eel, Anguilla japonica, to the Japanese coast. J. Fish Biol. 1990, 36, 659–671. [Google Scholar] [CrossRef]
- Arai, T.; Taha, H. Contrasting patterns of genetic population structure in tropical freshwater eels of genus Anguilla in the Indo-Pacific. Heliyon 2021, 7, e07097. [Google Scholar] [CrossRef] [PubMed]
- Arai, T. Evidence of local short-distance spawning migration of tropical freshwater eels, and implications for the evolution of freshwater eel migration. Ecol. Evol. 2014, 4, 3812–3819. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Number of Specimens | Total Length (mm) | Otolith Radius (µm) | Duration of Larval Stage (Days) | Duration of Metamorphosis Stage (Days) | Age at Recruitment (Days) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | ||
| December 1998 | 18 | 49.9 ± 1.2 | 48.0–52.1 | 149.7 ± 8.4 | 137.1–163.3 | 120.1 ± 25.5 | 79–157 | 15.9 ± 3.2 | 10–20 | 150.1 ± 22.9 | 113–184 |
| January 1999 | 4 | 50.7 ± 0.8 | 49.8–51.5 | 141.1 ± 6.9 | 133.1–149.3 | 122.3 ± 15.5 | 103–138 | 21.8 ± 5.7 | 14–27 | 161.8 ± 12.4 | 148–175 |
| February 1999 | 25 | 49.2 ± 2.0 | 44.9–54.2 | 151.6 ± 6.6 | 138.6–162.7 | 123.1 ± 13.4 | 100–155 | 17.2 ± 4.0 | 12–24 | 154.3 ± 17.0 | 133–189 |
| March 1999 | 21 | 49.1 ± 1.8 | 44.8–52.0 | 148.6 ± 6.8 | 138.2–163.8 | 123.5 ± 12.8 | 100–152 | 20.6 ± 3.4 | 13–27 | 160.3 ± 10.7 | 148–192 |
| Country | Total Length (mm) | Duration of Larval Stage (Days) | Age at Recruitment (Days) | Reference | |||
|---|---|---|---|---|---|---|---|
| Mean | Range | Mean | Range | Mean | Range | ||
| Fiji | 51.0–51.5 | 48.0–56.3 | 115–113 | 101–146 | 152–154 | 124–183 | [44] |
| Indonesia | 50.9 | 47.9–54.8 | 120 | 96–147 | 152 | 129–177 | [36] |
| 47.9–52.3 | 128 | 114–158 | 155 | 144–182 | [37] | ||
| Japan | 49.2–49.7 | 45.1–54.2 | 123 | 100–155 | 154 | 133–189 | [39,41] |
| 46.7 | 118 | 145 | [43] | ||||
| Philippines | 49.9 | 47.2–51.6 | 120 | 105–140 | 154 | 136–178 | [36] |
| 49.5 | 110 | 145 | [43] | ||||
| Taiwan | 50.2–50.3 | 48.0–53.4 | 116 | 92–145 | 145 | 116–167 | [39,41] |
| 51.6 | 112 | 134 | [43] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arai, T. Early Life History and Recruitment Processes of a Tropical Anguillid Eel Anguilla marmorata to the Pacific Coast, as Revealed by Otolith Sr:Ca Ratios and Microstructure. Biology 2022, 11, 803. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060803
Arai T. Early Life History and Recruitment Processes of a Tropical Anguillid Eel Anguilla marmorata to the Pacific Coast, as Revealed by Otolith Sr:Ca Ratios and Microstructure. Biology. 2022; 11(6):803. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060803
Chicago/Turabian StyleArai, Takaomi. 2022. "Early Life History and Recruitment Processes of a Tropical Anguillid Eel Anguilla marmorata to the Pacific Coast, as Revealed by Otolith Sr:Ca Ratios and Microstructure" Biology 11, no. 6: 803. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11060803