
Contribution of Soil Type to Quantity and Nutritional Value of Grass Species on the South African Highveld


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
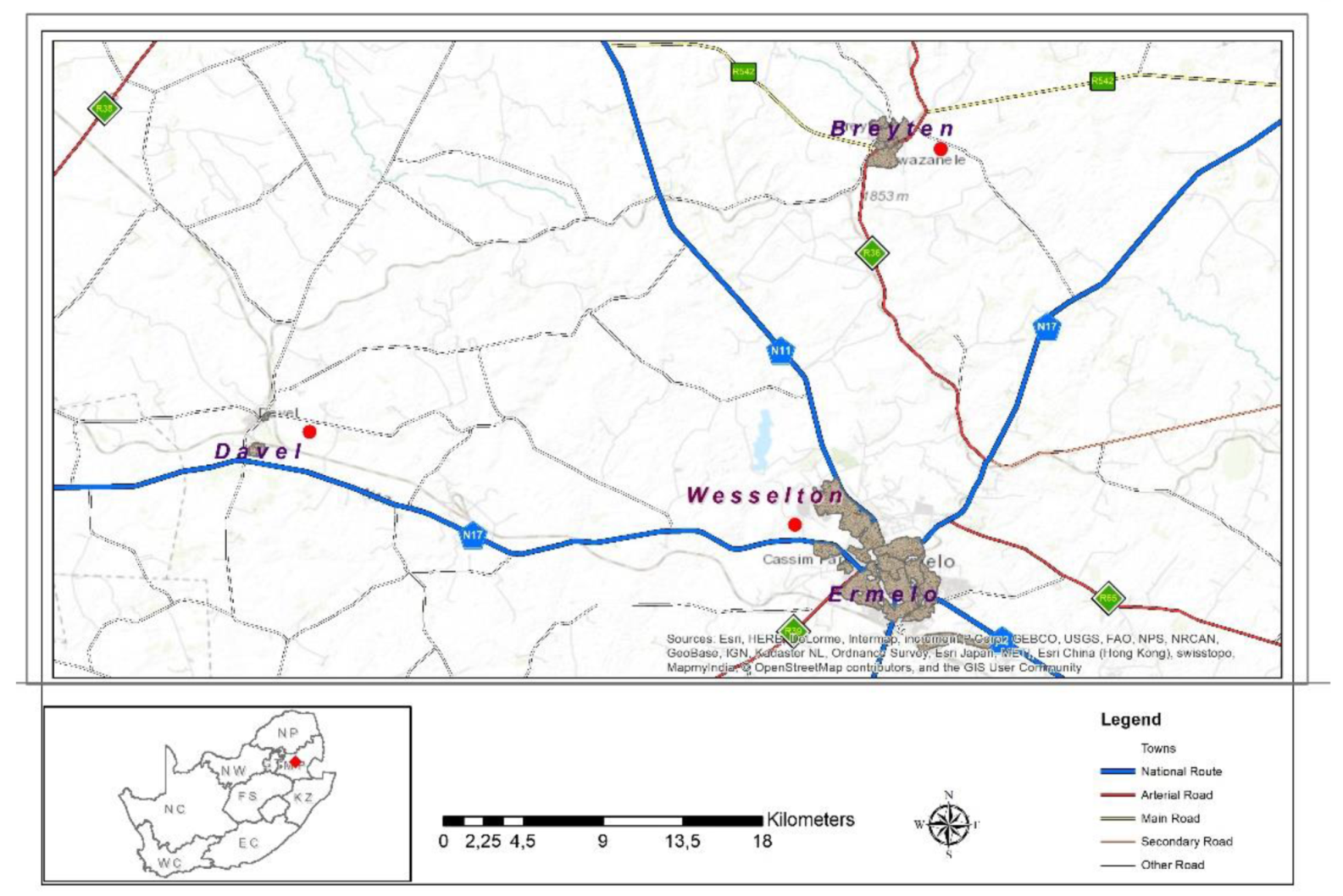
2.1. Study Sites
2.2. Soil and Grass Sampling and Species Identification
2.3. Chemical Composition
2.4. In Vitro Ruminal Dry Matter Degradability
2.5. Statistical Analysis
3. Results
4. Discussion
4.1. Soil Parameters
4.2. Grass Species Composition and Distribution
4.3. Biomass and the Species Grazing Values
4.4. Chemical Composition of Common and Dominant Grass Species in Different Soil Types
4.5. In vitro Ruminal Dry Matter Degradability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selemani, I.S.; Eik, L.O.; Holand, Ø.; Ådnøy, T.; Mtengeti, E.; Mushi, D. The effects of a deferred grazing system on rangeland vegetation in a North-Western, Semi-Arid region of Tanzania. Afr. J. Range Forage Sci. 2013, 30, 141–148. [Google Scholar] [CrossRef]
- Andualem, D.; Negesse, T.; Tolera, A. Chemical composition, in vitro organic matter digestibility and kinetics of rumen dry matter degradability of morphological fractions of stinging nettle (Urtica simensis). Adv. Biol. Res. 2016, 10, 183–190. [Google Scholar]
- Matney, C.A. Effects of grazing on grass. In Proceedings of the The Range Beef Cow Symposium, Loveland, CO, USA, 17–19 November 2015. [Google Scholar]
- Kebede, G.; Feyissa, F.; Assefa, G.; Alemayehu, M.; Mengistu, A.; Kehaliew, A.; Melese, K.; Mengistu, S.; Tadesse, E.; Wolde, S.; et al. Chemical composition and in-vitro organic matter digestibility of napier grass (Pennisetum purpureum (L.) Schumach) accessions in the Mid and Highland areas of Ethiopia. Int. J. Livest. Res. 2016, 6, 1–20. [Google Scholar] [CrossRef]
- Khan, N.A.; Rahman, S.U.; Cone, J.W. Chemical composition, ruminal degradation kinetics and methane production (in vitro) potential of local and exotic grass species grown in Peshawar. Pak. J. Bot. 2020, 52, 161–166. [Google Scholar]
- Augustine, D.J.; Frank, D.A. Effects of migratory grazers on spatial heterogeneity of soil nitrogen properties in a grassland ecosystem. Ecology 2001, 82, 3149–3162. [Google Scholar] [CrossRef]
- Wei, L.; Hai-Zhou, H.; Zhi-Nan, Z.; Gao-Lin, W. Effects of grazing on the soil properties and C and N storage in relation to biomass allocation in an alpine meadow. J. Soil Sci. Plant Nutr. 2001, 11, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Bilotta, G.S.; Brazier, R.E.; Haygarth, P.M. The impacts of grazing animals on the quality of soils, vegetation, and surface waters in intensively managed grasslands. Adv. Agron. 2007, 94, 237–280. [Google Scholar]
- Bullock, J.M.; Marriott, C.A. Plant responses to grazing, and opportunities for manipulation. In Grazing Management, the Principles, and Practice of Grazing, for Profit and Environmental Gain, within Temperate Grassland Systems, Proceedings of the British Grassland Society Conference: BGS Occasional Symposium 34. Reading, Harrogate, UK, 29 Febraury–2 March 2000; Rook, A.J., Penning, P.D., Eds.; British Grassland Society: Harrogate, UK, 2000; pp. 17–26. [Google Scholar]
- Roodt, V. Grasses of the Kruger National Park and Surrounding Bushveld; Veronica Roodt Publications: Hartbeespoort, South Africa, 2011. [Google Scholar]
- Kohler, F.; Gillet, F.; Gobat, J.M.; Buttler, A. Seasonal vegetation changes in mountain pastures due to simulated effects of cattle grazing. J. Veg. Sci. 2004, 15, 143–150. [Google Scholar] [CrossRef]
- Zhang, B.; Beck, R.; Pan, Q.; Zhao, M.; Hao, X. Soil physical and chemical properties in response to long-term cattle grazing on sloped rough fescue grassland in the foothills of the Rocky Mountains, Alberta. Geoderma 2019, 346, 75–83. [Google Scholar] [CrossRef]
- Taboada, M.A.; Rubio, G.; Chaneton, E.J. Grazing impacts on soil physical, chemical, and ecological properties in forage production systems. In Soil Management: Building a Stable Base for Agriculture; Wiley: Hoboken, NJ, USA, 2011; Chapter 20; pp. 301–320. [Google Scholar]
- Mayel, S.; Jarrah, M.; Kuka, K. How does grassland management affect physical and biochemical properties of temperate grassland soils? A review study. Grass Forage Sci. 2021, 76, 215–244. [Google Scholar] [CrossRef]
- Chiepe, Q.M.; Amantle, I.A.; Amos, K.R. A review on grazing management systems and effects on savanna ecosystem dynamics. Afr. J. Ecol. Ecosyst. 2017, 4, 251–257. [Google Scholar]
- Msiza, N.H.; Ravhuhali, K.E.; Mokoboki, H.K.; Mavengahama, S.; Motsei, L.E. Ranking species for veld restoration in semi-arid regions using agronomic, morphological and chemical parameters of selected grass species at different developmental stages under controlled environment. Agronomy 2021, 11, 1–14. [Google Scholar]
- Murillo, M.; Herrera, E.; Carrete, F.O.; Ruiz, O.; Serrato, J.S. Chemical composition, in vitro gas production, ruminal fermentation and degradation patterns of diets by grazing steers in native range of North Mexico. Asian-Australas. J. Anim. Sci. 2012, 25, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- An, P.; Li, X.; Zheng, Y.; Eneji, A.E.; Qiman, Y.; Zheng, M.; Inanaga, S. Distribution of plant species and species-soil relationship in the east central Gurbantunggut Desert, China. J. Geog. Sci. 2015, 25, 101–112. [Google Scholar] [CrossRef]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- AgriLASA. Feed and Plant Analysis Methods; Agricultural Laboratory Association of Southern Africa: Pretoria, South Africa, 1998. [Google Scholar]
- Baker, K.F. The determination of organic carbon in soil using a protein colorimeter. Lab. Pract. 1976, 25, 82–83. [Google Scholar]
- Van Oudtshoorn, F.P. Guide to Grasses of Southern Africa; Briza Publication: Pretoria, South Africa, 2014. [Google Scholar]
- Moyo, C.S.; Campbell, B.M. Grass species composition, yield and quality under and outside tree crowns in a semi-arid rangeland in south-western Zimbabwe. Afr. J. Range Forage Sci. 1998, 15, 23–34. [Google Scholar] [CrossRef]
- Jawuoro, S.O.; Koech, O.K.; Karuku, G.N.; Mbau, J.S. Plant species composition and diversity depending on piospheres and seasonality in the southern rangelands of Kenya. Ecol. Process. 2017, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Ravhuhali, K.E. Spatial Variation in Density, Species Composition and Nutritive Value of Vegetation in Selected Communal Areas. Ph.D. Thesis, Department of Animal Science, North-West University, Mmabatho, South Africa, 2018. [Google Scholar]
- Ravhuhali, K.E.; Moyo, B.; Mmbi, N.; Silinda, D. Effect of soil type on herbaceous species composition and distribution in community property association projects of Bela-Bela municipality, South Africa. Trop. Agric. 2021, 97, 104–117. [Google Scholar]
- Du Toit, J.T. Feeding-height stratification among African browsing ruminants. Afr. J. Ecol. 1990, 28, 55–61. [Google Scholar] [CrossRef]
- AOAC. Method number 973.18. In Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- AOAC. Method number 976.06. In Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- SAS. Statistical Analytical Systems Users Guide; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Pietri, J.A.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Tefera, S.B. Rangeland degradation in semi-arid Swaziland: Effects of dip-tanks on herbaceous vegetation and soil properties. Afr. J. Range Forage Sci. 2013, 30, 127–140. [Google Scholar] [CrossRef]
- Beyene, S.T.; Mlambo, V. Botanical and chemical composition of common grass species around di-tank areas in Semi-Arid communal rangelands of Swaziland. Trop. Subtrop. Agroecosyst. 2012, 15, 142–143. [Google Scholar]
- Matsumoto, S.; Shimada, H.; Sasaoka, T.; Miyajima, I.; Kusuma, G.J.; Gautama, R.S. Effects of acid soils on plant growth and successful revegetation in the case of mine site. In Soil pH for Nutrient Availability and Crop Performance; IntechOpen: London, UK, 2017; pp. 9–27. [Google Scholar]
- Lončarić, Z.; Karalić, K.; Popović, B.; Rastija, D.; Vukobratović, M. Total and plant available micronutrients in acidic and calcareous soils in Croatia. Cereal Res. Commun. 2008, 36, 331–334. [Google Scholar]
- Ch’ng, H.Y.; Ahmed, O.H.; Majid, N.M.A. Improving phosphorus availability in an acid soil using organic amendments produced from agroindustrial wastes. Sci. World J. 2014, 506356, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lempesi, A.; Hormoya, E.; Orfanoudakis, M.; Korakis, G.; Kyriazopoulos, A.P. Soil properties along grazing gradients in an open canopy oak forest. In Proceedings of the 9th European Dry Grassland Meeting, Prespa, Greece, 19–23 May 2012. [Google Scholar]
- Horwath, W.R.; Kuzyako, Y. Chapter three—The potential for soils to mitigate climate change through carbon sequestration. Dev. Soil Sci. 2018, 35, 61–92. [Google Scholar]
- Thomas, A.D. Impact of grazing intensity on seasonal variations in soil organic carbon and soil CO2 efflux in two Semi-Arid grasslands in Southern Botswana. Biol. Sci. 2012, 367, 3076–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Y.; Kadambot, H.M.; Siddique, K.H.M.; Turner, N.C.; GangLi, X.; YiNiu, J.; Yang, C.; Liu, L.; Chai, Q. Chapter seven-Ridge-Furrow mulching systems—An innovative technique for boosting crop productivity in Semi-Arid rain—fed environments. Adv. Agron. 2013, 118, 429–476. [Google Scholar]
- Fenton, M.; Albers, C.; Ketterings, Q. Agronomy Fact Sheet Series. Soil Organic Matter; Cornell University Cooperative Extension: Ithaca, NY, USA, 2008; pp. 1–2. [Google Scholar]
- Seabi, W.M. The Relative Influence of Rainfall, Topographical Position and Distance from Village on Composition and Structure of Herbaceous Vegetation in a Communal Rangeland of Bushbuckridge. Master’s Thesis, University of Witwatersrand, Johannesburg, South Africa, 2017. [Google Scholar]
- Siyabulela, S.; Tefera, S.; Wakindiki, I.; Keletso, M. Comparison of grass and soil conditions around water points in different land use systems in Semi-Arid South African rangelands and implications for management and current rangeland paradigms. Arid Land Res. Manag. 2020, 34, 207–230. [Google Scholar] [CrossRef]
- Beck, P.; Jennings, J.; Rogers, J. Management of pastures in the upper south: The I-30 and I-40 Corridors. In Management Strategies for Sustainable Cattle Production in Southern Pastures; Rouquette, M., Jr., Aiken, G., Eds.; Academic Press-Elsevier: Cambridge, MA, USA, 2020; pp. 189–226. [Google Scholar]
- Nawaz, H.; Javed, I.; Hameed, M. Mineral profile, palatability and digestibility of marginal land grasses of Trans-Himalayan grasslands of Pakistan. Pak. J. Bot. 2008, 40, 237–248. [Google Scholar]
- Gusha, J.; Masocha, A.D.M.; Mugabe, P.H. Impact of grazing system on rangeland condition and grazing capacity in Zimbabwe. Rangel. J. 2017, 39, 219–225. [Google Scholar] [CrossRef]
- Kioko, J.; Kiringe, J.W.; Seno, S.O. Impacts of livestock grazing on Savannah grassland in Kenya. J. Arid Lands 2012, 4, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Dalle, G.; Maass, B.L.; Isselstein, J. Relationships between vegetation composition and environmental variables in the Borana rangelands, Southern Oromia, Ethiopia. Ethiop. J. Sci. 2014, 37, 1–12. [Google Scholar]
- Abusuwar, A.O.; Ahmed, E.O. Seasonal variability in nutritive value of ruminant diets under open grazing system in the Semi-Arid rangeland of Sudan (South Darfur State). J. Agric. Biol. 2010, 1, 243–249. [Google Scholar] [CrossRef]
- Deng, L.; Sweeney, S.; Shangguan, Z.P. Grassland responses to grazing disturbance: Plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci. 2013, 69, 524–533. [Google Scholar] [CrossRef]
- Ravhuhali, K.E.; Motsepe, K.; Mnisi, C.M.; Sebolai, T. Chemical composition and in vitro ruminal DM degradability of native pasture grasses and their plant parts. Range Manag. Agroforest. 2018, 39, 243–250. [Google Scholar]
- Kwaza, A.; Tefera, S.; Mlambo, V.; Keletso, M. Short-term grazing exclusion impacts using brush packs on soil and grass layers in degraded communal rangelands of semi-arid South Africa and implications for restoration and pasture utilization. Trop. Grassl. 2020, 8, 220–233. [Google Scholar] [CrossRef]
- Bansal, S.; James, J.J.; Sheley, R.L. The effects of precipitation and soil type on three invasive annual grasses in the western United States. J. Arid Environ. 2014, 104, 38–42. [Google Scholar] [CrossRef]
- Kotze, E. Response of Soil Properties to Rangeland Use in Grassland and Savannah Biomes of South Africa. Ph.D. Thesis, Department of Soil, Crop and Climate Sciences, University of the Free State, Bloemfontein, South Africa, 2015. [Google Scholar]
- Gizachew, L.; Smith, G.N. The status and importance of crude protein and macro minerals in native pastures growing on Vertisols of the central highlands of Ethiopia. J. Environ. Manag. 2012, 93, 177–184. [Google Scholar] [CrossRef]
- Rambau, M.D.; Fushai, F.; Baloyi, J.J. Productivity, chemical composition and ruminal degradability of irrigated Napier grass leave harvested at three stages of maturity. S. Afr. J. Anim. Sci. 2016, 46, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Evitayani, E.; Warly, L.; Fariani, A.; Ichinohe, T.; Fujihara, T. Seasonal changes in nutritive value of some grass species in West Sumatra, Indonesia. Asian-Australas. J. Anim. Sci. 2004, 17, 1663–1668. [Google Scholar] [CrossRef]
- Gargano, A.O.; Adúriz, M.A.; Arelovich, H.M.; Amela, M.I. Forage yield and nutritive value of Eragrostis curvula and Digitaria eriantha in central-south semi-arid Argentina. Trop. Grassl. 2001, 35, 161–167. [Google Scholar]
- Distel, R.A.; Didoné, N.G.; Moretto, A.S. Variations in chemical composition associated with tissue aging in palatable and unpalatable grasses native to central Argentina. J. Arid Environ. 2005, 62, 351–357. [Google Scholar] [CrossRef]
- Herrero, M.; Havlík, P.; Valin, H.; Notenbaert, A.; Rufino, M.C.; Thornton, P.K.; Blümmel, M.; Weiss, F.; Grace, D.; Obersteiner, M. Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20888–20893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravhuhali, K.E.; Ng’ambi, J.W.; Norris, D. Chemical composition and enzymatic in vitro digestibility of cowpea cultivars and buffalo grass hay grown in Limpopo province of South Africa. Livest. Res. Rural Dev. 2010, 22, 1–8. [Google Scholar]
- Ravhuhali, K.E.; Ng’ambi, J.W.; Norris, D.; Ayodele, V.I. The feeding value of four cowpea hay cultivars and effect of their supplementation on intake and digestibility of buffalo grass hay fed to Pedi goats. Asian J. Anim. Vet. Adv. 2011, 6, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.G.O.; Velasco, F.O.; Faria Júnior, W.G.; Teixeira, A.M.; Machado, F.S.; Magalhães, F.A.; Jayme, D.G.; Gonçalves, L.C. In situ degradation kinetic of Andropogon Gayanus grass silages harvested at three stages of maturity. Braz. J. Vet. Anim. Sci. 2014, 66, 1883–1890. [Google Scholar]
- Yayneshet, T.; Eik, L.O.; Moe, S.R. Seasonal variations in the chemical composition and dry matter degradability of exclosure forages in the Semi-Arid region of Northen Ethiopia. Anim. Feed Sci. Technol. 2009, 148, 12–33. [Google Scholar] [CrossRef]
- Jančíc, F.; Koukolová, V.; Homolka, P. Ruminal degradadability of dry matter and neutral detergent fibre of grasses. Czech. J. Anim. Sci. 2010, 55, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.O.; Coates, D.B.; Whitbread, A.M.; Clem, R.L.; Robertson, M.J.; Pengelly, B.C. Seasonal changes in pasture quality and diet selection and their relationship with live-weight gain of steers grazing tropical grass and grass–legume pastures in northern Australia. Anim. Prod. Sci. 2009, 49, 983–993. [Google Scholar] [CrossRef]


{kind=link}
| Site | ||||
|---|---|---|---|---|
| Soil Properties | Breyten | Davel | Wesselton | SEM |
| Type | Hutton soil | Avalon soil | Clovelly soil | |
| pH value | 3.6 b | 4.5 a | 3.5 b | 0.24 |
| Nitrogen nitrate (mg/kg) | 0.77 a | 0.53 b | 0.03 c | 0.03 |
| Nitrogen-peroxynitric acid (mg/kg) | 4.8 a | 5.2 a | 5.2 a | 0.13 |
| Carbon (%) | 1.6 a | 0.5 b | 1.3 a | 0.12 |
| Extractable acidity (%) | 0.90 a | 0.28 c | 0.57 b | 0.02 |
| Extractable alkalinity (%) | 0.70 a | 0.30 c | 0.33 b | 0.02 |
| Sand (%) | 76.3 ab | 74.3 b | 77.7 a | 0.88 |
| Silt (%) | 9.3 a | 9.0 b | 9.0 a | 0.61 |
| Clay (%) | 13.7 b | 15.7 a | 12.3 c | 0.33 |
| Macro-minerals (mg/kg) | ||||
| Potassium | 110.7 a | 91.3 b | 79.0 c | 3.50 |
| Calcium | 233.0 c | 486.7 a | 401.7 b | 6.50 |
| Magnesium | 91.3 b | 107.7 a | 112.7 a | 2.80 |
| Phosphorus | 3.3 b | 9.3 b | 10.3 a | 2.20 |
| Sodium | 3.7 b | 2.7 b | 28.0 a | 0.72 |
| Micro-minerals (mg/kg) | ||||
| Iron | 74.6 b | 53.3 c | 135.9 a | 5.50 |
| Copper | 2.8 a | 1.4 c | 2.4 b | 0.10 |
| Zinc | 1.6 c | 3.7 b | 11.5 a | 0.30 |
| Aluminium | 54.6 a | 9.4 c | 27.7 b | 0.48 |
| Manganese | 25.3 b | 28.5 ab | 32.1 a | 1.10 |
| Abundance 4 | ||||||
|---|---|---|---|---|---|---|
| Grass Species | Life Form 1 | Ecological Status 2 | Grazing Value 3 | HS | AS | CS |
| Aristida congesta | WP | Inc ii | NP | C | C | R |
| Andropogon schirensis | TP | Inc i | MP | C | P | P |
| Brachiaria serrata | TP | Dec | MP | P | P | P |
| Ctenium concinnum | TP | Inc i | NP | R | P | P |
| Cynodon dactylon | CG | Inc ii | HP | R | C | D |
| Diheteropogon amplectens | TP | Dec | MP | R | P | P |
| Digitaria eriantha | TP | Dec | HP | D | R | C |
| Digitaria setifolia | TP | Inc ii | NS | P | P | P |
| Eragrostis capensis | TP | Inc ii | MP | R | P | P |
| Eragrostis chloromelas | TP | Inc ii | MP | C | C | D |
| Eragrostis curvula | TP | Inc ii | MP | C | D | C |
| Eragrostis gammiflua | TP | Inc ii | NP | R | C | C |
| Eragrostis micrantha | TP | Inc ii | NS | R | R | P |
| Elionurus muticus | TP | Inc iii | MP | C | P | P |
| Eragrostis plana | TP | Inc ii | NP | C | D | D |
| Eragrostis planiculmis | TP | Inc ii | NS | C | P | P |
| Eragrostis racemosa | TP | Inc ii | MP | C | P | R |
| Eragrostis rigidior | TP | Inc ii | MP | D | P | P |
| Heteropogon contortus | TP | Inc ii | MP | C | P | R |
| Harpochloa falx | TP | Inc i | NP | C | P | P |
| Hyperrhenia hitra | TP | Inc i | MP | P | C | R |
| Microchloa caffra | TP | Inc ii | NP | R | P | P |
| Monocymbium ceresiiforme | TP | Dec | MP | R | P | P |
| Paspalum dilatatum | TP | EG | HP | R | P | C |
| Stiburus alopecuriodes | TP | CG | NP | C | P | P |
| Sporobolus africanum | TP | Inc ii | NP | R | R | P |
| Setaria sphacelata | TP | Dec | HP | P | P | R |
| Trichoneura grandiglumis | WP | Inc ii | NP | R | P | P |
| Tristachya leucotrix | TP | Dec | MP | C | P | R |
| Themeda triandra | TP | Dec | HP | C | P | C |
| Urochloa paniciodes | TP | Inc ii | NP | C | P | P |
| Soil Type | ||||
|---|---|---|---|---|
| Parameters | Hutton Soil | Avalon Soil | Clovelly Soil | SEM |
| Highly palatable (%) | 9.63 a | 4.83 b | 6.27 ab | 1.06 |
| Moderately palatable (%) | 3.83 b | 13.60 a | 5.06 b | 0.358 |
| Unpalatable (%) | 3.50 b | 6.87 ab | 7.70 a | 0.978 |
| Biomass (kg) | 5159.1 | 4110.7 | 5113.3 | 392.38 |
| Grazing capacity (Ha/LSU) | 1.6 | 2.0 | 1.6 | |
| CP | NDF | ADF | ADL | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grass Species | HS | AS | CS | HS | AS | CS | HS | AS | CS | HS | AS | CS |
| A. congesta | 82.9 b | 78.3 b | 77.1 b | 779.8 ab | 814.9 a | 763.9 ab | 714.9 Aa | 576.3 Ba | 461.3 Cab | 174.0 a | 220.4 a | 179.2 a |
| D. eriantha | 106.5 Aa | 94.6 Ba | 86.6 Ba | 688.2 c | 696.4 b | 684.7 c | 427.7 bc | 436.1 b | 392.4 b | 109.5 b | 116.9 c | 86.6 b |
| E. chloromelas | 67.1 c | 64.6 cd | 69.3 b | 715.7 c | 764.1 a | 732.1 bc | 400.2 Bc | 588.9 Aa | 467.7 Bab | 111.6 Bab | 189.3 Aab | 115.9 Bab |
| E. curvula | 50.0 Bd | 64.8 Acd | 70.2 Ab | 813.0 a | 810.2 a | 806.5 a | 444.5 bc | 461.6 b | 502.0 a | 129.4 ab | 91.3 c | 130.7 ab |
| E. gummiflua | 65.4 Ac | 57.2 Bd | 54.3 Bc | 774.0 abc | 781.8 a | 802.8 a | 362.9 Bc | 514.5 Aab | 525.4 Aa | 73.9 b | 103.7 c | 115.6 ab |
| E. plana | 80.0 Ab | 65.8 Bc | 52.9 Cc | 728.1 Bbc | 771.9 ABa | 801.6 Aa | 508.0 b | 448.6 b | 477.5 ab | 117.1 ab | 127.0 bc | 109.7 b |
| SEM | 2.7 | 20.6 | 34.3 | 22.3 | ||||||||
| DMD12 | DMD24 | DMD36 | DMD48 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grass Species | HS | AS | CS | HS | AS | CS | HS | AS | CS | HS | AS | CS |
| A. congesta | 67.4 a | 102.1 b | 121.1 b | 152.5 a | 175.5 a | 169.6 b | 194.7 b | 253.9 abc | 229.1 c | 238.5 a | 328.7 ab | 264.6 b |
| D. eriantha | 105.3 Ba | 185.7 Aba | 202.1 Aa | 205.1 a | 252.2 a | 327.6 b | 288.0 ab | 293.8 ab | 381.0 b | 320.4 a | 390.9 a | 409.4 b |
| E. chloromelas | 98.8 Ba | 176.0 Aa | 95.9 Bb | 158.5 Ca | 253.0 Ba | 537.0 Aa | 229.6 Cab | 334.5 Ba | 649.3 Aa | 289.8 Ba | 373.8 Bab | 623.6 Aa |
| E. curvula | 116.7 a | 85.2 b | 112.5 b | 164.5 a | 107.0 a | 211.4 b | 196.6 b | 168.4 bc | 256.3 bc | 245.1 a | 248.4 ab | 287.2 b |
| E. gammiflua | 122.7 a | 77.0 b | 112.8 b | 193.0 a | 140.6 a | 206.5 b | 331.5 Aa | 179.7 Bbc | 252.6 ABbc | 395.3 a | 231.0 ab | 281.9 b |
| E. plana | 105.0 a | 75.2 b | 130.7 b | 123.0 a | 124.1 a | 209.1 b | 182.0 b | 158.9 c | 289.1 bc | 241.7 a | 207.6 b | 364.6 b |
| SEM | 22.6 | 56.2 | 45.6 | 60.7 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokgakane, T.J.; Mlambo, V.; Ravhuhali, K.E.; Magoro, N. Contribution of Soil Type to Quantity and Nutritional Value of Grass Species on the South African Highveld. Resources 2021, 10, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/resources10100106
Mokgakane TJ, Mlambo V, Ravhuhali KE, Magoro N. Contribution of Soil Type to Quantity and Nutritional Value of Grass Species on the South African Highveld. Resources. 2021; 10(10):106. https://0-doi-org.brum.beds.ac.uk/10.3390/resources10100106
Chicago/Turabian StyleMokgakane, Thabile Joyce, Victor Mlambo, Khuliso Emmanuel Ravhuhali, and Norman Magoro. 2021. "Contribution of Soil Type to Quantity and Nutritional Value of Grass Species on the South African Highveld" Resources 10, no. 10: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/resources10100106