Melanogenesis Effect of 7-acetoxy-4-methylcoumarin in B16F10 Melanoma Cells



1
Department of Pharmaceutical Engineering & Biotechnology, Sunmoon University, Chungnam 31460, Korea
2
Microorganism Resources Division, Biological Resources Research Department, National Institute of Biological Resource, Incheon 22689, Korea
*
Author to whom correspondence should be addressed.
Cosmetics 2020, 7(4), 94; https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040094
Submission received: 16 November 2020
/
Revised: 30 November 2020
/
Accepted: 1 December 2020
/
Published: 2 December 2020
(This article belongs to the Special Issue Aesthetic and Cosmetic Dermatology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The increased interest in anti-whitening dyes has enhanced the research interest to identify efficient melanogenic activators. Melanogenesis is the process of melanin production by melanocytes in the hair follicles and skin, which is mediated by several enzymes, such as microphthalmia-associated transcription factor (MITF), tyrosinase (TYR), tyrosinase-related protein (TRP)-1, and TRP-2. This study investigated the melanogenesis-stimulating effect of 4-Methylumbelliferone (4MUMB) and its synthetic derivatives, 7-acetoxy-4-methylcoumarin (7A4MC) and 4-methylheriniarin (4MH) in B16F10 melanoma cells. The cytotoxicity of these compounds was investigated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, followed by the assessment of the melanin content and the intracellular TYR activity. Finally, the expression levels of the key enzymes involved in melanogenesis were investigated. 7A4MC increased melanin production in B16F10 cells relative to that by 4MUMB and 4MH treated cells in a dose-dependent manner without significant cytotoxicity. Concomitantly, 7A4MC significantly increased TYR activity and enhanced the expression of MITF, which significantly induced the expression of TRP-1, TRP-2, and TYR. Furthermore, 7A4MC stimulated melanogenesis via increased phosphorylation of c-Jun N-terminal kinases (JNK) and reduced phosphorylation of protein kinase B (AKT). These results confirmed the melanogenesis-inducing effects of 7A4MC and indicated its potential use as an anti-hair bleaching agent in cosmetics industries.
1. Introduction
Melanin, a black/brown skin pigment produced by melanocytes in the skin through the complex process of melanogenesis, determines the color of the eyes and hair and also protects the skin against the damage induced by UV radiation (UVR) [1,2]. The increase in the common use of anti-whitening hair dyes in cosmetic industries has increased the interest in identifying novel sources of melanogenesis activators [3]. UV radiation (UVR) is absorbed by different cellular chromophores present in the skin, such as melanin, proteins, amino acids, RNA, DNA, and other biomolecules. Excessive exposure of UVR induces several photochemical reactions involving reactive oxygen species (ROS), consequently leading to skin damage [1]. In particular, UVR can trigger an inflammatory response through various mechanisms, such as the direct activation of keratinocytes and other cells that release inflammatory mediators by activating components of the skin’s immune system and the redistribution of autoantigens isolated from cells damaged by UVR [2]. UV-induced DNA damage activates the binding of the transcription factor, p53, to the promoter of the pro-opiomelanocortin (POMC) gene, which consequently produces α-melanocyte-stimulating hormones (α-MSH), adrenocortical stimulating hormones, and β-endorphins. α-MSH, the propigmentation hormone, determines the constitutive human pigmentation and is responsible for the cutaneous response to UV, which is modulated by the G protein-combined receptor melanocortin 1 receptor (MC1R), responsible for signaling the secreted α-MSH to melanocytes [4]. Moreover, binding of α-MSH to MC1R stimulates the adenylyl cyclase (AC) dependent production of the secondary messenger cyclic adenosine monophosphate (cAMP). The increased intracellular concentration of cAMP activates protein kinase A (PKA), which phosphorylates the cAMP response element (CREB) protein. CREB is a transcription factor regulating the expression of various genes, including the expression of microphthalmia-associated transcription factor (MITF). MITF regulates the expression of melanogenesis enzymes tyrosinase-related protein 1, 2 (TRP-1, 2), and tyrosinase (TYR) [5]. TRP-1, TRP-2, and TYR proteins migrate to the melanosomes to produce eumelanin and pheomelanin [6]. Moreover, recent studies have shown the involvement of several signaling pathways, including mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3-kinase (PI3K)/AKT in the process of melanogenesis [7,8]. It has also been shown that MAPK, a family of serine/threonine kinases, including JNK, ERK, and p38 MAPK, are particularly involved in regulating MITF expression [9,10,11]. Therefore, understanding the regulation of these factors is expected to elucidate the underlying mechanism of melanogenesis induced by different compounds.
Although the exact etiopathogenesis of gray hair has not been elucidated, an aberrant melanogenesis could contribute to hair graying, which adversely affects a person’s self-esteem [12]. Moreover, recent studies have shown that the UVR-induced oxidative stress and external stimulation could cause melanocyte apoptosis leading to dysfunctional melanogenesis [12,13]. To date, dietary supplementations and hair dyes are the main therapeutic and/or cosmetic options to alleviate hair graying; however, the hair dyes could lead to adverse side effects [12]. Therefore, identifying natural, nontoxic melanogenesis-inducers has gained interest in discovering novel therapeutic agents and functional cosmetics. Coumarins (2H-benzopyran-2-ones) belong to a class of polyphenolic compounds and abundantly found in plants have been reported to exhibit different biological activities [14,15,16,17], such as anticoagulant, anti-inflammatory, anti-HIV, antioxidant, anti-allergic, anti-cancer, anti-proliferative, and antiviral activities [18,19]. Moreover, these compounds have also been reported as melanogenesis modulators [20]. Of the several compounds in this class, 4-methylumbelliferone (4MUMB), a hydroxycoumarin, has been reported to exert antimicrobial and anti-inflammatory effects [21,22,23,24]. In addition, a recent screening-level experimental study has reported the anti-melanogenic effects of 4-methylheriniarin (4MH), a derivative of 4MUMB; however, its efficacy was insignificant [25].
Several studies have reported that structurally modified coumarin derivatives influence the pharmacological and biochemical properties of the coumarins [26]. For example, acetylation replaces the active hydrogen atoms with acetyl groups in a compound, resulting in an ester-acetoxy group [27], which has been shown to increase the cytotoxicity of coumarins [28]. In addition, earlier studies have also reported the efficacies of methyl derivatives of coumarins with increased antimicrobial activities [25,29].
Therefore, in the present study to discover new melanogenic activators, we evaluated the melanogenesis-inducing effects of 4MUMB and its acetylated and methylated derivatives, 7-acetoxy-4-methylcoumarin (7A4MC), and 4MH in B16F10 murine melanoma cells and investigated the mechanism underlying their melanogenic activity. The study demonstrates the melanogenic efficacy of 7A4MC and unravels the underlying mechanism of the regulation of its melanogenic activities. The findings of the present study suggest that 7A4MC, which induces melanin production in a dose-dependent manner, could be used as an anti-whitening agent in hair cosmetics.
2. Materials and Methods
2.1. Acetylation of 4MUMB
For the acetylation experiment, 4MUMB (0.5 mmol) (Sigma-Aldrich, St. Louis, MA, USA) was dissolved in acetic anhydride (Samchun Chemical Co., Ltd., Seoul, Korea) and pyridine (Junsei Chemical Co., Ltd., Tokyo, Japan) was added. The solution was kept overnight at 90 °C under magnetic stirring. After slow cooling, the mixture was thoroughly washed with acetone and ethanol to remove unreacted acetic acid from the product and acetic anhydride. The final product was dried at 40 °C for 1 h under vacuum.
2.2. Methylation of 4MUMB
For the methylation experiment, 4MUMB (0.5 mmol) (Sigma-Aldrich, St. Louis, MA, USA) was dissolved in dimethyl carbonate (4 mL) (Sigma-Aldrich, St. Louis, MA, USA). Then, 1,8-Diazabicyclo [5.4.0]undec-7-ene (DBU; 0.6 mmol) (Sigma-Aldrich, St. Louis, MA, USA) was added. The solution was kept overnight at 90 °C under magnetic stirring. After the complete dissolution of the substrate, the solvent was evaporated as an azeotropic mixture in the presence of methanol (3 mL). The residue was then solubilized in a solution of 1 mol/L HCl (5 mL) and ethyl acetate (10 mL). The final product was dried at 40 °C for 1 h under vacuum.
2.3. High-Pressure Liquid Chromatography (HPLC) Analysis and Purification of 4MUMB, 7A4MC, and 4MH
A Shimadzu SpectroMonitor, 3200 digital UV/Vis detector, equipped with a Shim-pack GIS 0.5 mm ODS C18 column (250 mm × 4.6 mm I.D.) was used for HPLC analysis (Shimadzu, Kyoto, Japan). The mobile phase consisted of acetonitrile and water, with 0.1% Trifluoroacetic acid (TFA). A gradient method was used with a flow rate of 1 mL/min, and the analysis was performed by increasing solvent B (acetonitrile) from 10% to 100% for 30 min.
2.4. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis 7A4MC
For determination of the exact mass of different peaks, high resolution-quadrupole-time of flight-electrospray ionization-mass spectrometry (HR-QTOF ESI/MS) was performed in positive ion mode using an ACQUITY UPLC system (Waters Corp., Milford, MA, USA) coupled with an SYNAPT G2-Si C18 column (Shim-pack GIS (4.6 × 250 nm, 5 μm)). S. The obtained mass data were subsequently analyzed using MassLynx version 4.1.
2.5. Cell Culture and Viability Assay
B16F10 melanoma cells were obtained from the Korean Cell Line Bank (KCLB, Seoul, Korea). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS) with 1% penicillin-streptomycin and placed in a humidified incubator in a 5% CO2 atmosphere at 37 °C. At 80–90% confluence, cells were plated at a density of 1.0 × 104 cells/well in 24-well plates and incubated for 24 h. Cells were treated with varying concentrations of 7A4MC (12.5, 25, and 50 µM) with or without α-MSH (200 nM) for 72 h. Cell viability was measured using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. MTT reagent (Sigma-Aldrich, St. Louis, MO, USA) was added at a concentration of 1 mg/mL to each well, and cells were then incubated for 3 h. Subsequently, the formed formazan crystals were dissolved in DMSO, and the absorbance at 570 nm was recorded using a microplate reader (Bio-Tek instrument Inc., Vermont, WI, USA).
2.6. Measurement of Melanin Content
B16F10 melanoma cells were incubated in the culture medium for 24 h. The cells (4.0 × 104 cells/well) were seeded and incubated for 72 h with different concentrations of 7A4MC (12.5, 25, and 50 µM). α-MSH (200 nM) was used as a positive control. Cell pellets were harvested and lysed in 1 mL NaOH (1 mol/L) containing 10% DMSO at 90 °C. The absorbance at 405 nm was recorded using a microplate reader. Protein concentration was quantified using the bicinchoninic acid (BCA) protein assay [30].
2.7. Assessment of Intracellular Tyrosinase (TYR) Activity
B16F10 melanoma cells were incubated for 24 h. The cells (4.0 × 104 cells/well) were incubated in 6-well plates and cultured for 72 h at various concentrations of 7A4MC (12.5, 25, and 50 µM). α-MSH (200 nM) was used as positive control. After incubation, the cells were collected in a tube and centrifuged at 13,000 × g for 3 min. Next, the supernatant was removed, and the cells were lysed in radioimmunoprecipitation assay (RIPA) buffer (Bio-rad, Hercules, CA, USA) containing the protease inhibitor cocktail (1.0%). Cell lysates were vortexed for 1 h every 10 min and then centrifuged at 13,000 × g for 30 min. The same protein concentration (12.5, 25, and 50 µM) was obtained by adjusting the amount of each cell lysate using 0.1 M sodium phosphate buffer (pH 6.8). Next, 80 µL of L-3,4-dihydroxyphenylalanine (L-DOPA; 2 mg/mL) and 20 µL of each lysate were added into a 96-well plate. After incubation at 37 °C for 2 h, the absorbance was measured at 490 nm using a microplate reader.
2.8. Western Blotting
Total protein was extracted from the cells using RIPA buffer and measured with a Bradford assay kit (Pierce BCA Protein Assay Kit, Thermo Scientific, Waltham, MA, USA). Twenty micrograms of total protein samples were resolved by 10% SDS-PAGE gel and transferred onto polyvinylidene difluoride (PVDF) membranes (Bio-rad, Hercules, CA, USA) at 150 V and 2 h. Membranes were blocked with 5% skim milk at 2 h and incubated with primary antibodies, including MITF antibody (1:500), TRP-1 antibody (1:5000), TRP-2 antibody (1:500), TYR antibody (1:500), Phospho-p44/42 MAPK (Erk1/2)(Thr202/Tyr204) antibody (1:1000), Phospho-p38 MAP Kinase (Thr180/Tyr182) antibody (1:500), Phospho-SAPK/JNK (Thr183/Tyr185) antibody (1:500), p44/42 MAPK (Erk1/2) antibody (1:1000), p38 MAPK antibody (1:500), SAPK/JNK antibody (1:500), P-AKT rabbit antibody (1:1000), (all the antibodies were from Cell signaling, USA) at 4 ℃ overnight, followed by incubation with HRP Anti-Rabbit IgG secondary antibody (1:10,000) (H&L, Rockland Immunochemicals, Inc., USA) at 25 ℃ for 1 h. Proteins were detected using an ECL kit (Bio-rad, Hercules, CA, USA) and measured using an image reader (LAS-4000, FUJIFILM, Tokyo, Japan).
2.9. Statistical Analysis
All the experiments were conducted in triplicate, and the results are expressed as mean ± SD (standard deviation). The statistical significance of the differences was evaluated using Student’s t-test for the data acquired.
3. Results
3.1. Acetylation of 4MUMB and Methylation of 4MUMB
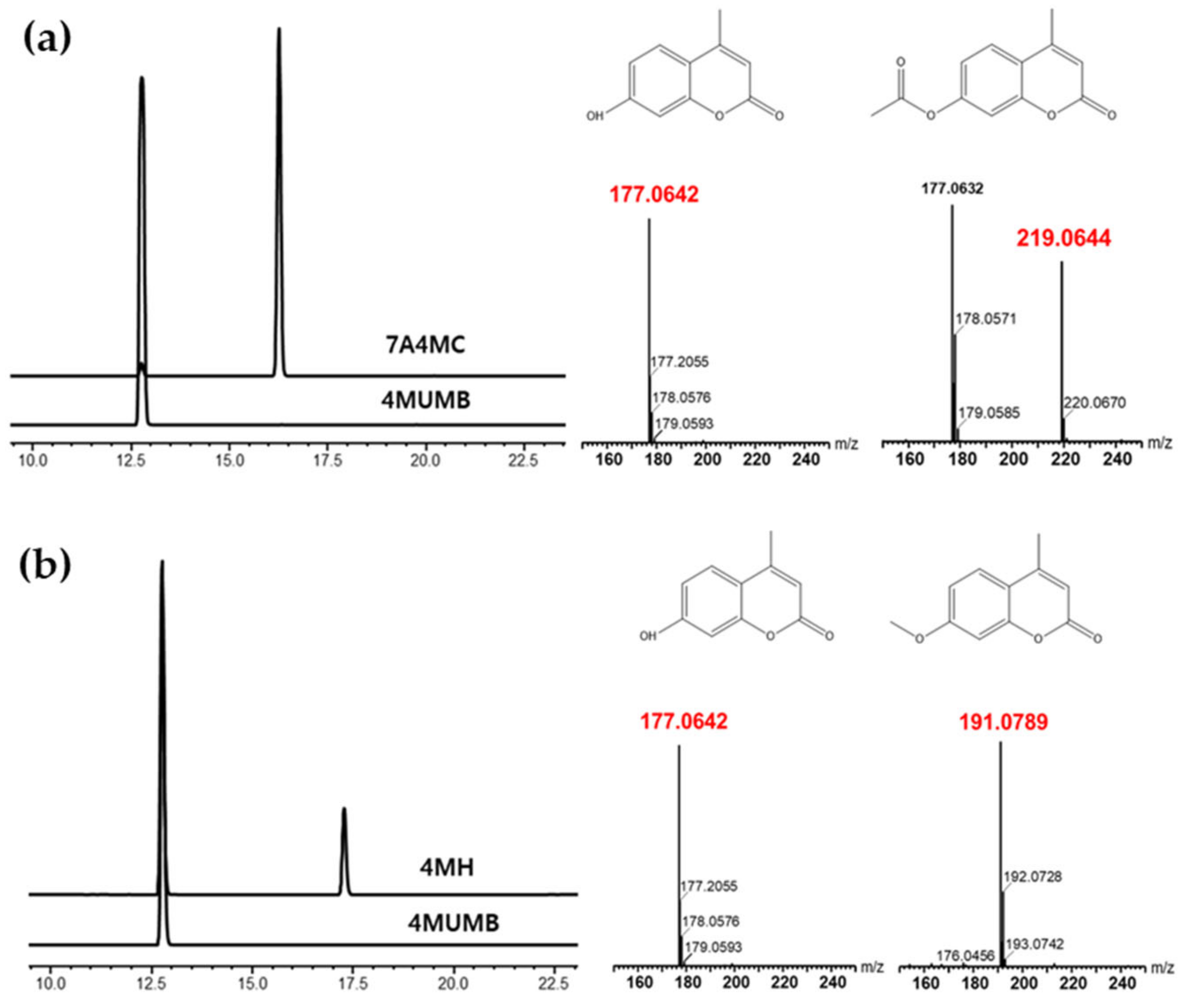
We performed HPLC and mass analysis for the synthesis and purification of the derivatives of 4MUMB. The acetylation and methylation of 4MUMB replaced its hydrogen atom with an acetyl group and a methyl group, respectively. The reaction products were detected and analyzed by HPLC (Figure 1a,b). Acetylation and methylation reactions were investigated using electrospray ionization-mass spectrometry (ESI/MS), and the resulting 7A4MC and 4MH were purified by preparative HPLC. As shown in Figure 1, the synthesis of 7A4MC and 4MH was confirmed through HPLC analysis, and the molecular weights of 4MUMB, 7A4MC, and 4MH were obtained through HR-ESI/MS and mass analysis. The molecular weights of these three compounds were HR-ESI/MS: m/z [M+H] 176.0624, calcd. 176.0624, HR-ESI/MS: m/z [M+H] 219.0644, calcd. 219.0644, and HR-ESI/MS: m/z [M+H] 191.0789, calcd. 191.0789, respectively. Subsequently, a comparison of the HPLC data of the synthesized 7A4MC and 4MH and the purchased 7A4MC and 4MH (Tokyo Chemical Industry, Kita-Ku, Tokyo, Japan) confirmed the same peak, whereas mass analysis confirmed their molecular weight and retention time (data not shown). Although it was synthesized, we speculated that the purchased product might possess a stereoisomer imparting an altered efficacy; however, we could not record any difference in their efficacies.
3.2. Effects of 4MUMB, 7A4MC, and 4MH on the Viability of B16F10 Cells
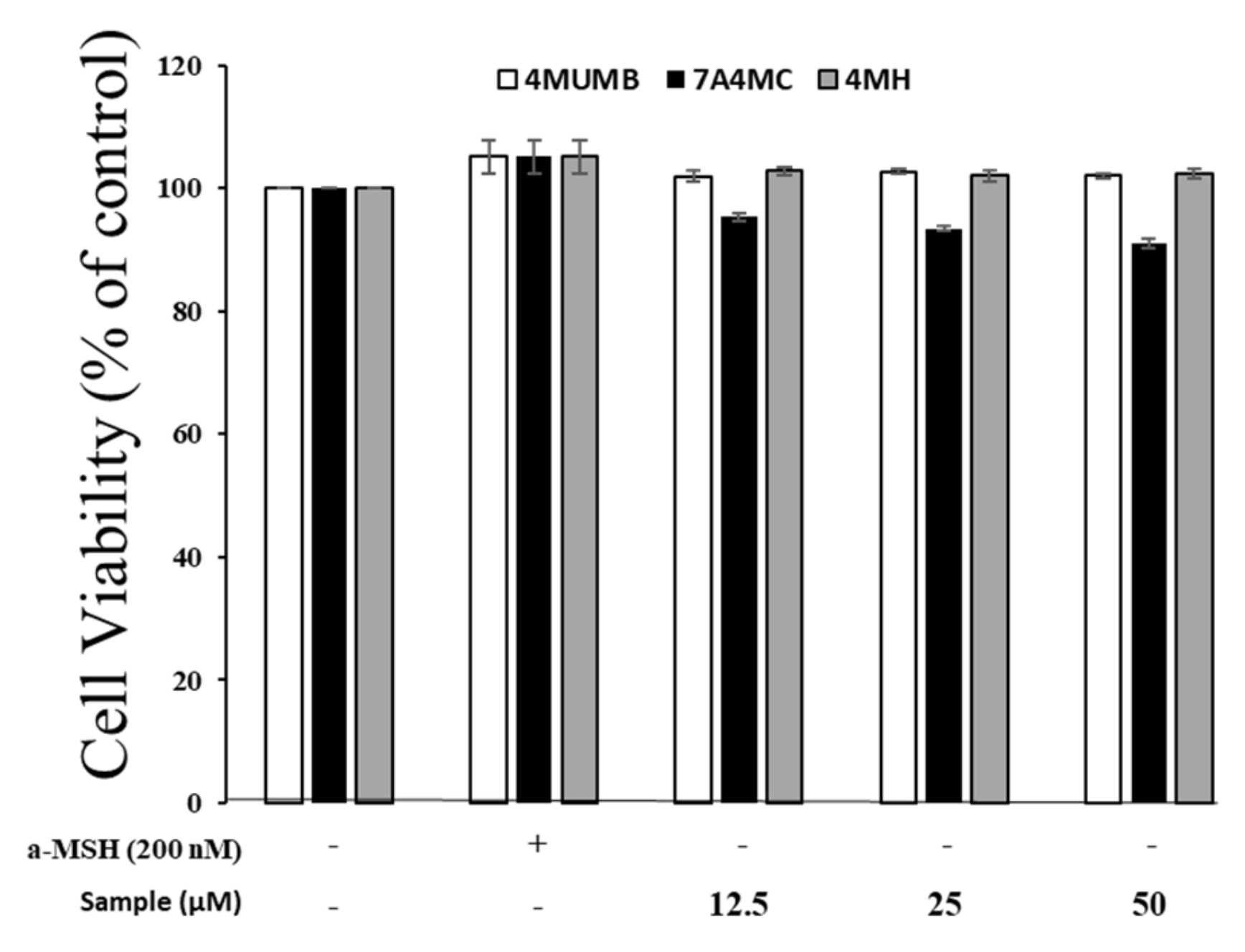
After synthesis and purification, the cytotoxicity of 7A4MC and 4MH in B16F10 cells were tested by MTT assay. B16F10 cells were treated with 4MUMB, 7A4MC, and 4MH (12.5, 25, and 50 µM) for 72 h to determine the concentration applied to the next experiment. α-MSH (200 nM) and DMEM were used as controls. As shown in Figure 2, there were no significant differences in proliferation rates between the control and 4MUMB, 7A4MC, and 4MH treated cells. These results indicate that the effect of 4MUMB, 7A4MC, and 4MH on melanin biosynthesis are not attributable to cell proliferation at the above mentioned concentrations.
3.3. Effects of 4MUMB, 7A4MC, and 4MH on Melanin Production
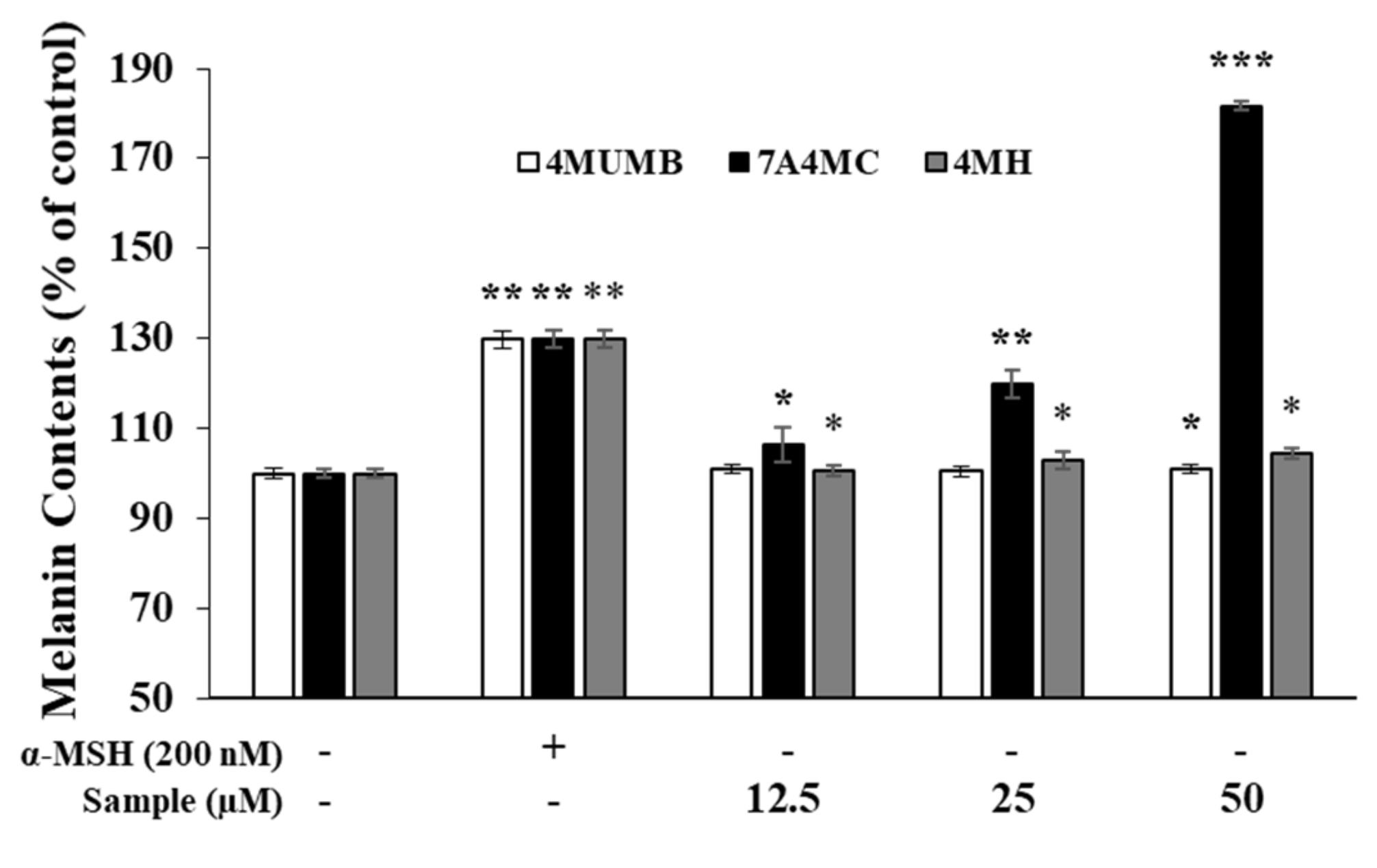
To assess the melanogenic activity of 4MUMB, 7A4MC, and 4MH, we measured melanin production in α-MSH-untreated B16F10 murine melanoma cells. As shown in Figure 3, in α-MSH-untreated B16F10 cells, 4MUMB did not show any significant change, whereas 4MH slightly induced the melanin production in a dose-dependent manner. Furthermore, 7A4MC produced the highest amount of melanin in a dose-dependent manner and was used for subsequent analyses.
3.4. Effect of 7A4MC on TYR Activity in B16F10 Melanoma Cells
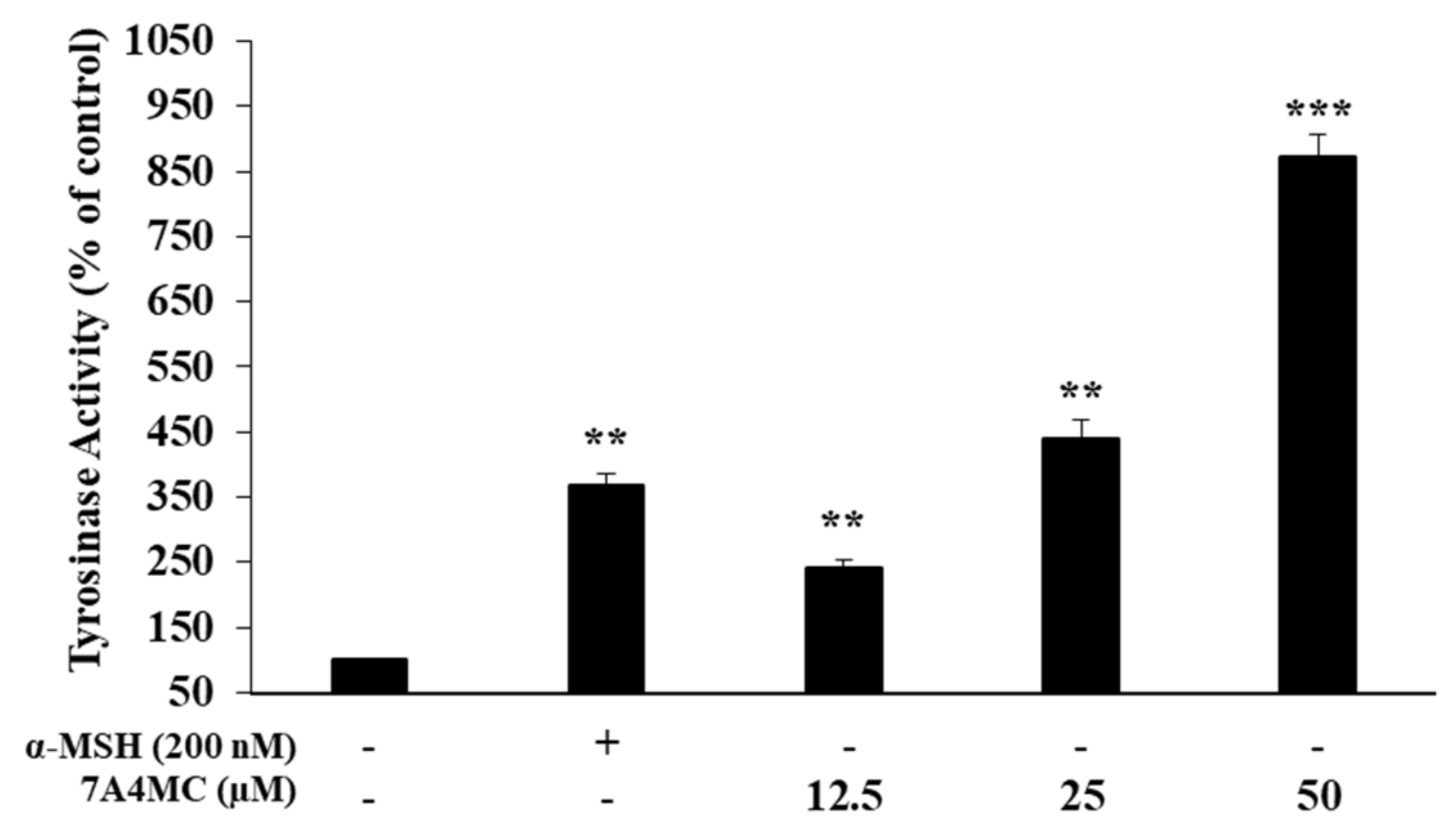
Next, we examined the effect of 7A4MC on the regulation of TYR activity in B16F10 melanoma cells. As shown in Figure 4, the TYR activities were significantly increased up to approximately 141.8%, 340.5%, and 771.7% compared to the control. These results suggest that 7A4MC increases melanogenesis through the upregulation of TYR activities in B16F10 melanoma cells. In addition, given that the stimulant was not treated, we expect 7A4MC to be used as a tanning agent and hair dye.
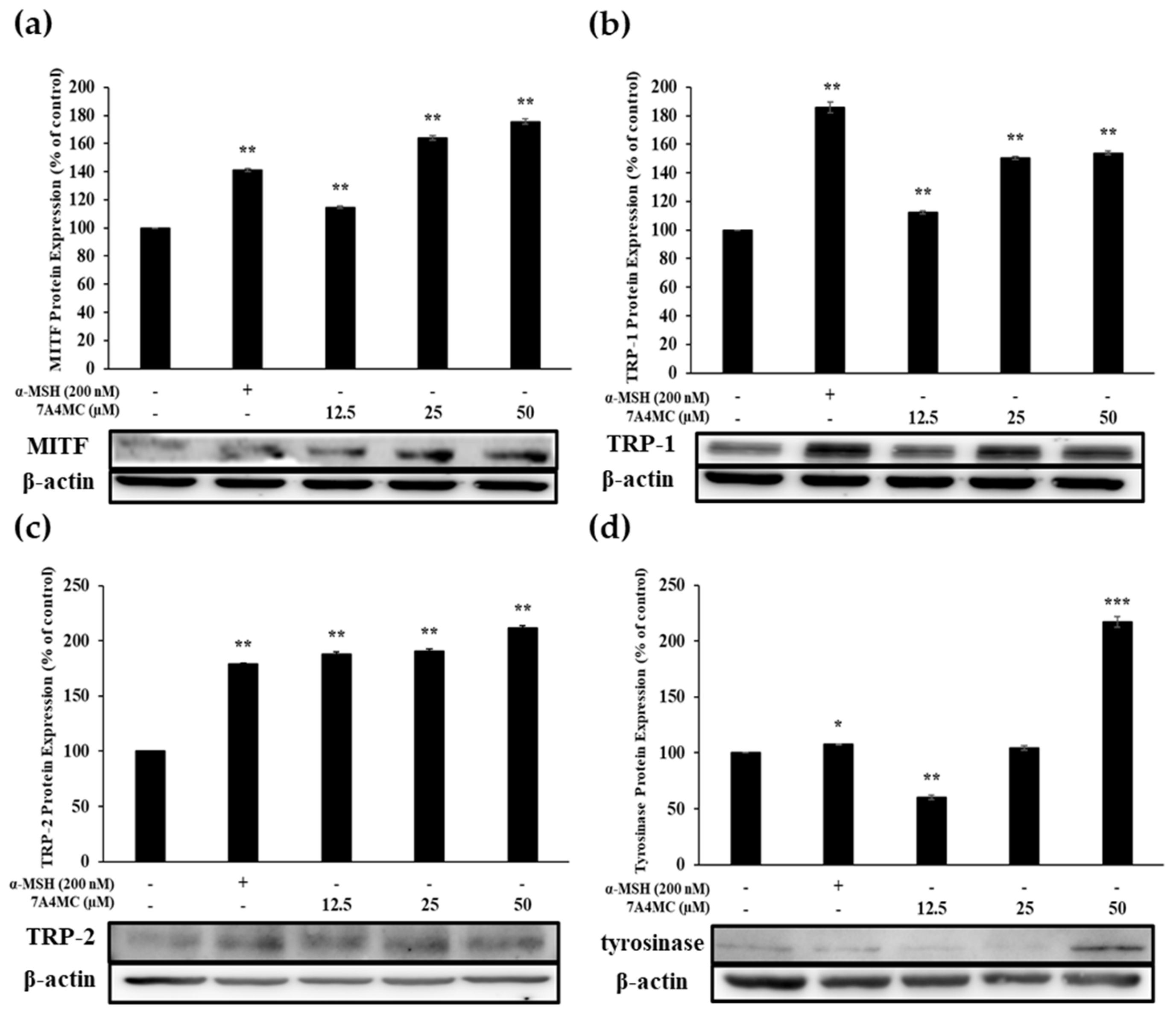
3.5. Effect of 7A4MC on the Protein Expression of MITF and Melanogenic Enzymes
To identify the mechanism by which 7A4MC stimulates melanin biosynthesis, we examined the expression of MITF and target genes, including TRP-1, TRP-2, and TYR, through Western blot analysis.
MITF protein expression was increased in B16F10 cells treated with 7A4MC for 48 h in a dose-dependent manner (Figure 5a). In addition, as shown in Figure 5b–d, B16F10 murine melanoma cells treated with 7A4MC showed a significantly increased expression of TRP-1, TRP-2, and TYR. These data imply that 7A4MC treatment increases MITF expression, which in turn upregulates TRP-1, TRP-2, and TYR levels.
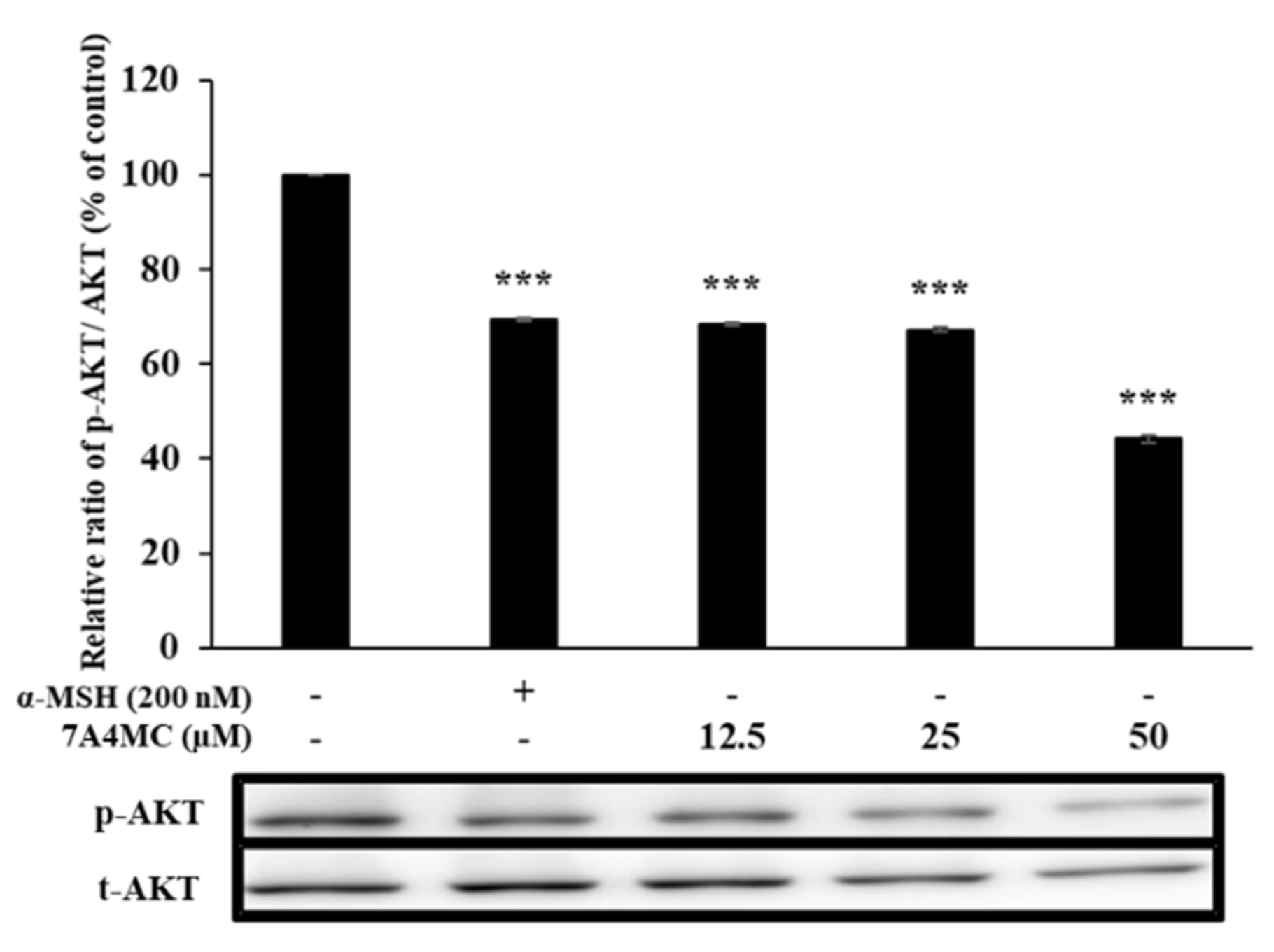
3.6. 7A4MC Suppresses p-AKT Expression in B16F0 Melanoma Cells
Next, we investigated the AKT signaling pathways, the upstream signaling pathways to elucidate the underlying mechanism of the pigmentation effect of 7A4MC. The expression analysis of AKT in 7A4MC treated cells by Western blot showed a significant reduction in AKT phosphorylation in a dose-dependent manner with the maximum reduction at a concentration of 50 µM (Figure 6). These findings indicate that the melanogenesis-stimulating effects of 7A4MC could be exerted through reduced AKT phosphorylation.
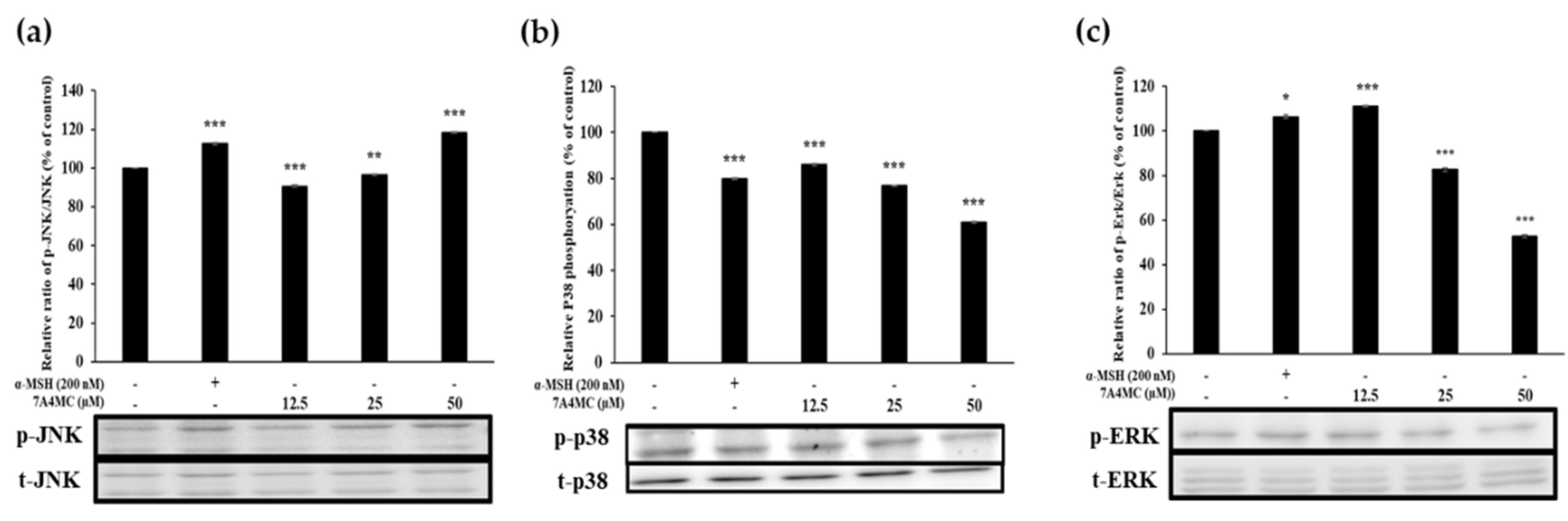
3.7. 7A4MC Increases p-JNK, p-p38, and p-ERK Expression in the MAPK Pathway
The MAPK signaling pathway has been implicated in regulating MITF expression. MAPK family proteins, including JNK, p38, and ERK1/2, play an important role in melanogenesis. Therefore, we investigated the effects of 7A4MC on JNK protein expression levels. The results revealed an increasing trend of expression with increasing concentrations of 7A4MC (Figure 7a). However, the expression of p38 and ERK showed the opposite trend to that of JNK (Figure 7b,c). These results indicated that the effect of 7A4MC on the MAPK signaling pathway increased the expression of MITF with an increased protein level of JNK, which consequently increased the protein expression of TRP-1, TRP-2, and TYR.
4. Discussion
The melanin present in the skin, hair, and eyes protects the skin from UVR [31,32]. In the present study, we investigated the melanogenic effects of 4MUMB, a hydroxycoumarin and its derivatives, 7A4MC and 4MH. The comparison of the synthesized and purchased 7A4MC and 4MH confirmed the purity of the compounds with the same molecular weight and retention time. Moreover, we did not observe any alterations in their efficacy, which further confirmed the absence of stereoisomers. We demonstrated the 7A4MC effect on melanin formation in B16F10 melanoma cells. The MTT assay conducted to test the cytotoxicity of 4MUMB, 7A4MC, and 4MH, ruled out the possibilities of regulating melanogenesis by these compounds through cell proliferation; moreover, the tested concentrations were found to be non-cytotoxic. However, of the tested compounds, 7A4MC markedly increased the melanin levels in a dose-dependent manner, whereas the effects of 4MUMB and 4MH were not significant. On the contrary, 4MH has been reported to have a slight inhibitory effect on melanogenesis [25]. 7A4MC upregulated the activity of TRP-1, TRP-2, and TYR, the most important enzyme for melanin synthesis. Furthermore, 7A4MC reduced the expression of ERK, p38; however, it increased the expression of p-JNK, which in turn increased the MITF expression that upregulated TRP-1, TRP-2, and TYR levels, leading to heightened melanin synthesis. In addition, 7A4MC reduced the expressions of AKT, the protein involved in the PI3K/AKT signaling pathway, in a concentration-dependent manner. These results indicate the melanogenesis effect of 7A4MC is executed through the upregulation of the MAPK/JNK signaling pathways but downregulation of the AKT pathway. On the contrary, Chung et al. have reported that pratol, an O-methylated flavone, induces melanogenesis in B16F10 melanoma cells via p-p38 and p-JNK upregulation [33]. Therefore, it can be concluded that the pathway for melanogenesis activation is not conserved; instead, it is dependent on the melanogenesis-inducing compounds.
Here, we demonstrated that 7A4MC synthesized through acetylation of 4MUMB showed strong melanogenesis efficacy. However, previous studies have shown inhibitory effects of acetylated products on melanogenesis; for example, studies have shown that the whitening effect of the acetylated arbutin is better than that of arbutin, which is used as a representative whitening functional ingredient in cosmetics [34]. Taken together, it can be inferred that acetylation does not have absolute whitening or melanogenesis effects. Furthermore, the melanogenesis-inducing effects of 7A4MC could be attributed to its structural similarity with 4MUMB, whereas the structural difference between the acetyl group bond and the methyl group bond in 4MH was speculated to be responsible for its non-significant melanogenesis effects.
In conclusion, the findings show that 7A4MC increased melanin production without significant cytotoxicity. It increases TYR activity and enhances the expression of MITF, which concomitantly induces the expression of TRP-1, TRP-2, and TYR. Furthermore, the study demonstrates that 7A4MC stimulates melanogenesis via increased phosphorylation of JNK, ERK, and p38 and reduced AKT phosphorylation. The findings of the present study indicate the potential use of 7A4MC as an anti-hair bleaching or tanning agent in cosmetics industries. However, further studies using animal models are required to validate the findings of the present study.
Author Contributions
Conceptualization, S.Y.K.; Data curation, J.H.S., S.J., T.P., and S.Y.K.; Supervision, S.Y.K.; Writing—original draft, J.H.S.; Writing—review and editing, W.-J.C., and S.-Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR202020101).
Acknowledgments
This research was financially supported by the Ministry of Small and Medium-sized Enterprises(SMEs) and Startups(MSS), Korea, under the “Regional Enterprise linked with National-Innovation-Cluster Development Program(R&D, P0009943)” supervised by the Korea Institute for Advancement of Technology(KIAT).
Conflicts of Interest
The authors declare no conflict of interest.
References
- González, S.; Fernández-lorente, M.; Gilaberte-Calzada, Y. The latest on skin photoprotection. Clin Dermatol. 2008, 26, 614–626. [Google Scholar] [CrossRef]
- Maverakis, E.; Miyamura, Y.; Bowen, M.P.; Correa, G.; Ono, Y.; Goodarzi, H. Light, including ultraviolet. J. Autoimmu. 2010, 34, J247–J257. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Xu, J.; Long, M.; Tu, Z.; Yang, G.; He, G. 2, 3, 5, 40-tetrahydroxystillbene-2-O-d-glucoside (THSG) induces melanogenesis in B16 cells by MAP kinase activation and tyrosinase upregulation. Life Sci. 2009, 85, 345–350. [Google Scholar] [CrossRef]
- Moon, S.H.; You, C.C.; Hyun, C.G. Tobramycin promotes melanogenesis by upregulating p38 MAPK protein phosphorylation in B16F10 melanoma cells. Antibiotics 2019, 8, 140. [Google Scholar] [CrossRef] [Green Version]
- Aroca, P.; Urabe, K.; Kobayashi, T.; Tsukamoto, K.; Hearing, V.J. Melanin biosynthesis patterns following hormonal stimulation. J. Biol. Chem. 1993, 268, 25650–25655. [Google Scholar]
- Slominski, A.; Wortsman, J.; Plonka, P.M.; Schallreuter, K.U.; Paus, R.; Tobin, D.J. Hair follicle pigmentation. J. Investig. Dermatol. 2005, 124, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Imesch, P.D.; Wallow, I.H.; Albert, D.M. The color of the human eye: A review of morphologic correlates and of some conditions that affect iridial pigmentation. Surv. Ophthalmol. 1997, 41, S117–S123. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, S.Y.; Chung, J.H.; Kim, K.H.; Eun, H.C.; Park, K.C. Delayed ERK activation by ceramide reduces melanin synthesis in human melanocytes. Cell Signal. 2002, 14, 779–785. [Google Scholar] [CrossRef]
- Yao, C.; Jin, C.L.; Oh, J.H.; Oh, I.G.; Park, C.H.; Chung, J.H. Ardisia crenata extract stimulates melanogenesis in B16F10 melanoma cells through inhibition ERK1/2 and Akt activation. Mol. Med. Rep. 2015, 11, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Jin, S.H.; Kang, H.Y. LPS induces melanogenesis through p38 MAPK activation in human melanocytes. Arch. Dermatol. Res. 2008, 300, 325–329. [Google Scholar] [CrossRef]
- Widlude, H.R.; Fisher, D.E. Microphthalmia-associated transcription factor: A critical regulator of pigment cell development and survival. Oncoqene 2003, 22, 3035–3041. [Google Scholar]
- Anagha, B.K.; Huma, S.; Umashankar, N. Premature Graying of Hair: Review with Updates. Int. J. Trichology 2018, 10, 198–203. [Google Scholar]
- Bing, Z.; Sai, M.; Inbal, R.; Megan, H.; Pankaj, B.; Sekyu, C.; William, A.G.; Yulia, S.; Eva, M.F.; Yiqun, S.; et al. Hyperactivation of sympathetic nerves drives depletion of melanocyte stem cells. Nature 2020, 577, 676–681. [Google Scholar]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signaling to CREB. Pigment Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef]
- Borges, F.; Roleira, F.; Milhazes, N.; Uriarte, E.; Santana, L. Simple coumarins: Priviled scaffolds in medicinal chemistry. Front. Med. Chem. 2009, 4, 23–85. [Google Scholar]
- Matos, M.J.; Terán, C.; Pérez-Castillo, Y.; Uriarte, E.; Santana, L.; Viña, D. Synthesis and study of a series of 3-arylcoumarins as potent and selective monoamine oxidase B inhibitors. J. Med. Chem. 2011, 54, 7127–7137. [Google Scholar] [CrossRef]
- Matos, M.J.; Viña, D.; Vázquez-Rodríguez, S.; Uriarte, E.; Santana, L. Focusing on new monoamine oxidase inhibitors: Differently substituted coumarins as an interesting scaffold. Curr. Top. Med. Chem. 2012, 12, 2210–2239. [Google Scholar] [CrossRef]
- Pérez-Cruz, F.; Serra, S.; Delogu, G.; Lapier, M.; Maya, J.D.; Olea-Azar, C.; Santana, L.; Uriarte, E. Antitrypanosomal and antioxidant properties of 4-hydroxycoumarins derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 5569–5573. [Google Scholar] [CrossRef]
- Khan, Z.A.; Rama, N.H.; Hussain, M.T.; Khan, G.S. Synthesis and antimicrobial evaluation of some new 3-alkyl-,3-(2’-naphthyl)isocoumarins and their (dl)-3,4 dihydroderivatives. Indian J. Chem. 2007, 46B, 1322–1327. [Google Scholar] [CrossRef]
- Seth, J.O.; Li, N.K. Coumarin compounds as melanogenesis modifiers and uses thereof. United. States Patent Application No. 0220545 A1, 30 August 2012. [Google Scholar]
- Balaji, P.N.; Sreevani, M.S.; Harini, P.; Johnsi, R.P.; Prathusha, K.; Chandu, T.J. Antimicrobial activity of some novel synthesized heterocyclic compounds from substituted chalcones. J. Chem. Pharm. Res. 2010, 2, 754–758. [Google Scholar]
- Piccioni, F.; Malvicini, M.; Garcia, M.G.; Rodriguez, A.; Atorrasagasti, C.; Kippes, N.; Buena, I.T.P.; Rizzo, M.M.; Bayo, J.; Aquino, J.; et al. Antitumor effects of hyaluronic acid inhibitor 4-methylumbelliferone in an orthotopic hepatocellular carcinoma model in mice. Glycobiology 2011, 22, 400–410. [Google Scholar] [CrossRef]
- Piccioni, F.; Fiore, E.; Bayo, J.; Atorrasagasti, C.; Peixoto, E.; Rizzo, M.; Malvicini, M.; Tirado-González, I.; García, M.G.; Alaniz, L.; et al. 4-methylumbelliferone inhibits hepatocellular carcinoma growths by decreasing IL-6 production and angiogenesis. Glycobiology 2015, 25, 825–835. [Google Scholar] [CrossRef] [Green Version]
- Al-Amiery, Y.K.; Al-Amiery, A.; Kadhum, A.A.H.; Mohamad, A.B. Antioxidant activities of 4-methylumbelliferone derivatives. PLoS ONE 2016, 11, e0156625. [Google Scholar]
- Balaji, P.N.; Seleena, S.K.P.; Vidya, G.; Himateja, T.; Neelima, B.; Lakshmiteja, T. Synthesis and charecterisation of Schiff bases derived from acetyl coumarin and evaluated for anti-micribial activity. Int. J. Drug Dev. Res. 2013, 4, 0975–8585. [Google Scholar]
- Musiliyu, A.M.; Jonh, S.C.; Khan, M.O.F. A review of coumarin derivatives in pharmacotherapy of breast cancer. Curr. Med. Chem. 2008, 15, 2664–2679. [Google Scholar]
- Zhao, D.Y.; Hao, Q.X.; Kang, L.P.; Zhang, Y.; Chen, M.L.; Wang, T.L.; Guo, L.P. Advance in Studying Early Bolting of Umbelliferae Medicinal Plant. China J. Chin. Mater. Med. 2016, 41, 20–23. [Google Scholar]
- Musiliyu, A.M.; Veera, L.D.B.; Lekan, M.L.; John, C.; Andre, S.; Ahkinyala, A. Cytotoxic activity of new acetoxycoumarin derivatives in cancer cell lines. Anticancer Res. 2011, 31, 2017–2022. [Google Scholar]
- Bernini, R.; Crisante, F.; Ginnasi, M.C. A comvenient and safe O-methylation of flavonoids with dimethyl carbonate (DMC). Molecules 2011, 16, 1418. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.; Mian, L.; Bo, H. Competitive binding to cuprous lons of protein and BCA in the bicinchoninic acid protein assay. Biomed. Eng. J. 2010, 4, 271–278. [Google Scholar]
- Gratacap, B.; Attard, A.; Laurent, A.; Stoebner, P.; Smirou, D.; Charachon, R. Melanin in the inner ear. An experimental study with control and kanamycin-intoxicated colored guinea-pigs. Arch. Oto Rhino Laryngol. 1989, 246, 235–237. [Google Scholar] [CrossRef]
- Zecca, L.; Bellei, C.; Costi, P.; Albertini, A.; Monzani, E.; Casella, L.; Gallorini, M.; Bergamaschi, L.; Moscatelli, A.; Turro, N.J.; et al. New melanic pigments in the human brain that accumulate in aging and block environmental toxic metals. Proc. Natl. Acad. Sci. USA 2008, 105, 17567–17572. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.C.; Kim, J.H.; Lee, G.S.; Lee, J.N.; Lee, N.H.; Hyun, C.G. Pratol, an O-methylated flavone, induces melanogenesis in B16F10 melanoma cells via p-p38 and p-jnk upregulation. Molecules 2017, 22, 1704. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, D.; Zhang, Y.; Li, J.; Wu, Z.; Wang, Z.; Wang, D. Investigation of the pro-apoptotic effects of arbutin and its acetylated derivative on murine melanoma cells. Int. J. Mol. Med. 2018, 41, 1048–1054. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(a) High-pressure liquid chromatography (HPLC) analysis of 4-methylumbelliferone (4MUMB), 7-acetoxy-4-methylcoumarin (7A4MC), Structure and Mass analysis of 4MUMB and 7A4MC. (b) HPLC analysis of 4MUMB, 4-methylheriniarin (4MH), Structure and Mass analysis of 4MUMB and 4MH.
Figure 1.
(a) High-pressure liquid chromatography (HPLC) analysis of 4-methylumbelliferone (4MUMB), 7-acetoxy-4-methylcoumarin (7A4MC), Structure and Mass analysis of 4MUMB and 7A4MC. (b) HPLC analysis of 4MUMB, 4-methylheriniarin (4MH), Structure and Mass analysis of 4MUMB and 4MH.

Figure 2.
Cell viability of 4MUMB, 7A4MC, and 4MH-treated B16F10 mouse cells with (+) or without (-) α-MSH. The cells were supplemented with different concentrations (12.5, 25, and 50 µM) of 4MUMB, 7A4MC, and 4MH for 72 h and the proliferation of the cells was assessed.
Figure 2.
Cell viability of 4MUMB, 7A4MC, and 4MH-treated B16F10 mouse cells with (+) or without (-) α-MSH. The cells were supplemented with different concentrations (12.5, 25, and 50 µM) of 4MUMB, 7A4MC, and 4MH for 72 h and the proliferation of the cells was assessed.

Figure 3.
Effects of 4MUMB, 7A4MC, and 4MH on melanin production in B16F10 melanoma cells treated with (+) or without (-) α-MSH. The cells were treated with 4MUMB, 7A4MC, and 4MH (12.5, 25, and 50 µM) for 72 h. α-MSH (200 nM) was used as positive control. The data represent the mean ± SD of quadruple experiments. * p < 0.05, ** p < 0.01, *** p < 0.005.
Figure 3.
Effects of 4MUMB, 7A4MC, and 4MH on melanin production in B16F10 melanoma cells treated with (+) or without (-) α-MSH. The cells were treated with 4MUMB, 7A4MC, and 4MH (12.5, 25, and 50 µM) for 72 h. α-MSH (200 nM) was used as positive control. The data represent the mean ± SD of quadruple experiments. * p < 0.05, ** p < 0.01, *** p < 0.005.

Figure 4.
Effects of 7A4MC on tyrosinase (TYR) activity. B16F10 cells were treated with 7A4MC (12.5, 25, and 50 µM) for 72 h. α-MSH (200 nM) was used as positive control. The data represent the mean ± SD of quadruple experiments. ** p < 0.01, *** p < 0.005.
Figure 4.
Effects of 7A4MC on tyrosinase (TYR) activity. B16F10 cells were treated with 7A4MC (12.5, 25, and 50 µM) for 72 h. α-MSH (200 nM) was used as positive control. The data represent the mean ± SD of quadruple experiments. ** p < 0.01, *** p < 0.005.

Figure 5.
Effect of 7A4MC on MITF, TRP-1, 2, and TYR expression in B16F10 melanoma cells. Cells were treated with different concentrations of 7A4MC (12.5, 25, and 50 µM) for 48 h. α-MSH (200 nM) was used as positive control. (a) Result of Western blot and protein level of MITF, (b) TRP-1, (c) TRP-2, and (d) TYR. The data represent the mean ± SD of triplicate experiments. * p < 0.05, ** p < 0.01, *** p < 0.005.
Figure 5.
Effect of 7A4MC on MITF, TRP-1, 2, and TYR expression in B16F10 melanoma cells. Cells were treated with different concentrations of 7A4MC (12.5, 25, and 50 µM) for 48 h. α-MSH (200 nM) was used as positive control. (a) Result of Western blot and protein level of MITF, (b) TRP-1, (c) TRP-2, and (d) TYR. The data represent the mean ± SD of triplicate experiments. * p < 0.05, ** p < 0.01, *** p < 0.005.

Figure 6.
Effects of 7A4MC on the phosphorylation of p-AKT. B16F10 cells were treated with 7A4MC at the indicated concentrations for 4 h—the result of Western blot and protein level of p-AKT. The data represent the mean ± SD of triplicate experiments. *** p < 0.005. p: Phosphorylated, t: Total.
Figure 6.
Effects of 7A4MC on the phosphorylation of p-AKT. B16F10 cells were treated with 7A4MC at the indicated concentrations for 4 h—the result of Western blot and protein level of p-AKT. The data represent the mean ± SD of triplicate experiments. *** p < 0.005. p: Phosphorylated, t: Total.

Figure 7.
Effect of 7A4MC on the phosphorylation of p-JNK, p-p38, p-ERK. B16F10 cells were treated with 7A4MC at different concentrations for 4 h. (a) Results of Western blot and protein level of p-JNK, (b) p-p38, and (c) p-ERK. Results are expressed as a percentage of the control. The data represent the mean ± SD of triplicate experiments. * p < 0.05, ** p < 0.01, *** p < 0.005. p: Phosphorylated, t: Total.
Figure 7.
Effect of 7A4MC on the phosphorylation of p-JNK, p-p38, p-ERK. B16F10 cells were treated with 7A4MC at different concentrations for 4 h. (a) Results of Western blot and protein level of p-JNK, (b) p-p38, and (c) p-ERK. Results are expressed as a percentage of the control. The data represent the mean ± SD of triplicate experiments. * p < 0.05, ** p < 0.01, *** p < 0.005. p: Phosphorylated, t: Total.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sim, J.-H.; Jang, S.-C.; Park, T.-J.; Chi, W.-J.; Kim, S.-Y. Melanogenesis Effect of 7-acetoxy-4-methylcoumarin in B16F10 Melanoma Cells. Cosmetics 2020, 7, 94. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040094
AMA Style
Sim J-H, Jang S-C, Park T-J, Chi W-J, Kim S-Y. Melanogenesis Effect of 7-acetoxy-4-methylcoumarin in B16F10 Melanoma Cells. Cosmetics. 2020; 7(4):94. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040094
Chicago/Turabian StyleSim, Ji-Han, Sung-Chan Jang, Tae-Jin Park, Won-Jae Chi, and Seung-Young Kim. 2020. "Melanogenesis Effect of 7-acetoxy-4-methylcoumarin in B16F10 Melanoma Cells" Cosmetics 7, no. 4: 94. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics7040094
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.