3.2. Hyperspectral and Broadband Indices
The graphs of
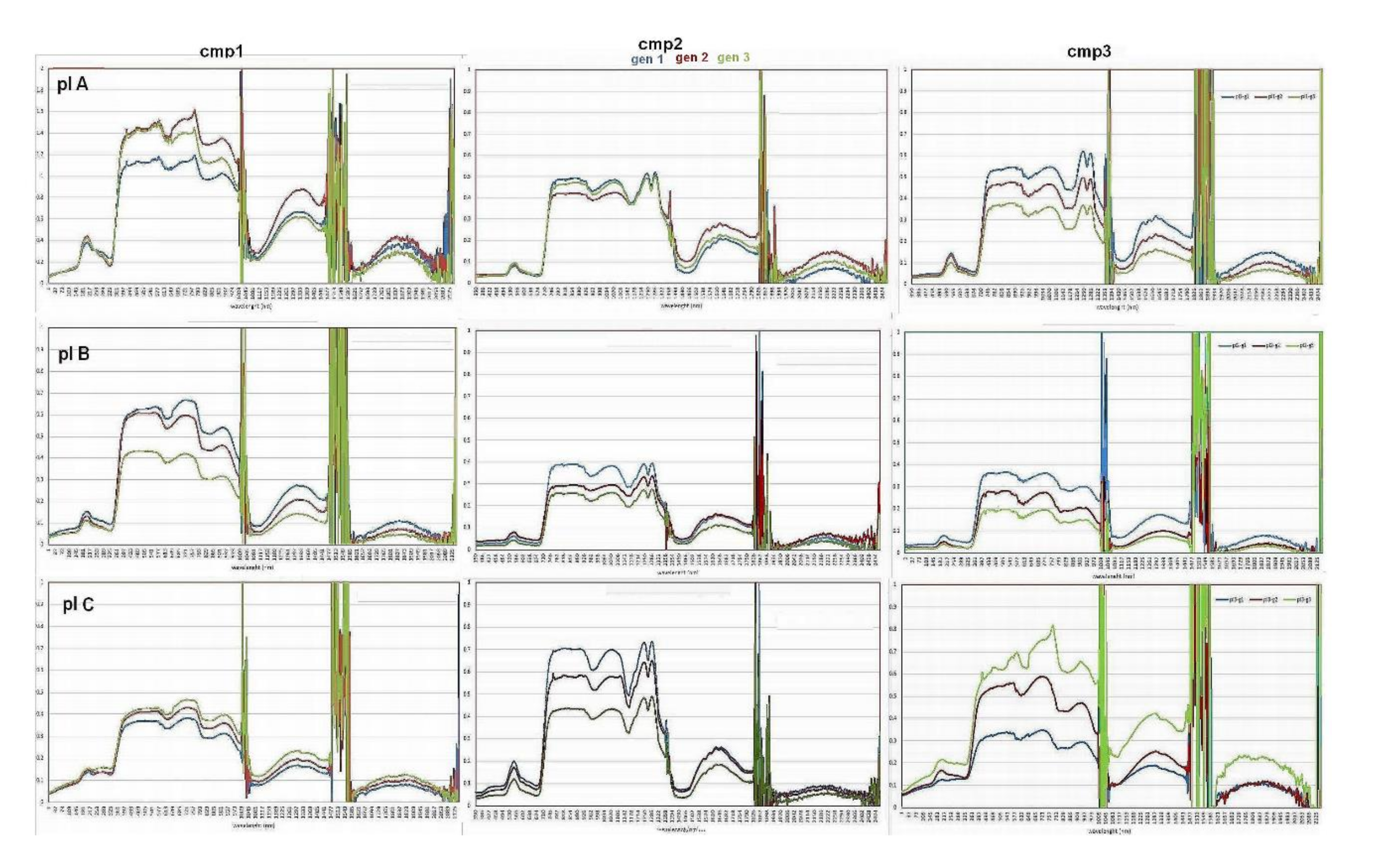
Figure 5 show the canopy hyperspectral signatures averaged for each of the three cardoon genotypes growing in the plateaus A, B, C, with rising water salinization concentrations, for all three campaigns. The similar graphs obtained from leaf spectral signatures were reported in the auxiliary materials (
Figure A2). The graphs along the column show the hyperspectral signatures of cardoon genotypes growing at different salinization degrees in the three campaigns. All graphs highlight the instabilities of the reflectance values recorded around the 1380 and 1890 nm wavelength ranges corresponding to air water vapor effects, with reflectance rapid variations exceeding the physical threshold (0, 1). In addition, as shown in the graph of the first campaign (cmp1), for plateau A (pl A), the effects due to the calibration fault led to unreliable reflectance values higher than 1. The spectral signatures of the different genotypes are sufficiently distinguishable in all the graphs, while those referred to plateaus A and B demonstrated a general decrease in spectral VIS responses and increase in NIR ones in the subsequent campaign, according to the increase in the chlorophyll absorption and leaves number of developing plantlets. The plants developed with the highest water salinization (pl C) showed instead an inverse distribution of the genotype hyperspectral signatures in the first (cmp1) and in the last (cmp3) measurement with an agreement on their trends of intermediate campaign (cmp2). In the first campaign, the development of most plants is in the early stages, and due to their low LAI values, the background reflectance contribution was significant. In the second campaign, the hyperspectral reflectance curves of the different plant genotypes stabilized and increased their differentiation in the third one.
Although most of the undesirable effects of noise above evidenced were corrected and mitigated in a pre-processing step, the exploitation of hyperspectral signatures under the form of normalized ratio spectral indices allowed us to further reduce this problem.
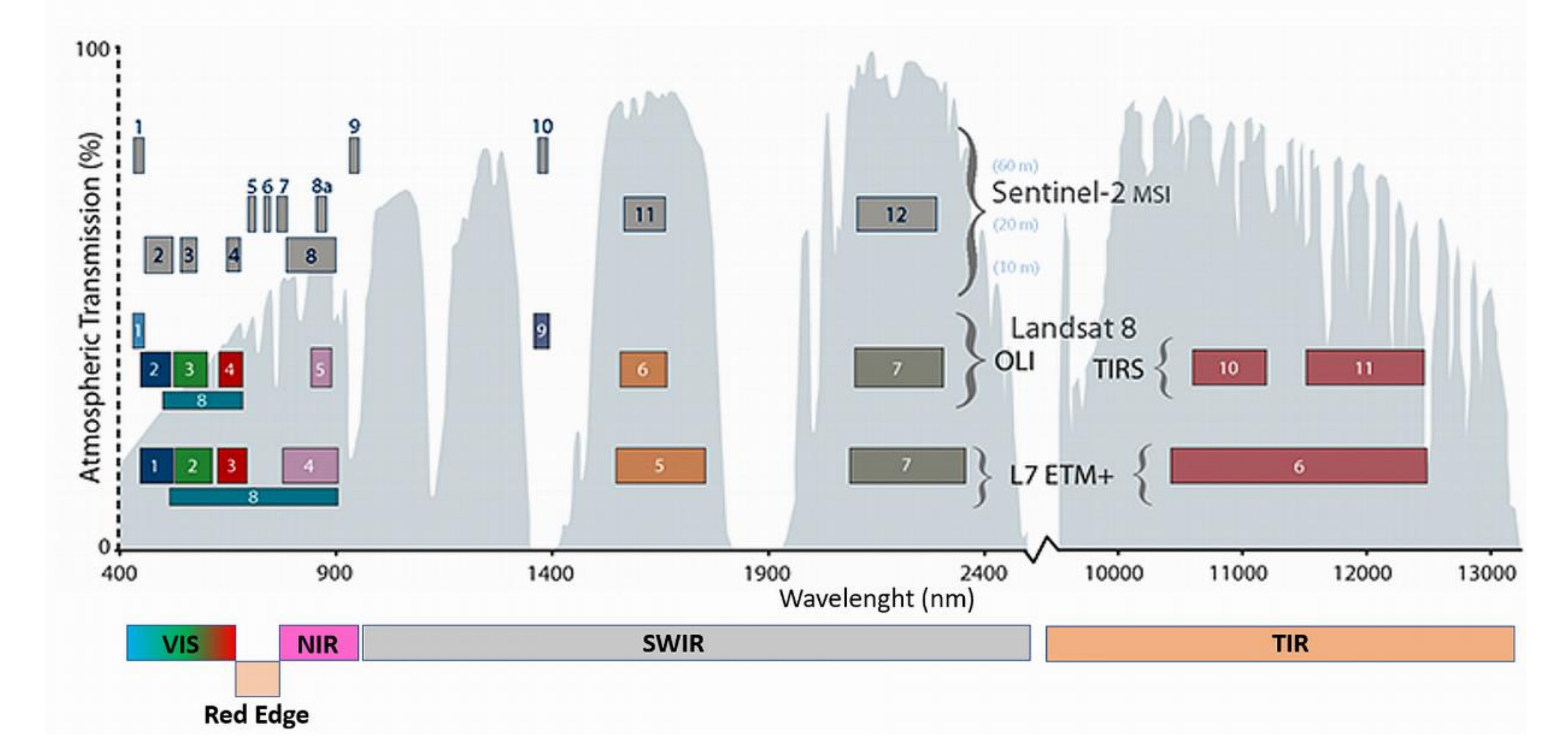
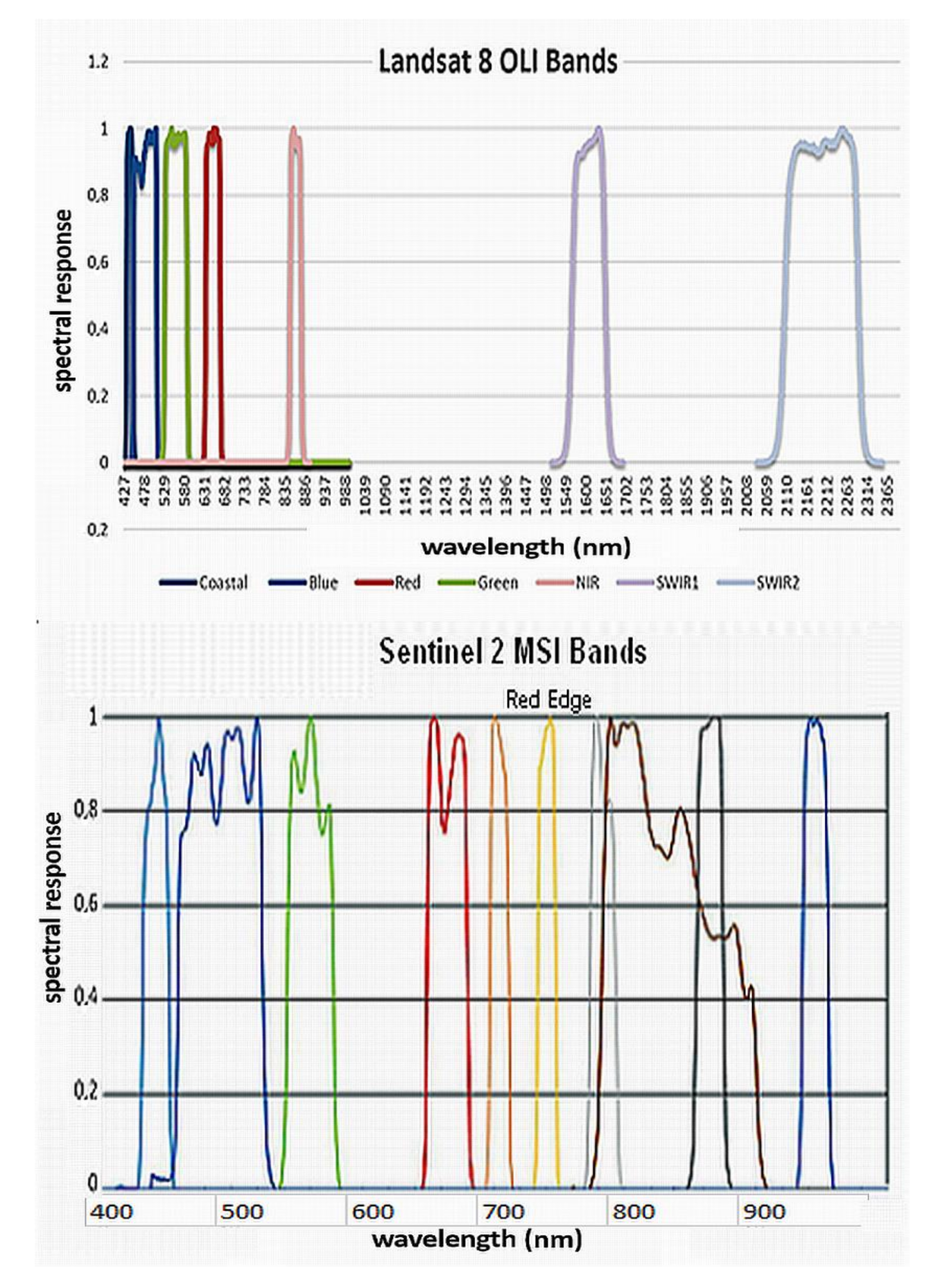
All the leaf and canopy (narrow band) indices introduced above were derived from the pre-processed hyperspectral data, then also the broadbands ones were assessed using the Landsat 8 OLI and Sentinel 2 MSI spectral bands’ filter functions (
Figure A1). The results obtained from the two-factor ANOVA analysis, through Tukey’s test, were summarized in
Table 4. For each of the selected spectral indices, the table includes the number of combination pairs referred to the plateau (plat. label) and genotype (gen. label) found significantly different (discriminated) in terms of means (
p-values < 0.05).
Table 5 and
Table 6 show instead the number of pairs discriminated by the broadband indices (at canopy level) assessed, respectively, for the L8 OLI and S2 MSI satellite sensors. The corresponding ANOVA detailed
p-value tables for all the indices were provided within the
Appendix A (
Table A4,
Table A5,
Table A6,
Table A7,
Table A8,
Table A9 and
Table A10). The number of discriminated combination pairs of each spectral index in the different measurement campaigns was then used as a proxy of its effectiveness in capturing the effects of the three water salinization levels on the three cardoon genotypes.
Globally, the canopy indices were found to perform better than those assessed by leaf measurements, especially in the most advanced development stages (cmp3), where only the NDVI, NPCI, SIPI RIRE, NDVI1, NDWI1 indices remain sensible to different salinization concentration at leaf level. The behavior of most leaf indices was significantly sensitive to the salinization level (plat label) in the first plant development stages (cmp1, cmp2) while once the plants were sufficiently grown, their capability to capture the effects of different water salinization levels (cmp3) decreases. In general, the leaf spectral indices capability to discriminate the spectral effects of the water salinization level on different genotype is poor, only the NDVI, TVI, SIPI, RIRE, NDVI1, NDWI1 and NDWI2 leaf indices were sensible to genotype effects at leaf level in the first growth stage (cmp1). Their capability to discriminate the effects due to the different cardoon genotypes becomes null (
Table 4 leaf reflectance side) in the subsequent development phases (i.e., cmp2, cmp3). Among the foliar spectral indices sensible only to the water salinization effects on cardoon, the most performant, in the 3rd development phase (cmp3) were neatly the RIRE and NDWI (able to discriminate, respectively, two and three of index combinations pairs), with the others at the same level of performance (
Table 6). Leaf indices seem mostly ineffective to capture the most subtle responses related to genotype differentiation in more advanced development stages coupled with a poorer effectiveness in detecting spectral effects of water salinization. In fact, at leaf level, only the RIRE was found capable of discriminating the spectral effects of all three levels of water salinization corresponding to three growing plots in the 3th measurement campaign (
Table 4).
Contrary to their leaf counterparts, the canopy spectral indices discrimination power of spectral effects linked, respectively, to water salinization level and genotype, increases with the development stage, according to the amplification of the spectral effects of the 3-d vegetation architecture growth compared to those of single leaves. In fact, all the models related to the selected canopy spectral indices became significant in the last development stage (cmp3, canopy reflectance side), with the effective ability of all indices to discriminate the water salinization effects, coupled with higher ability for 9 of them to capture also spectral effect differentiation related to the three cardoon genotypes (
Table 4, cmp3). The performance of these latter indices, evaluated for the cmp3 data by Tukey’s test, was reported in the canopy reflectance side of
Table 6. These achievements demonstrate that the OSAVI and TCARI indices were able to discriminate the spectral effects related to all water salinization levels, while SAVI, TCARI, and TVI were the most sensible to differentiations in the genotype spectra, with two pairs discriminated. Finally, the most effective index in discriminating the different levels of salinization of the water and the different cardoon genotypes was the TCARI, with its total score of five (three water salinization level + two genotype combination pairs discriminated). In the “total” column of
Table 4, the total sum of the combination pairs discriminated was reported as proxy of global performance of the index (for both genotype and plateau factors at leaf and canopy level). Between the structural indices, the SAVI scored the maximum number of discriminated pairs for both plateau and genotype factors, at leaf and canopy level, while TCARI resulted in the best performing of chlorophyll/pigment indices with a superior score number. Globally, also the red-edge indices have shown a good effectiveness with RE2 and REIP2 that maintained a significant performance in discriminating spectral effects of water salinization for different genotypes in the second measurement campaign. Despite its scarce capability to detect the spectral effects due to different genotypes, the NDWI of water spectral indices resulted in the best performing respect to all others. The global score, as number of discriminated pairs for both plateau and genotype combination pairs, at leaf and canopy levels, assessed for each spectral index ranges between 7 and 13, with many indices of the four groups gathering an intermediate value (9–10).
Most of L8 OLI structural indices have shown their effective capability to detect the spectral differences due to effects of different water salinization levels during all the plant growth phases (
Table 5). The other chlorophyll/pigment and water indices performed well only in the early and late stages. Their discrimination capability of spectral differences due to different genotypes is weak and limited to the advanced development stage. The SAVI
L, EVI
L, and NPCI
L were the best overall in terms of total number of discriminated pairs.
The Sentinel 2 MSI structural indices showed mostly a similar trend in detecting water salinization spectral effects (
Table 6) to which we added the useful capability of those based on Red-Edge Inflection Point (REIP) working also in intermediate and advanced stages of plantar development. The capability to detect the most subtle spectral differences in the three cardoon genotypes due to water salinization levels (gen) remains weak and limited to intermediate and advanced plant growth stages. The most performant indices, which reached a total score of six combination pairs discriminated, were SAVI
S, NPCI
S, NWI1
S, and the red-edge-based that demonstrated also the ability to detect genotype spectral differences both at the intermediate and advanced stages of plant development.
The different leaf and canopy hyperspectral indices through the data acquired during the three measurement campaigns evidenced a wide range of spectral indices sensibility, at plateau and genotype levels. In the early stage (cmp1), both the leaf and canopy indices demonstrated a useful capability to detect the plant spectral differences induced by the three water salinization levels, while only water coupled with some structural and pigment indices was found able to capture the effects at the genotype level. In the following stage (cmp2), contrary to canopy indices, the leaf ones maintained their aptitude to discriminate the effects of water salinization level without genotype distinction.
In the later stage (cmp3), the sensibility of the canopy indices, especially those of the structural and pigment ones including the genotype effects increased, while diminishing the reactivity of the leaf indices decreased. Similar trends were found for the S2 MSI broadband indices with additional capabilities demonstrated by those based on the red-edge, acquisition channels (
Figure 4). Both the OLI and S2 broadband indices demonstrated a suitable aptitude to detect the spectral effects of water salinization levels on cardoon plants, especially in early and sufficiently developed stages (1.2–1.4 stages of BBCH scale), with a weak additional sensitivity to different genotypic effects of water indices in these latter.
3.3. Biometry Modelling
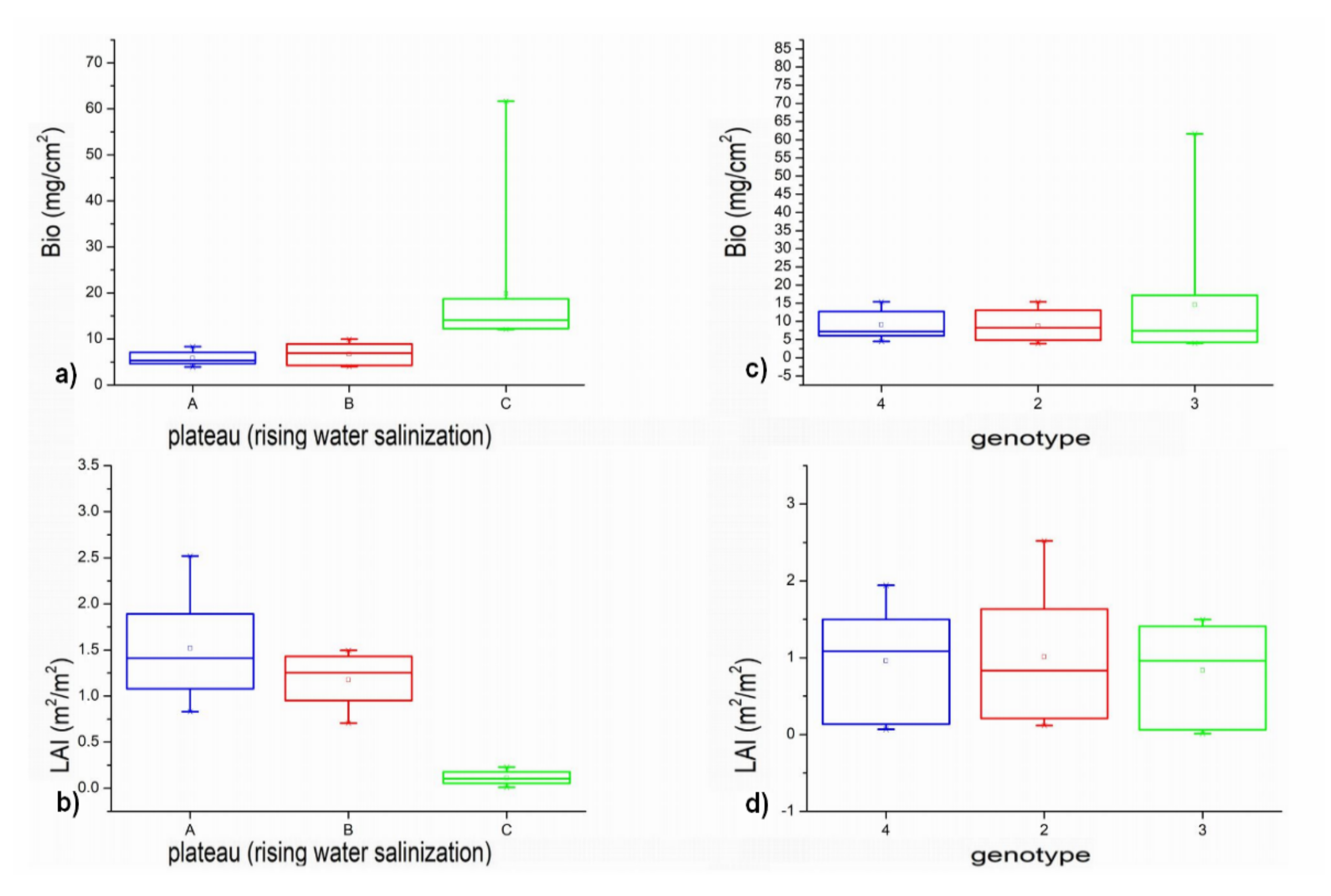
Figure 6 shows the boxplot graphs related to Biomass (mg/cm
2) and LAI (m
2/m
2) biometric measurements derived from cardoon samples acquired on field during the second campaign (cmp2). The graphs a and b show data related to the three plateaus with different water salinization levels (starting from A with lowest salinization to C with the maximum one). The graphs c and d show the boxplots assessed for the three different
C. cardunculus genotypes.
The average trend of LAI and biomass density (Bio) do not show significant (at 95% of confidence level) variations between plateaus A and B, with raising water salinization, while the differences get significant for samples of C plateau (
p-value < 0.05). This find is confirmed also by the results of Tukey’s test in
Table 7, where only the two pairs A-C and B-C of LAI/Bio mean values were discriminated at plateau level, while no significant variation was found for samples of different genotypes (
p-value > 0.05).
In agreement with these results, most of canopy indices of the second campaign show significant differences only at plateau level, while a weak discrimination of genotype effects was shown only by red-edge and water spectral indices (
Table 6, canopy reflectance, cmp2). Similar trends were found for the OLI and MSI broadband indices.
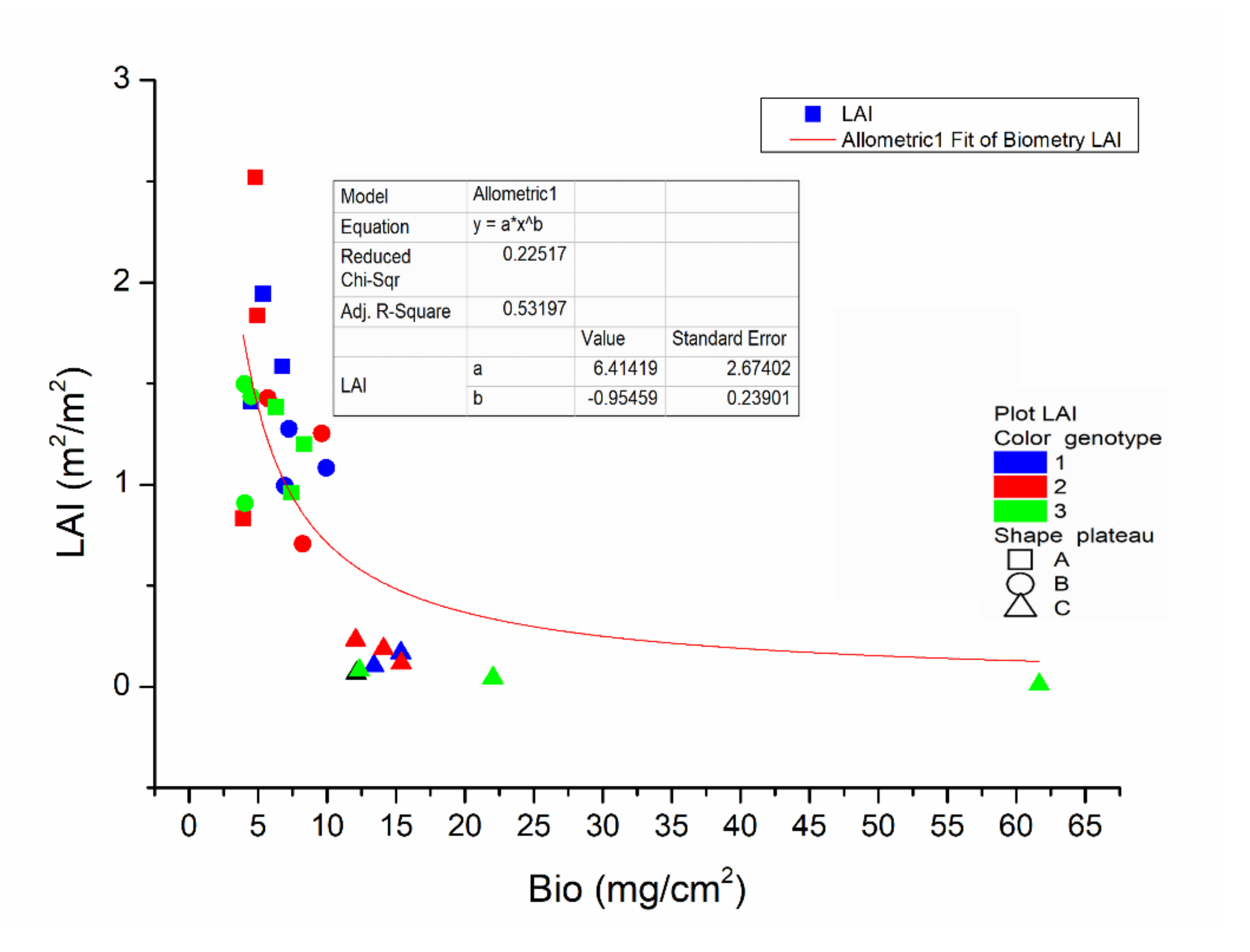
Figure 7 shows the plot of the biomass density (Bio) and the corresponding LAI measurements including their best-fit allometric curve as:
with the significant correlation found between them evidenced by an R
2adj (adjusted correlation coefficient) = 0.53197. The LAI shows a rising trend linked to that of the lowering Bio, depicted by the fitting allometric curve, with the best-fit intermediate LAI values around 1.25. The Bio variable measurements registered the maximum in case of the smallest leaf development caused by water salinization increase, in particular for the 3rd genotype whose values are the highest ones. The distribution of measurements from the samples of the C plateau (triangular dots) is mainly located in the area near the origin of the axes, in the lower left corner of the graph (average LAI = 0.112). The other data, referring to decreasing water salinization levels, spread along the best-fit curve, with those related to the B plateau (mean LAI = 1.175) and those from the A plateau above them (mean LAI = 1.518). From this graph and from the previous analyses of the biometric data, it appears that, above a water salinization threshold (i.e., water salt concentration of the plateau C), the global development of cardoon plants is significantly inhibited, while under this threshold, the water salinization neither affects the development of the leaf surface (LAI) nor the biomass density (Bio) of
C. cardunculus plants. In order to link the biometric data to the corresponding spectral indices for modelling purposes, both the average values at genotype and plateau levels have been assessed from the related measurement datasets. In particular, the leaf and canopy hyperspectral indices and the broadband ones of the second campaign have been selected to be extensively tested for linear modelling of the yield Y, derived from measured LAI and Bio biophysical parameters (Equation (1)). The results of this modelling approach were reported below, under the form of bi-dimensional scatter plots where the coordinates of each index are the R correlation (Spearman) coefficients derived from the linear model of Y (
y-axis) and LAI (
x-axis) parameters, more directly linked to biomass resources on field by the bioenergetic crops. In particular,
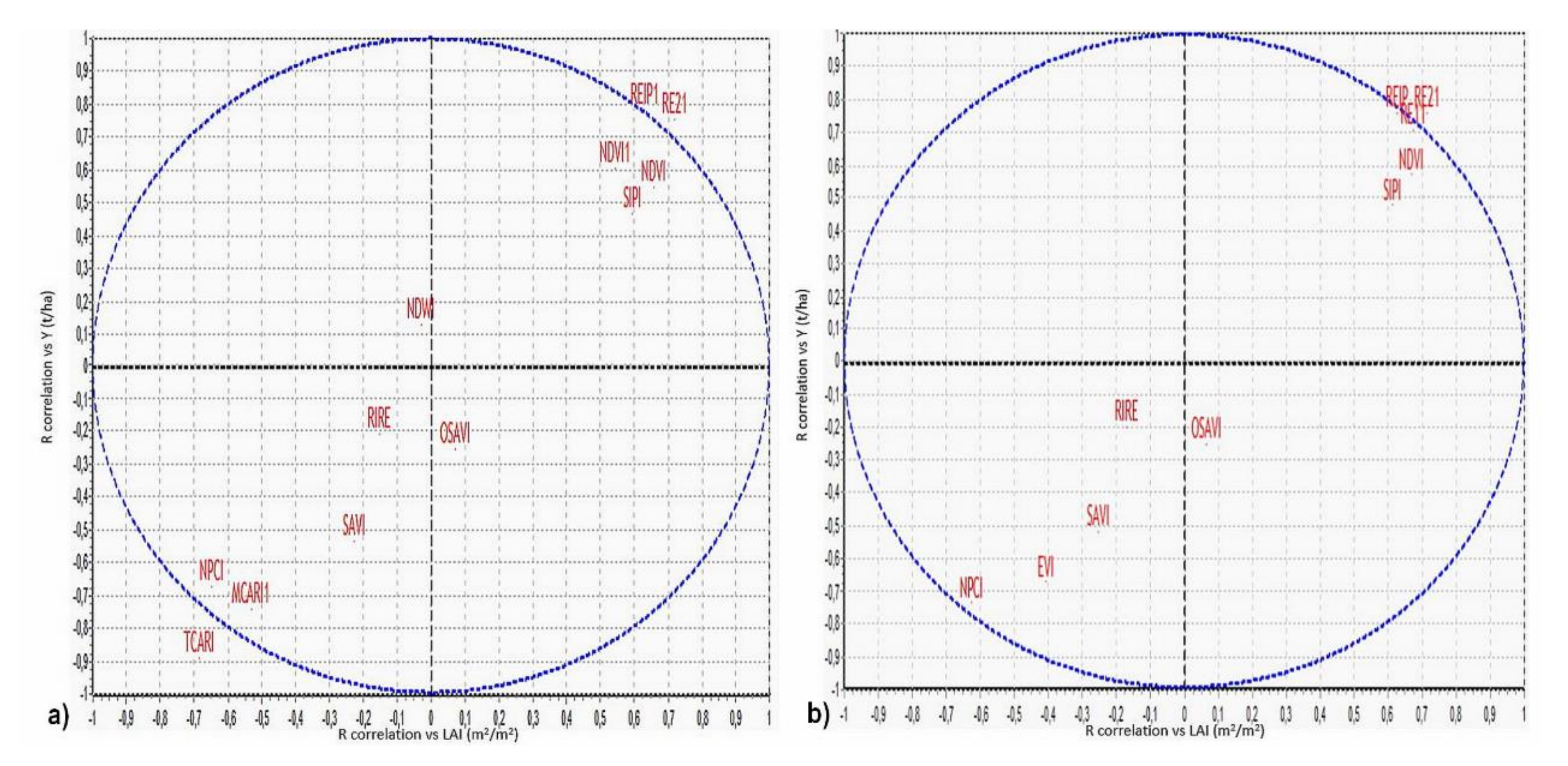
Figure 8a shows the scatter plots for canopy hyperspectral indices, while
Figure 8b includes that assessed for the Sentinel MSI broadband indices. In the graphs, some spectral indices that overlap each other are not reported to avoid confusion and improve the clearness of distributions. In general, in all the graphs, the position related to all indices is distributed along the bisector of the second and third quadrants, which means that the correlations of the spectral indices with LAI and Y may have the opposite sign depending also on the implicit contribution of Bio in Y without significant differences among the three genotypes.
As reported in graph a of
Figure 8, the correlation of the red-edge (REIP, RE21, RE11) canopy’s spectral indices, with the LAI/Y is positive and significant, with that slightly inferior of NDVI, NDVI1, and SIPI. The soil corrected SAVI and OSAVI, RIRE, and the NDWI indices showed instead poor correlation values with those of SAVI inferior of others and negative for both LAI and Y. The other indices (MCARI and NPCI) are characterized by a varying negative correlation with TCARI reaching the minimum values.
Most of the S2 broadband spectral indices (
Figure 8, graph b), presented a behavior similar to that of the corresponding hyperspectral canopy index, with those red-edge-based (RE11, RE12, REIP) more concentrated and having the maximum positive correlation with LAI and Y. Contrary to NDVI, the EVI index, specifically designed for HR satellite optical sensors (to minimize soil and atmospheric noise contribution), shows a negative correlation, probably due to the overcorrection of absent atmospheric effects, based on blue channel. The correlation of the OLI spectral indices showed a trend similar to that of the S2 MSI, with the NDVI achieving the maximum positive correlation but inferior to that of S2 red-edge ones. In general, from these preliminary R correlation test, the red-edge indices evaluated for the second field campaign (cmp2) happened most suitably in modelling the Y biometric data (
Figure 8a,b), maintaining at same time a discrimination capability for the corresponding effects of different water salinization levels (
Table 6, canopy reflectance indices, cmp2).
The most relevant positive correlation with Y and LAI is provided by the RE21 index, normalized difference red-edge index that reached the maximum value, followed by RE11 (
Figure 8). The biomass production assessment at field level requires spatial modeling of the yield Y, in order to provide input for sustainable management and bioenergy exploitation. In this perspective, a deeper analysis of this index was carried out considering that currently the hyperspectral and multispectral data collected remotely of the areas of interest for spatial distributions assessment can be provided by the airborne or satellite operating platforms (i.e., S2 MSI satellite).
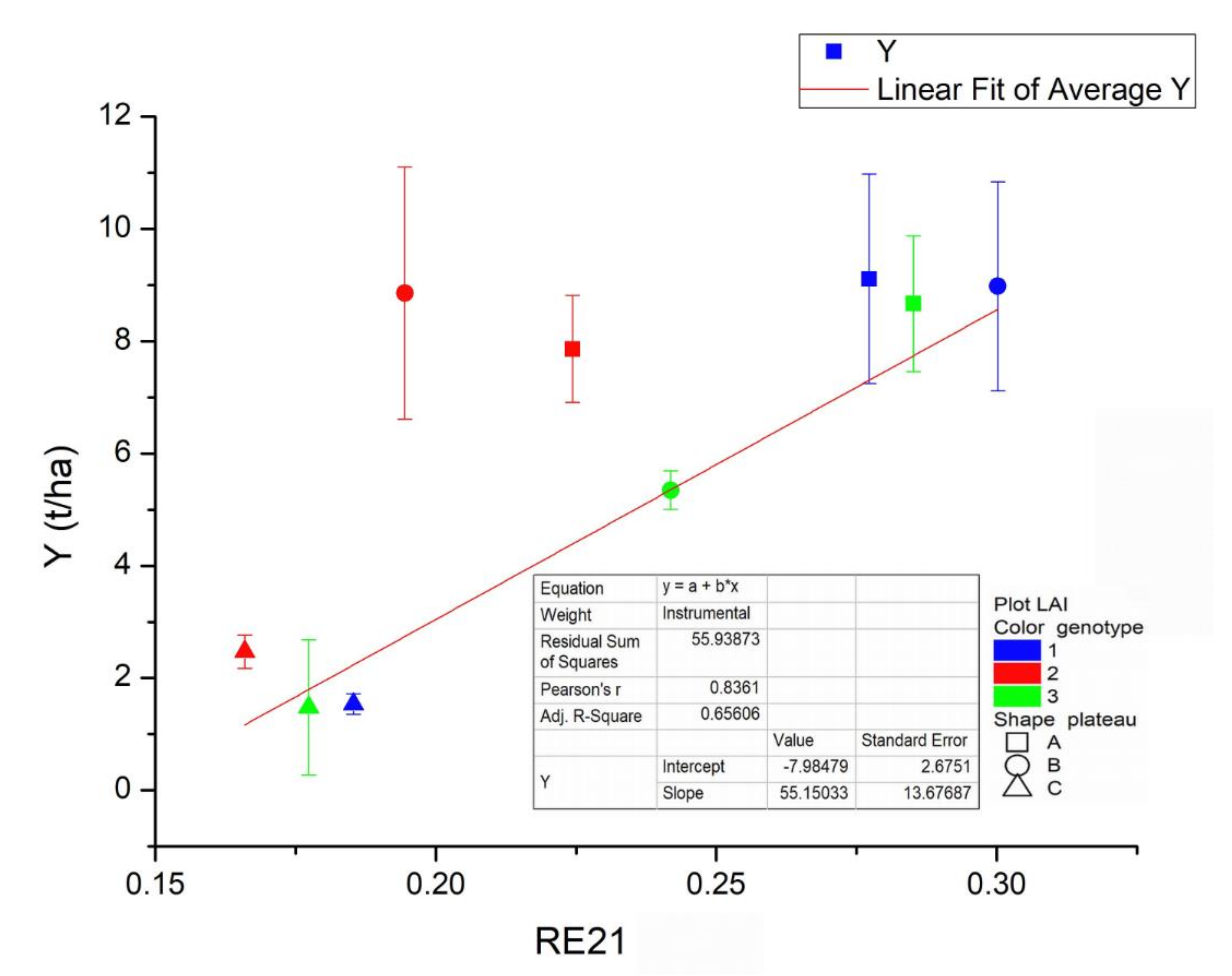
Figure 9 shows the average trends of the Y derived from the biometric data and the corresponding RE21 values for each plateau (different dot shape) and genotype (different dot color). The regression equation found is:
The R2 correlation of the related Y-RE21 linear model scored 0.836 (R2adj = 0.656) that means an effective capability to capture the above-ground biomass spatial variations supported by a model significance better than 95% (F-value = 16.26, Prob > F = 0.00498). The lowest Y (around 1.5 t/ha) and RE21 values were found for the cardoon plants of the three genotypes of the C plateau (triangular dots) fed with the most salinized water. The maximum (around 9 t/ha), instead, has been reached both by the first cardoon genotype grown in water at intermediate salinization (circle black dot) or not salinized water (square black dot) and by the second genotype (circle red dot) irrigated with water at intermediate salt concentration. The Y of plants of genotype 1 (black dots) and 2 (red dots), grown using water at intermediate salt concentration, approaches 9 t/ha (maximum); in particular, the Y of genotype 2 exceed that obtained with water non-salinized (about 8 t/ha).
The RE21 index and the corresponding Y values of all cardoon genotypes feeding with the most salinized water are rightly the lowest (triangular dots), but they differ in the trends of the remaining plateau. In fact, although the Y rising trend of the genotype 1 (black dots) is in agreement with water salinization reduction, the Y and RE21 index values corresponding to water middle salinization level (circle dot) were found too high respect to those of the genotype 3, while the genotype 2 showed a similar Y value compared to a significantly lower index. All the plateau averages of the genotype 3 (green dots) are rightly located along the fitting line, with Y related to middle water salinization significantly smaller than the other two and that obtained through the non-salinized water located between the others corresponding two. All the plateau Y averages of the second genotype (red dots) are located above and at a significant distance from the red line of best fit with the yield Y corresponding to the intermediate water salinization (red circle dot) higher than that of the plants fed with no salinized water (red square dot), which present an inversion not evidenced by the trends of the measurement of the other two genotypes.
A similar trend was found for the RE21
s index, derived from the S2 MSI broadband spectral responses, with a higher R
2 correlation coefficient and the remaining modelling parameters reported in
Table 8, including those stating the relevant model confidence level.
Although the ANOVA analysis of biometry data did not evidence significant differences of genotypes’ responses to three water salinization levels, the Y biomass growing trends of genotypes 1 and 3 were found in raw agreement each other and with water salinization decreasing, except for intermediate values, corresponding to different rises in Y and corresponding RE21 index. In particular, the Y and RE21 intermediate values of this genotype (black circle dot) approaches those referring to those obtained for no salinized water (black and green square dot). The plants of 2nd genotype (red dots) instead, after a more relevant growth of the Y related to intermediate water salinization (similar to that of genotype 1 but without a corresponding rise in RE21 index) show a decreased Y respect to the same genotype thistle grown with water not salinized. In addition, these plants presented a rise in corresponding RE21 (square red dot), equivalent to around one half respect to others without water salt concentration. These results show that, at irrigation water salt concentrations of 200 mM/L (plateau C), the growth of all thistle genotypes used here is significantly inhibited both in term of Y and spectral indices responses (Y = 1–2 t/ha), while at half concentrations (plateau B), the different genotypes exhibit different behaviors. In particular, the plants of genotype 1 and 2 (black and red circle dots) evidence productivity growth (Y = 9 t/ha) approaching those obtained for genotype 1 and 3 (black and green square dots), with water not salinized (plateau A). The Y values corresponding to the plateau A for genotypes 1 and 3 look quite similar, while that of the 2nd genotype diminishes respect the corresponding Y amount of plateau B (100 mM/L).
Ultimately, from these preliminary findings, the intermediate salt concentration around 100 mM/L seems to have a negative impact on the productivity Y of the cardoon genotype 3, not affecting that of the genotype 1 and favoring the development of the second genotype used here. Although the intermediate water salinization feeding involves a rise in RE21 spectral index responses of plants of all three genotypes, their growth amount differs. The plants of the cardoon genotype 2 exhibit the smallest index value, those of the cardoon genotype 3 presented intermediate RE21 values, while the highest were assessed for the genotype 1 plants. From the comparison with the data referred to samples irrigated without salinized water, we can evidence that the 100 mM/L concentration does not impact on the Y of cardoon genotype 1, while it improves productivity of the genotype 2 (of about 1 t/ha) and inhibits that of the genotype 3 (of about 3 t/ha), as can be derived from the differences in y-coordinates in the graphs. In any case, it should be highlighted that, although Y (t/ha) is specifically linked to the production of extensive field cultivation, its estimated maxima values in our controlled and limited experimental context (about 10 t/ha of above-ground biomass) are roughly compatible with those reported in the literature for bioenergy productions growing in the field at comparable agronomic condition and with low fertilizer inputs, taking into account also of biomass partitioning between leaves, stalks, and heads and wet/dry mass mutual dependence [
6,
9].

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










