Citrate-Coated Platinum Nanoparticles Exhibit a Primary Particle-Size Dependent Effect on Stimulating Melanogenesis in Human Melanocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Transmission Electron Microscopy (TEM)
2.3. Cell Culture
2.4. Cytotoxicity Assay
2.5. Melanogenesis Assay in MNT-1 Cells
2.6. Recovery Study of Melanogenesis by PtNPs in MNT-1 Cells
2.7. Intracellular Tyrosinase Activity in MNT-1 Cells
2.8. Intracellular Reactive Oxygen Species (ROS) in MNT-1 Cells
2.9. Melanogenesis Assay in HEMn-MP Cells
2.10. Dendricity Measurement in HEMn-MP Cells
2.11. Statistical Analysis
3. Results
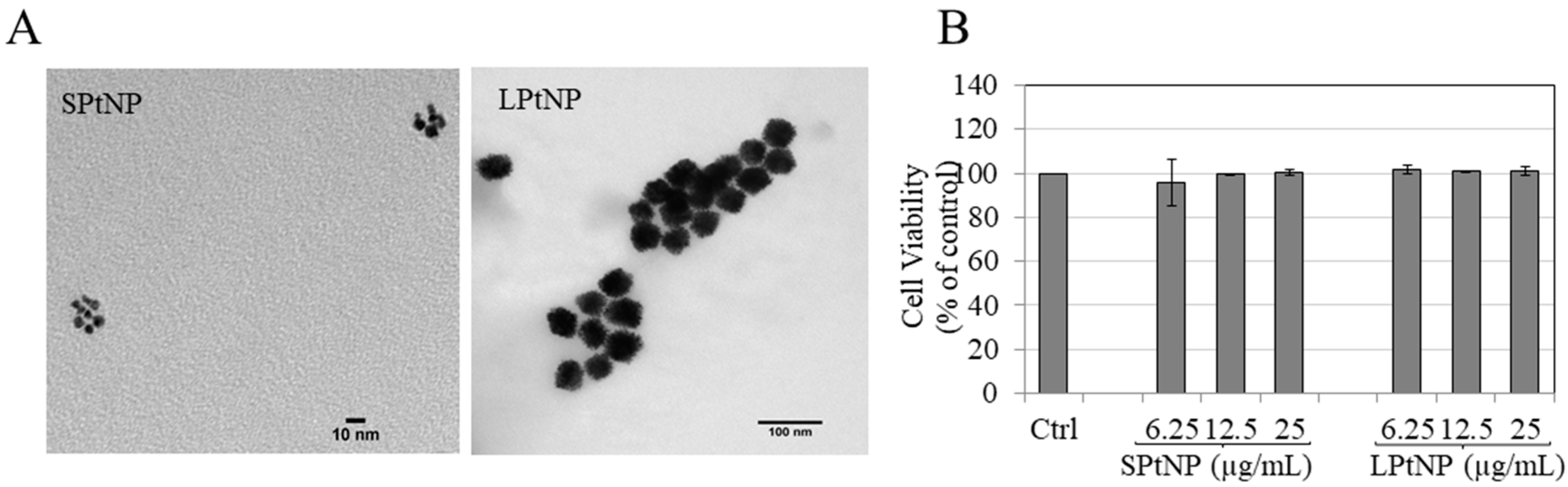
3.1. Characterization of PtNPs
3.2. Cellular Viability in MNT-1 Cells
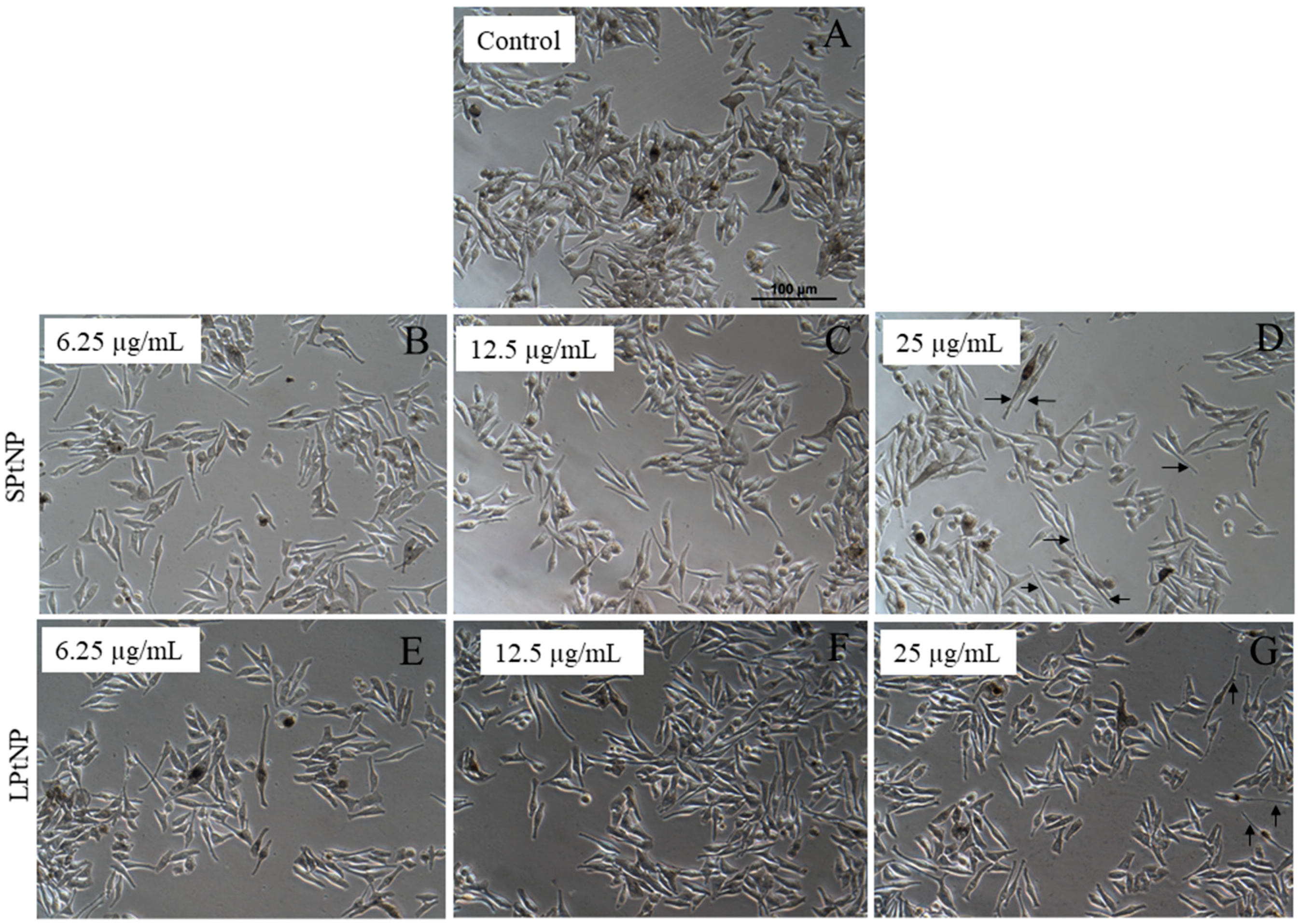
3.3. Cellular Morphology of MNT-1 Cells
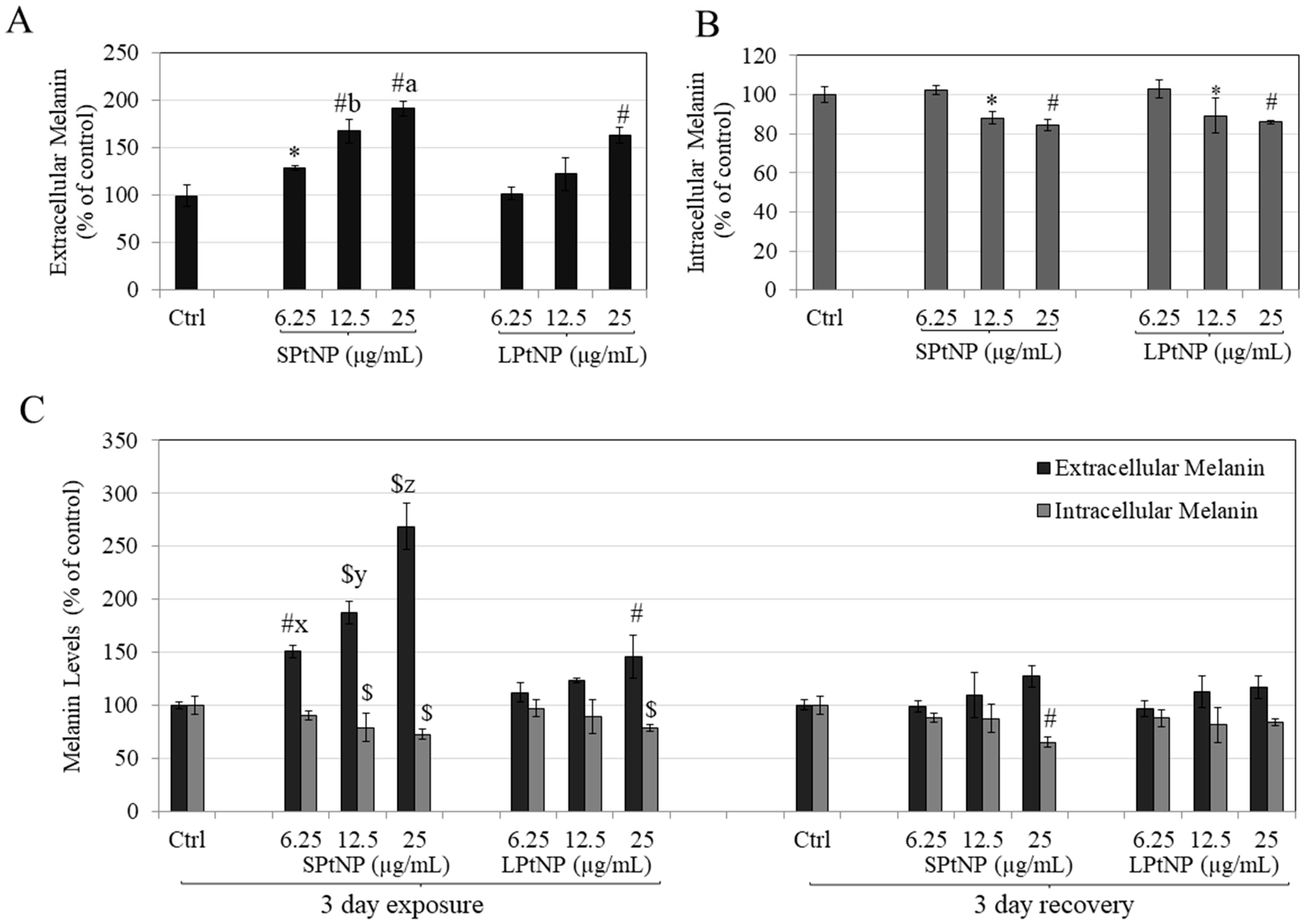
3.4. PtNPs Stimulated Extracellular Melanin in MNT-1 Human Melanoma Cells
3.5. Effects of PtNPs on Reversibility of Melanosome Export in MNT-1 Cells
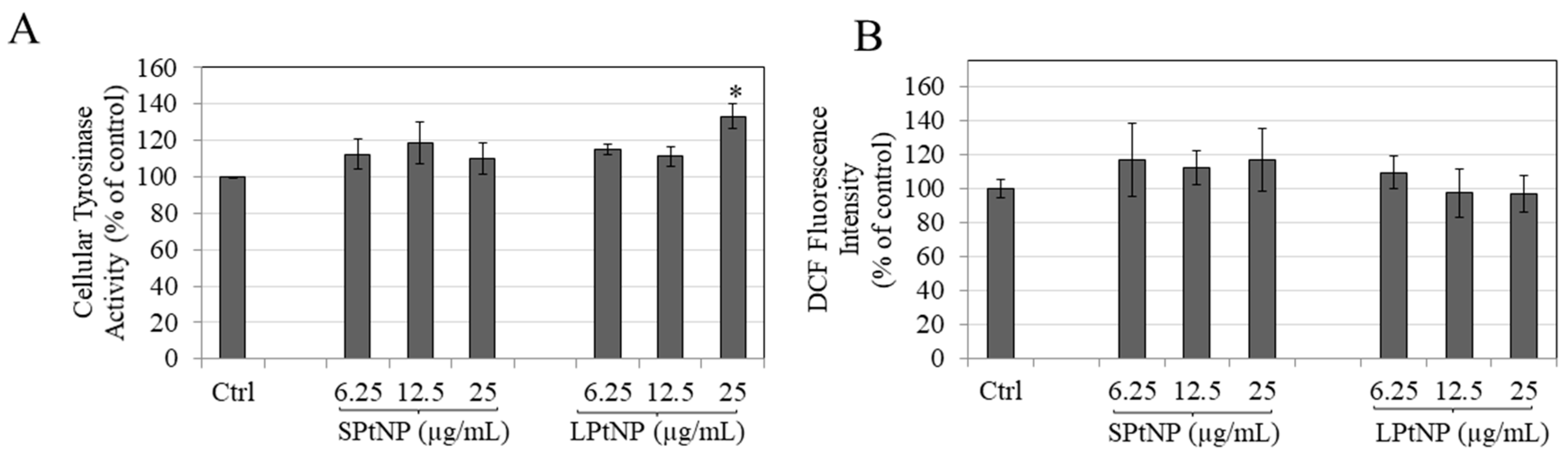
3.6. PtNPs Effects on Intracellular Tyrosinase Activity
3.7. Effects of PtNPs on Intracellular Reactive Oxygen Species (ROS) Levels in MNT-1 Cells
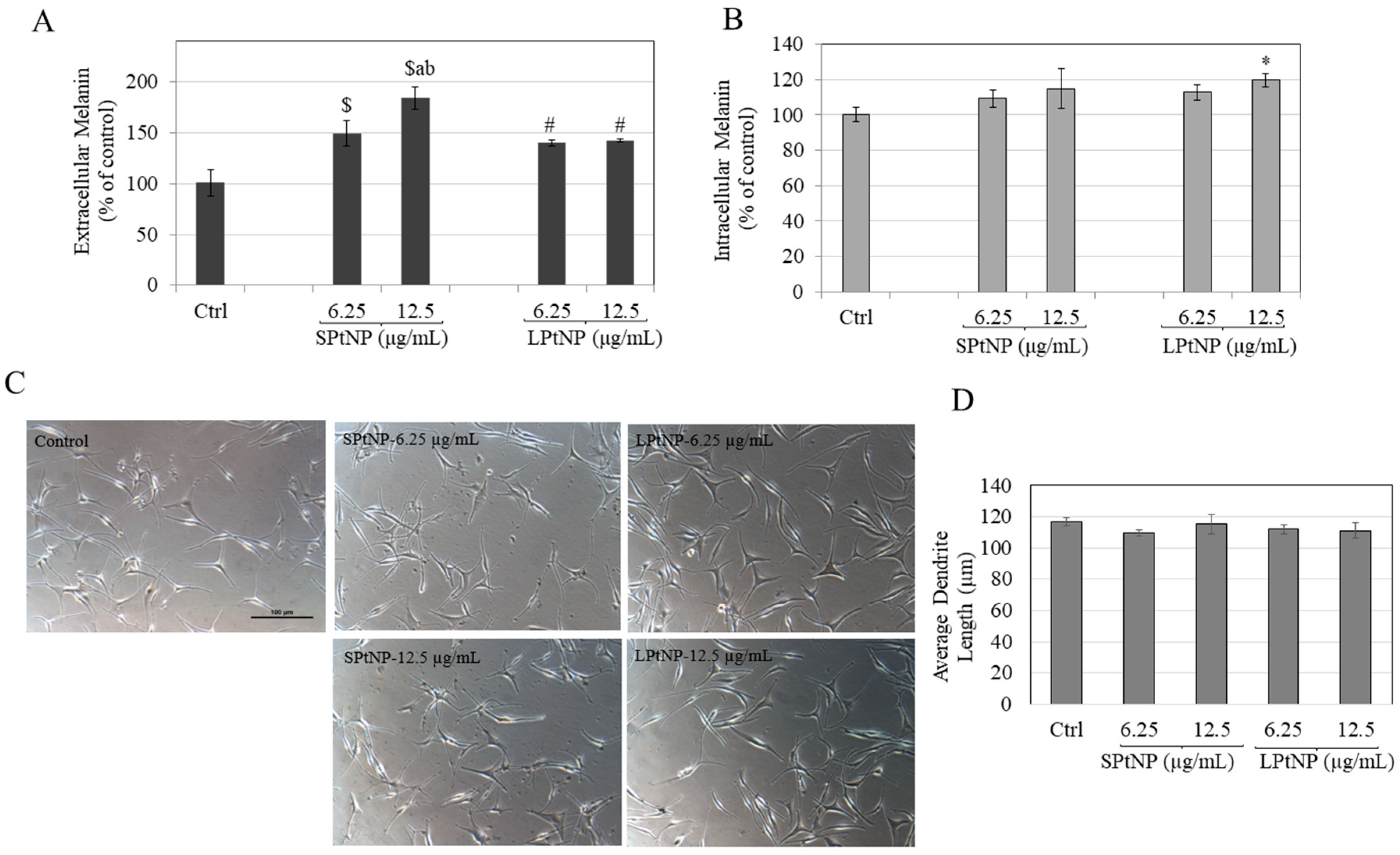
3.8. Effects of PtNPs in HEMn-MP Cells
3.9. PtNPs Did Not Alter Dendritic Morphology of HEMn-MP Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| L-DOPA | L-3,4 dihydroxyphenylalanine |
| DMEM | Dulbecco’s Modified Eagle Medium |
| HMGS | Human Melanocyte Growth Supplement |
| HEM | Human-Epidermal Melanocytes |
| MP | Moderately Pigmented |
| TEM | Transmission Electron Microscopy |
| PtNPs | Platinum Nanoparticles |
| SPtNP | Small Platinum Nanoparticles |
| LPtNP | Large Platinum Nanoparticles |
| ANOVA | Analysis of Variance |
| HBSS | Hank’s Balanced Salt Solution |
| ROS | Reactive Oxygen Species |
| DCF | Dichlorofluorescein |
References
- Fernandez-Flores, A.; Saeb-Lima, M.; Cassarino, D.S. Histopathology of Aging of The Hair Follicle. J. Cutan. Pathol. 2019, 46, 508–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and Physiological Implications of Melanic Pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, P.A. Melanogenesis and Melanoma. Pigment Cell Res. 2003, 16, 548–552. [Google Scholar] [CrossRef] [PubMed]
- D’orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. Uv Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hearing, V.J. Determination of Melanin Synthetic Pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, T.B.; Breathnach, A. The Epidermal Melanin Unit System. Dermatol. Wochenschr. 1963, 147, 481. [Google Scholar] [PubMed]
- Hirobe, T. Structure and Function of Melanocytes: Microscopic Morphology and Cell Biology of Mouse Melanocytes in The Epidermis and Hair Follicle. Histol. Histopathol. 1995, 10, 223–237. [Google Scholar] [PubMed]
- Dutta, S.; Panda, S.; Singh, P.; Tawde, S.; Mishra, M.; Andhale, V.; Athavale, A.; Keswani, S.M. Hypopigmentation in Burns Is Associated with Alterations in the Architecture of the Skin and the Dendricity of The Melanocytes. Burns 2020, 46, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes Are Transferred from Melanocytes to Keratinocytes Through the Processes of Packaging, Release, Uptake, and Dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Petersen, A.B.; Wulf, H.C.; Gniadecki, R.; Gajkowska, B. Dihydroxyacetone, The Active Browning Ingredient in Sunless Tanning Lotions, Induces Dna Damage, Cell-Cycle Block and Apoptosis in Cultured Hacat Keratinocytes. Mutat. Res. 2004, 560, 173–186. [Google Scholar] [CrossRef]
- Levine, J.A.; Sorace, M.; Spencer, J.; Siegel, D.M. The Indoor Uv Tanning Industry: A Review of Skin Cancer Risk, Health Benefit Claims, and Regulation. J. Am. Acad. Derm. 2005, 53, 1038–1044. [Google Scholar] [CrossRef]
- Pawelek, J.M. Approaches to Increasing Skin Melanin with Msh Analogs and Synthetic Melanins. Pigment Cell Res. 2001, 14, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.; Swope, V.B.; Suzuki, I.; Akcali, C.; Harriger, M.D.; Boyce, S.T.; Urabe, K.; Hearing, V.J. Mitogenic and Melanogenic Stimulation of Normal Human Melanocytes by Melanotropic Peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 1789–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fytianos, G.; Rahdar, A.; Kyzas, G.Z. Nanomaterials in Cosmetics: Recent Updates. Nanomaterials 2020, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Z.A.A.; Mohd-Nasir, H.; Ahmad, A.; Mohd Setapar, S.H.; Peng, W.L.; Chuo, S.C.; Khatoon, A.; Umar, K.; Yaqoob, A.A.; Mohamad Ibrahim, M.N. Role of Nanotechnology for Design and Development of Cosmeceutical: Application In Makeup and Skin Care. Front. Chem. 2019, 7, 739. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H. New Insights on Unique Features and Role of Nanostructured Materials in Cosmetics. Cosmetics 2020, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Rigano, L.; Lionetti, N. Nanobiomaterials In Galenic Formulations and Cosmetics. In Nanobiomaterials in Galenic Formulations and Cosmetics; Elsevier: Amsterdam, The Netherlands, 2016; pp. 121–148. [Google Scholar]
- Kockler, J.; Oelgemöller, M.; Robertson, S.; Glass, B.D. Influence of Titanium Dioxide Particle Size on the Photostability of the Chemical Uv-Filters Butyl Methoxy Dibenzoylmethane and Octocrylene in a Microemulsion. Cosmetics 2014, 1, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Haddada, M.B.; Gerometta, E.; Chawech, R.; Sorres, J.; Bialecki, A.; Pesnel, S.; Spadavecchia, J.; Morel, A.-L. Assessment of Antioxidant and Dermoprotective Activities of Gold Nanoparticles as Safe Cosmetic Ingredient. Colloids Surf. B Biointerfaces 2020, 189, 110855. [Google Scholar] [CrossRef]
- Gajbhiye, S.; Sakharwade, S. Silver Nanoparticles in Cosmetics. J. Cosmet. Dermatol. Sci. Appl. 2016, 6, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Foote, M.; Prow, T.W. Therapeutic Gold, Silver, and Platinum Nanoparticles. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 428–445. [Google Scholar] [CrossRef]
- Choosing Platinum Skin Care. Available online: https://bionyxskincare.com/choosing-platinum-skincare/ (accessed on 6 September 2020).
- Pedone, D.; Moglianetti, M.; De Luca, E.; Bardi, G.; Pompa, P.P. Platinum Nanoparticles in Nanobiomedicine. Chem. Soc. Rev. 2017, 46, 4951–4975. [Google Scholar] [CrossRef]
- Yoshihisa, Y.; Honda, A.; Zhao, Q.L.; Makino, T.; Abe, R.; Matsui, K.; Shimizu, H.; Miyamoto, Y.; Kondo, T.; Shimizu, T. Protective Effects of Platinum Nanoparticles Against Uv-Light-Induced Epidermal Inflammation. Exp. Dermatol. 2010, 19, 1000–1006. [Google Scholar] [CrossRef]
- Gatto, F.; Moglianetti, M.; Pompa, P.P.; Bardi, G. Platinum Nanoparticles Decrease Reactive Oxygen Species and Modulate Gene Expression without Alteration of Immune Responses in Thp-1 Monocytes. Nanomaterials 2018, 8, 392. [Google Scholar] [CrossRef] [Green Version]
- Kajita, M.; Hikosaka, K.; Iitsuka, M.; Kanayama, A.; Toshima, N.; Miyamoto, Y. Platinum Nanoparticle Is a Useful Scavenger of Superoxide Anion and Hydrogen Peroxide. Free Radic. Res. 2007, 41, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Moglianetti, M.; De Luca, E.; Pedone, D.; Marotta, R.; Catelani, T.; Sartori, B.; Amenitsch, H.; Retta, S.F.; Pompa, P.P. Platinum Nanozymes Recover Cellular Ros Homeostasis in an Oxidative Stress-Mediated Disease Model. Nanoscale 2016, 8, 3739–3752. [Google Scholar] [CrossRef]
- Yusof, F.; Ismail, N.A.S. Antioxidants Effects of Platinum Nanoparticles: A Potential Alternative Treatment to Lung Diseases. J. Appl. Pharm. Sci. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Katsumi, H.; Fukui, K.; Sato, K.; Maruyama, S.; Yamashita, S.; Mizumoto, E.; Kusamori, K.; Oyama, M.; Sano, M.; Sakane, T. Pharmacokinetics and Preventive Effects of Platinum Nanoparticles as Reactive Oxygen Species Scavengers on Hepatic Ischemia/Reperfusion Injury In Mice. Met. Integr. Biometal Sci. 2014, 6, 1050–1056. [Google Scholar] [CrossRef]
- Tuncer, S.; Colakoglu, M.; Ulusan, S.; Ertas, G.; Karasu, C.; Banerjee, S. Evaluation of Colloidal Platinum on Cytotoxicity, Oxidative Stress and Barrier Permeability Across the Gut Epithelium. Heliyon 2019, 5, E01336. [Google Scholar] [CrossRef] [Green Version]
- Depciuch, J.; Stec, M.; Klebowski, B.; Maximenko, A.; Drzymala, E.; Baran, J.; Parlinska-Wojtan, M. Size Effect of Platinum Nanoparticles in Simulated Anticancer Photothermal Therapy. Photodiagn. Photodyn. 2020, 29, 101594. [Google Scholar] [CrossRef]
- Buchtelova, H.; Dostalova, S.; Michalek, P.; Krizkova, S.; Strmiska, V.; Kopel, P.; Hynek, D.; Richtera, L.; Ridoskova, A.; Adam, P.; et al. Size-Related Cytotoxicological Aspects of Polyvinylpyrrolidone-Capped Platinum Nanoparticles. Food Chem. Toxicol. 2017, 105, 337–346. [Google Scholar] [CrossRef]
- Konieczny, P.; Goralczyk, A.G.; Szmyd, R.; Skalniak, L.; Koziel, J.; Filon, F.L.; Crosera, M.; Cierniak, A.; Zuba-Surma, E.K.; Borowczyk, J. Effects Triggered by Platinum Nanoparticles on Primary Keratinocytes. Int. J. Nanomed. 2013, 8, 3963. [Google Scholar]
- Kim, K.J.; Chang, S.E.; Choi, J.H.; Sung, K.J.; Moon, K.C.; Koh, J.K. Periungal Hyperpigmentation Induced by Cisplatin. Clin. Exp. Dermatol. 2002, 27, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Noori, M.; Hunter-Ellul, L.; Kelly, B. Serpentine Supravenous Hyperpigmentation Following Cisplatin and Pemetrexed Chemotherapy. Cutis 2017, 99, E20–E22. [Google Scholar]
- Al-Lamki, Z.; Pearson, P.; Jaffe, N. Localized Cisplatin Hyperpigmentation Induced by Pressure. A Case Report. Cancer 1996, 77, 1578–1581. [Google Scholar] [CrossRef]
- Shibuya, S.; Ozawa, Y.; Watanabe, K.; Izuo, N.; Toda, T.; Yokote, K.; Shimizu, T. Palladium and Platinum Nanoparticles Attenuate Aging-Like Skin Atrophy Via Antioxidant Activity in Mice. PLoS ONE 2014, 9, E109288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Yoshikawa, R.; Ichihashi, M. The Novel Therapy for Vitiligo Vulgaris: Topical Use of Cosmetic Cream of Platinum Nanoparticles and Palladium Nanoparticles Which Show Strong Catalase-Like Activity. J. Pigment. Disord. 2015, 2. [Google Scholar] [CrossRef]
- Tsuji, G.; Hashimoto-Hachiya, A.; Takemura, M.; Kanemaru, T.; Ichihashi, M.; Furue, M. Palladium and Platinum Nanoparticles Activate Ahr and Nrf2 In Human Keratinocytes-Implications In Vitiligo Therapy. J. Investig. Dermatol. 2017, 137, 1582–1586. [Google Scholar] [CrossRef] [Green Version]
- Takabe, W.; Yagi, M.; Ichihashi, M.; Yonei, Y. Anti-Glycative Effect of Palladium and Platinum Nanoparticle Solution. Glycative Stress Res. 2016, 3, 222–228. [Google Scholar]
- Liu, Y.; Wu, H.; Chong, Y.; Wamer, W.G.; Xia, Q.; Cai, L.; Nie, Z.; Fu, P.P.; Yin, J.-J. Platinum Nanoparticles: Efficient and Stable Catechol Oxidase Mimetics. ACS Appl. Mater. Interfaces 2015, 7, 19709–19717. [Google Scholar] [CrossRef]
- Sirimahachaiyakul, P.; Sood, R.F.; Muffley, L.A.; Seaton, M.; Lin, C.-T.; Qiao, L.; Armaly, J.S.; Hocking, A.M.; Gibran, N.S. Race Does Not Predict Melanocyte Heterogeneous Responses To Dermal Fibroblast-Derived Mediators. PLoS ONE 2015, 10, E0139135. [Google Scholar] [CrossRef]
- Denat, L.; Kadekaro, A.L.; Marrot, L.; Leachman, S.A.; Abdel-Malek, Z.A. Melanocytes As Instigators and Victims of Oxidative Stress. J. Investig. Dermatol. 2014, 134, 1512–1518. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Kajita, M.; Kim, J.; Kanayama, A.; Takahashi, K.; Mashino, T.; Miyamoto, Y. In Vitro Free Radical Scavenging Activity of Platinum Nanoparticles. Nanotechnology 2009, 20, 455105. [Google Scholar] [CrossRef]
- Asharani, P.V.; Xinyi, N.; Hande, M.P.; Valiyaveettil, S. Dna Damage and P53-Mediated Growth Arrest In Human Cells Treated With Platinum Nanoparticles. Nanomedicine 2010, 5, 51–64. [Google Scholar] [CrossRef]
- Shoshan, M.S.; Vonderach, T.; Hattendorf, B.; Wennemers, H. Peptide-Coated Platinum Nanoparticles With Selective Toxicity Against Liver Cancer Cells. Angew. Chem. Int. Ed. Engl. 2019, 58, 4901–4905. [Google Scholar] [CrossRef]
- Monopoli, M.P.; Aberg, C.; Salvati, A.; Dawson, K.A. Biomolecular Coronas Provide the Biological Identity of Nanosized Materials. Nat. Nanotechnol. 2012, 7, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Juling, S.; Niedzwiecka, A.; Bóhmert, L.; Lichtenstein, D.; Selve, S.R.; Braeuning, A.; Thünemann, A.F.; Krause, E.; Lampen, A. Protein Corona Analysis of Silver Nanoparticles Links to Their Cellular Effects. J. Proteome Res. 2017, 16, 4020–4034. [Google Scholar] [CrossRef]
- Lundqvist, M.; Stigler, J.; Elia, G.; Lynch, I.; Cedervall, T.; Dawson, K.A. Nanoparticle Size and Surface Properties Determine the Protein Corona with Possible Implications For Biological Impacts. Proc. Natl. Acad. Sci. USA 2008, 105, 14265–14270. [Google Scholar] [CrossRef] [Green Version]
- Tenzer, S.; Docter, D.; Rosfa, S.; Wlodarski, A.; Kuharev, J.R.; Rekik, A.; Knauer, S.K.; Bantz, C.; Nawroth, T.; Bier, C. Nanoparticle Size Is A Critical Physicochemical Determinant of the Human Blood Plasma Corona: A Comprehensive Quantitative Proteomic Analysis. ACS Nano 2011, 5, 7155–7167. [Google Scholar] [CrossRef]
- Bhatnagar, V.; Srirangam, A.; Abburi, R. In Vitro Modulation of Proliferation and Melanization of Melanoma Cells by Citrate. Mol. Cell. Biochem. 1998, 187, 57–65. [Google Scholar] [CrossRef]
- Lin, C.-X.; Gu, J.-L.; Cao, J.-M. The Acute Toxic Effects of Platinum Nanoparticles on Ion Channels, Transmembrane Potentials of Cardiomyocytes In Vitro and Heart Rhythm In Vivo In Mice. Int. J. Nanomed. 2019, 14, 5595. [Google Scholar] [CrossRef] [Green Version]
- Labrador-Rached, C.J.; Browning, R.T.; Braydich-Stolle, L.K.; Comfort, K.K. Toxicological Implications of Platinum Nanoparticle Exposure: Stimulation of Intracellular Stress, Inflammatory Response, and Akt Signaling In Vitro. J. Toxicol. 2018, 2018, 1367801. [Google Scholar] [CrossRef] [Green Version]
- Gollwitzer, C.; Bartczak, D.; Goenaga-Infante, H.; Kestens, V.; Krumrey, M.; Minelli, C.; Pálmai, M.; Ramaye, Y.; Roebben, G.; Sikora, A. A Comparison of Techniques for Size Measurement of Nanoparticles In Cell Culture Medium. Anal. Methods 2016, 8, 5272–5282. [Google Scholar] [CrossRef] [Green Version]
- Marucco, A.; Aldieri, E.; Leinardi, R.; Bergamaschi, E.; Riganti, C.; Fenoglio, I. Applicability and Limitations in the Characterization of Poly-Dispersed Engineered Nanomaterials In Cell Media By Dynamic Light Scattering (Dls). Materials 2019, 12, 3833. [Google Scholar] [CrossRef] [Green Version]
- Demir, V.; Bucher, J.; Kropf, C.; Arenz, M.; Segner, H. Comparative Study of Cytotoxicity By Platinum Nanoparticles and Ions In Vitro Systems Based on Fish Cell Lines. Toxicol. In Vitro 2020, 66, 104859. [Google Scholar] [CrossRef]
- Gehrke, H.; Pelka, J.; Hartinger, C.G.; Blank, H.; Bleimund, F.; Schneider, R.; Gerthsen, D.; Bräse, S.; Crone, M.; Türk, M. Platinum Nanoparticles and Their Cellular Uptake and Dna Platination At Non-Cytotoxic Concentrations. Arch. Toxicol. 2011, 85, 799–812. [Google Scholar] [CrossRef]
- Seiberg, M. Keratinocyte–Melanocyte Interactions During Melanosome Transfer. Pigment Cell Res. 2001, 14, 236–242. [Google Scholar] [CrossRef]
- Delevoye, C. Melanin Transfer: The Keratinocytes Are More Than Gluttons. J. Investig. Dermatol. 2014, 134, 877–879. [Google Scholar] [CrossRef] [Green Version]
- Hirobe, T. Keratinocytes Regulate the Function of Melanocytes. Dermatol. Sin. 2014, 32, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.N.; Marfatia, R.K.; Saikia, S.S. A Study of Noncultured Extracted Hair Follicle Outer Root Sheath Cell Suspension for Transplantation In Vitiligo. Int. J. Trichology 2016, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J.; Paus, R. Graying: Gerontobiology of the Hair Follicle Pigmentary Unit. Exp. Gerontol. 2001, 36, 29–54. [Google Scholar] [CrossRef]
- Campbell, K.I.; George, E.L.; Hall, L.L.; Stara, J.F. Dermal Irritancy of Metal Compounds. Studies with Palladium, Platinum, Lead, and Manganese Compounds. Arch. Environ. Health 1975, 30, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Czubacka, E.; Czerczak, S. Are Platinum Nanoparticles Safe To Human Health? Med. Pr. 2019, 70, 487–495. [Google Scholar] [CrossRef]
- Shibuya, S.; Watanabe, K.; Tsuji, G.; Ichihashi, M.; Shimizu, T. Platinum and Palladium Nanoparticle-Containing Mixture, Paplal, Does Not Induce Palladium Allergy. Exp. Dermatol. 2019, 28, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Aygun, A.; Gülbagca, F.; Ozer, L.Y.; Ustaoglu, B.; Altunoglu, Y.C.; Baloglu, M.C.; Atalar, M.N.; Alma, M.H.; Sen, F. Biogenic Platinum Nanoparticles Using Black Cumin Seed and Their Potential Usage as Antimicrobial and Anticancer Agent. J. Pharm. Biomed. Anal. 2020, 179, 112961. [Google Scholar] [CrossRef]
- Naseer, A.; Ali, A.; Ali, S.; Mahmood, A.; Kusuma, H.; Nazir, A.; Yaseen, M.; Khan, M.; Ghaffar, A.; Abbas, M. Biogenic and Eco-Benign Synthesis of Platinum Nanoparticles (Pt Nps) Using Plants Aqueous Extracts and Biological Derivatives: Environmental, Biological and Catalytic Applications. J. Mater. Res. Technol. 2020, 9, 9093–9107. [Google Scholar] [CrossRef]
- Joshi, P.G.; Nair, N.; Begum, G.; Joshi, N.B.; Sinkar, V.P.; Vora, S. Melanocyte-keratinocyte interaction induces calcium signalling and melanin transfer to keratinocytes. Pigment Cell Res. 2007, 20, 380–384. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goenka, S.; Toussaint, J. Citrate-Coated Platinum Nanoparticles Exhibit a Primary Particle-Size Dependent Effect on Stimulating Melanogenesis in Human Melanocytes. Cosmetics 2020, 7, 88. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040088
Goenka S, Toussaint J. Citrate-Coated Platinum Nanoparticles Exhibit a Primary Particle-Size Dependent Effect on Stimulating Melanogenesis in Human Melanocytes. Cosmetics. 2020; 7(4):88. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040088
Chicago/Turabian StyleGoenka, Shilpi, and Jimmy Toussaint. 2020. "Citrate-Coated Platinum Nanoparticles Exhibit a Primary Particle-Size Dependent Effect on Stimulating Melanogenesis in Human Melanocytes" Cosmetics 7, no. 4: 88. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040088