Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation

1
Jeju Inside Agency & Cosmetic Science Center, Department of Chemistry and Cosmetics, Jeju National University, Jeju 63243, Korea
2
CoseedBioPharm Co., Ltd., Jeju 63309, Korea
3
Bio-Convergence Center, Jeju Technopark, Jeju 63243, Korea
*
Author to whom correspondence should be addressed.
Cosmetics 2021, 8(1), 22; https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010022
Submission received: 1 February 2021
/
Revised: 18 February 2021
/
Accepted: 12 March 2021
/
Published: 15 March 2021
(This article belongs to the Special Issue Discovering and Exploiting Natural Cosmetic Materials and Their Functional Mechanisms)
Abstract
:In this study, in order to explore the anti-melanogenic effect of PFE (Paederia foetida L. extract) and suggest its availability, B16F10 cells, which are murine melanoma cells, were stimulated with alpha-Melanocyte-stimulating hormone (α-MSH) to conduct an in vitro experiment. Treatment with PFE in B16F10 cells with activated melanogenesis due to stimulants showed that PFE significantly inhibits melanin content as well as intracellular tyrosinase activity within a range that does not cause cytotoxicity. In addition, Western blot assay demonstrated that PFE strongly inhibited the protein expression of not only tyrosinase-related protein (TRP)-1, -2, and tyrosinase, but also microphthalmia-associated transcription factor (MITF). Moreover, mechanism studies have shown that PFE processing inhibited the activation of melanin production by regulating the phosphorylation of each mitogen-activated protein kinase (MAPK) family in the MAPK signaling pathway. To test the biocompatibility of PFE on human skin, a primary skin irritation test was performed. The results revealed that PFE did not have any side effects on human skin. These findings suggest that PFE holds great potential as a skin whitening agent and in the prevention of hyperpigmentation disorders.
Keywords:
Paederia foetida L.; anti-melanogenesis; MAPK; skin whitening; B16F10 melanoma cell; bioactivity; ERK; JNK1. Introduction
Melanin is present extensively in the tissues of the skin, eyes, and hair of mammals. Melanin is synthesized in melanosomes in the cytoplasm of melanocytes, which are dendritic cells; its expression is regulated by various mechanisms within the cells and is transferred to surrounding keratinocytes. Melanin is composed of brownish-black eumelanin as well as reddish-yellow pheomelanin. In particular, the amount of eumelanin plays a crucial role in determining the coloration in mammals. The major act of melanin is absorbing ultraviolet radiation and blocking its penetration into the human body, protecting the skin from photo-damage [1,2,3,4,5,6]. Thus, lack of melanin from the epidermis exposes skin to harmful UV light, and this may cause hypopigmentation disorders, including vitiligo, albinism, and grey hair [7,8]. However, excessive accumulation in the skin due to abnormal over-expression of melanin pigments can also lead to the development of skin disease, including lentigo, melanoma, blotches, and freckles, etc. [9,10].
Important enzymes involved in melanogenesis include tyrosinase and tyrosinase-related protein 1, 2 (TRP-1, -2). L-Tyrosine, an essential amino acid in the initial stages of melanin synthesis, which is converted to dihydroxyl-L-phenylalanine (L-DOPA) through hydroxie reaction by tyrosinase, and L-DOPA is oxidized to DOPAquinone, and then DOPAchrome. The DOPAchrome obtained thusly is converted to 5,6-dihydroxy-2-indolylcarboxylic acid (DHICA) by TRP-2. Ultimately, DHICA is transformed into in-dole-5,6-quinone-carboxylic acid by TRP-1 to produce black or brown eumelanin. Tyrosinase is a direct influence throughout the synthesis of eumelanin and pheomelanin, and TRP-1 and TRP-2 are known to be closely linked to the synthesis of eumelanin [11,12,13,14].
Microphthalmia-associated transcription factor (MITF), which is an important transcription modulating factor in melanin synthesis, binds to the tyrosinase promoter, M-box, to facilitate the transcription of tyrosinase, TRP-1, and TRP-2 within the nucleus of melanocyte. Therefore, the effect of suppressing MITF expression of the sample becomes an important indicator for whitening studies [15,16,17].
In addition, it has been observed in previous research that MITF expression is controlled by phosphorylation of mitogen-activated protein kinases (MAPK) [18,19,20]. Moreover, it also has been explored that inhibition of p38 and JNK, along with the activation of ERK phosphorylation, reduced MITF and melanogenic enzyme expression, resulting in the downregulation of melanogenesis [21,22,23].
Whitening agents currently used in the cosmetic field include arbutin and kojic acid, which inhibit the tyrosinase activity, and L-ascorbic acid and glutathione, which inhibit melanin synthesis. However, since these compounds are decomposed or have low stability, they are used only in a limited amount as additives, so development of whitening agents using natural products is required [24].
Paederia foetida L., an odorous sulfurous-smelling plant belonging to the Rubiaceae family, is widely distributed across Asia. It has been traditionally used in folk medicine to treat various gastrointestinal disorders such as diarrhea and dysentery, stomach edema, gastritis, abdominal pain, and indigestion [25,26]. The leaves are also used as anti-ulcer agents [27]. Paederia foetida L. possesses strong pharmaceutical bioactivities owing to its anti-cancer properties in human prostate cancer cells, along with it its anti-arthritic, anti-diarrheal, anti-inflammatory, as well as gastroprotective activities [28,29,30,31]. However, the anti-melanogenic effects of Paederia foetida L. have not been thoroughly investigated to date. Therefore, this study confirmed whether there is an effect of inhibiting melanin production by treating Paederia foetida L. extract (PFE) to B16F10 cells with increased melanin production due to alpha-Melanocyte-stimulating hormone (α-MSH) stimulation. In addition, it also checked how PFE regulates each signaling pathway in the cell through a mechanism study.
2. Materials and Methods
2.1. Paederia foetida L. Extract
Paederia foetida L. was obtained from CoseedBioPharm Co., Ltd. (Jeju, Korea). Air-dried Paederia foetida L. (20 g) was mixed with 70% EtOH (1.2 L) and incubated overnight at 21–23 °C. Subsequently, the extract was filtered three times using filter paper, and the solvent was evaporated to obtain a solid extract. The extract was powdered by freeze-drying and stored it at −4 °C. The sample was dissolved in 70% EtOH at the concentrations (50, 100, and 200 μg/mL) to use for further experiments.
2.2. Cell Culture
The B16F10 cells (Korean Cell Line Bank, Seoul, Korea) purchased for use in this experiment were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 1% penicillin/streptomycin and 10% fetal bovine serum (FBS) and incubated under 37 °C, 5% CO2 humidified conditions.
2.3. Measurement of Cell Viability
Cell cytotoxicity was determined via a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Cells were dispensed into a 96-well plate for cell culture at a concentration of 1.0 × 104 cells/well, and after a day, it was confirmed that the cells were completely attached. Next, PFE diluted with various concentrations was added in triplicate to each well and incubated for 2 days. Following this, the MTT solution was added and after 4 h the medium was subsequently removed. The insoluble MTT formazan crystals were solubilized by DMSO (1 mL). For MTT assays, after cell culture was complete, MTT solution was added to each well at a concentration of 0.5 mg/mL, reacted at 37 °C for 4 h. Then, after wash with PBS buffer, and 1 mL of DMSO was added to each well, followed by using a microplate spectrophotometer (Tecan, Mannedorf, Switzerland) to measure absorbance at 540 nm.
2.4. Analysis of Cell Melanin Content
Cells were dispensed into a 6-well plate for cell culture at a concentration of 5.0 × 104 cells/well, and after a day, it was confirmed that the cells were completely attached. Next, PFE diluted with various concentrations was pretreated in triplicate to each well and incubated for 2 h. The cells were then treated with the native hormone, α-MSH (200 nM), and incubated at 37 ºC for another 72 h. For melanin content analysis, after cell culture was completed, the medium was removed, the color of the cell pellet was observed, and the cells were washed twice with phosphate buffered saline (PBS). Then, 1 mL of 1N NaOH containing 10% DMSO was added directly to each well, the cells were lysed at 80 °C for 1 h at 300 rpm, and the absorbance was measured at 405 nm.
2.5. Analysis of Intracellular Tyrosinase Activity
The equal amount of cells as melanin contents assay was seeded and treated with various concentrations of PFE (50, 100, and 200 μg/mL). For tyrosinase activity test, after centrifugation was carried out to remove the medium, the cell pellets were incubated for 10 min in lysis buffer (radioimmunoprecipitation assay) containing a 1% protease/phosphatase inhibitor cocktail. The cells were scraped and collected in an e-tube and vortexed three times every 10 min to lyse the cells. After centrifugation at 15,000 rpm for 30 min at −8 °C, 80 μL of L-DOPA (2 mg/mL), which was diluted with 0.1 M sodium phosphate buffer, and 20 μL of each quantified protein sample were placed and mixed and then incubated at 37 °C for 2 h. The absorbance of each concentration after a 2 h reaction was measured once at 490 nm to obtain an optical density (O.D) value.
2.6. Preparation of Western Blot Analysis
The equal amount of cells as melanin contents assay was seeded and treated with various concentrations of PFE. Following this, the cell was lysed and centrifuged, and then each fresh lysate (without freezing) was quantified using the bicinchoninic acid assay (BCA) kit. The protein levels were adjusted, which were mixed with the 2 × Laemmli sample buffer to form a Western blot sample. These samples were subsequently denatured at 100 °C for 5 min. Each denatured sample (total volume 20 μL) was loaded on 10% sodium dodecyl sulfate (SDS) polyacrylamide gels for 1 h at 150 V. Following this, using Trans-Blot Turbo (Bio-Rad, Hercules, CA, USA), the gels were blotted onto polyvinylidene difluoride (PVDF) membranes for 7 min. The membranes were incubated using blocking solution, which contained 5% skim milk in Tris-buffered saline containing 0.4% Tween 20 (TBST) buffer for blocking the membranes during 1 h. After that, the membranes were washed every 10 min with TBST buffer over a period of 1 h. The specific primary antibodies (1:1000) were diluted with TBST containing 5% skim milk, which were incubated for 24 h at 4 °C with membrane. Subsequently, the membranes were washed every 10 min for 1 h, and each membrane was hybridized with antimouse IgG or antirabbit IgG conjugated with horseradish peroxidase for 1 h at 25 °C. The membranes were washed six times with TBST, reacted with a chemiluminescence (ECL; Biosesang, Seongnam, Gyeonggi-do, Korea) kit, and then photosensitized to visualize the protein. For quantitative analysis, the amount of protein expression was determined by the ImageJ program (NIH, Bethesda, MD, USA).
2.7. Preparation of Primary Skin Irritation Test
The test on human skin irritation of PFE conducted by the Korean Dermatology Research Institute was conducted as follows. A total of 32 subjects without any existing skin problems participated in the test. Their age range was from 20 to 58 years, while the average age of the subjects was 42 years. All of the subjects’ skins were kept clean and dry, and stabilized at room temperature for at least 30 min. The surface of the skin area to be tested was sterilized by ethanol (70%) before a PFE (1%) patch was placed on the skin and subsequently removed after 24 h. Thirty minutes after removal of the patch, the skin condition of the tested site was observed. The skin response evaluation was based on the guidelines issued by the Personal Care Products Council (PCPC) as well as the International Contact Dermatitis Research Group (ICDRG) (Table 1 and Table 2). All assessments were conducted by qualified professional researchers or dermatologists in this field after obtaining approval from the Ethics committee (Approval number: KDRI-IRB-20484).
2.8. Significance Test through Statistical Analysis
All result values were statistically processed after triplicates and were expressed as the mean ± standard deviation of the values obtained by the experiment. The statistical analysis of this study was verified by a one-way ANOVA test using SigmaStat (ver 2.1), and p < 0.05 was determined to be statistically significant.
3. Results and Discussion
3.1. Effect of PFE on Cell Viabilty
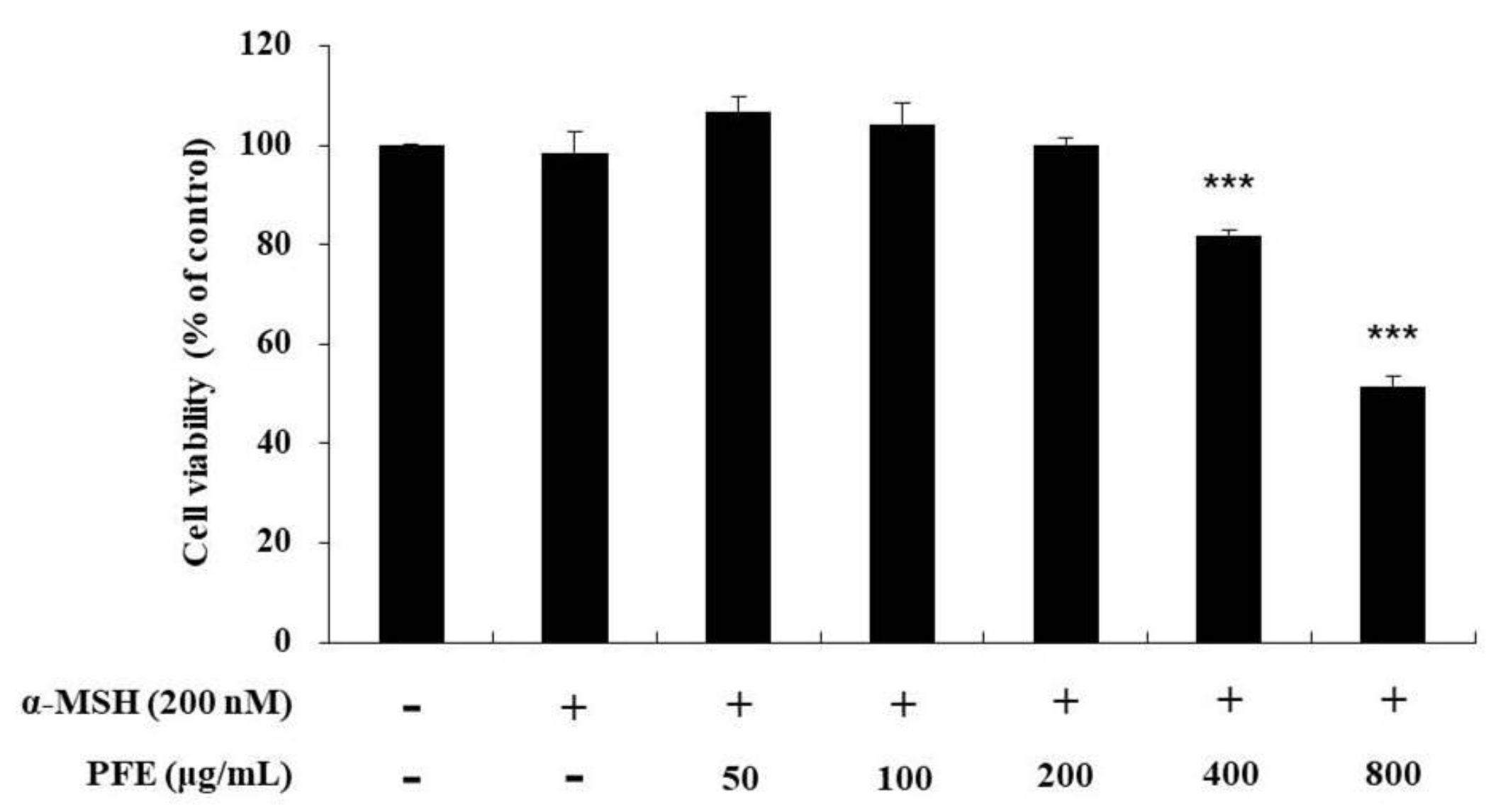
In order to determine the concentration of PFE that can be administered without causing cytotoxicity, the cells were treated with extracts diluted at various concentrations to confirm the survival range of cells. Recent studies indicated that α-MSH was normally used in treatment concentrations ranging from 10 to 1000 nM for B16F10 cells stimulation. Therefore, we treated 200 nM of α-MSH to stimulate B16F10 cells [32,33,34,35,36]. As depicted in Figure 1, the cell viability did not vary significantly at the different concentrations (50, 100, and 200 μg/mL) compared to the untreated control. Therefore, further experiments were conducted using these concentrations.
3.2. Melanin Content and Intracellular Tyrosinase Inhibitory Activity by PFE Treatment
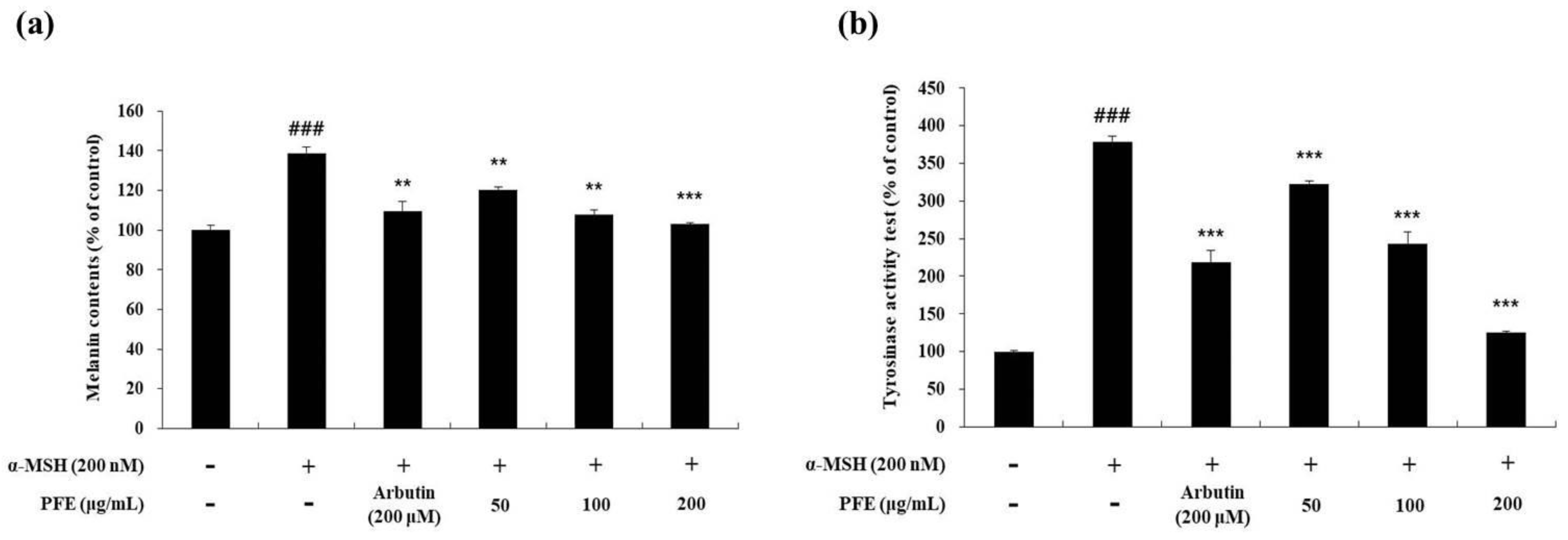
Subsequently, the variation of melanin content and tyrosinase activity due to PFE treatment was measured in cells treated with arbutin (positive control) and α-MSH (negative control). The cells stimulated with α-MSH alone increased the melanin content by 1.4-fold compared to the non-treated cells. However, melanin contents of arbutin (200 μM) and PFE (50, 100, and 200 μg/mL) was reduced by 20.9%, 13.7%, 22.3%, and 26%, respectively, compared to the α-MSH treated alone group (Figure 2A). Additionally, a tyrosinase activity test was performed to determine whether these results were due to a decrease in tyrosinase activity. As observed in Figure 2B, the stimulated cells increased the tyrosinase activity by 3.8-fold compared to the non-treated cells. However, tyrosinase activity of arbutin (200 μM) and PFE (50, 100, and 200 μg/mL) was reduced by 44.2%, 15.0%, 35.9%, and 67.0%, respectively, compared to α-MSH treatment alone. It was confirmed that the activity of tyrosinase in α-MSH-stimulated cells was significantly decreased due to PFE treatment.
3.3. Downregulation of PFE on Melanogenic Enzymes and MITF Expression
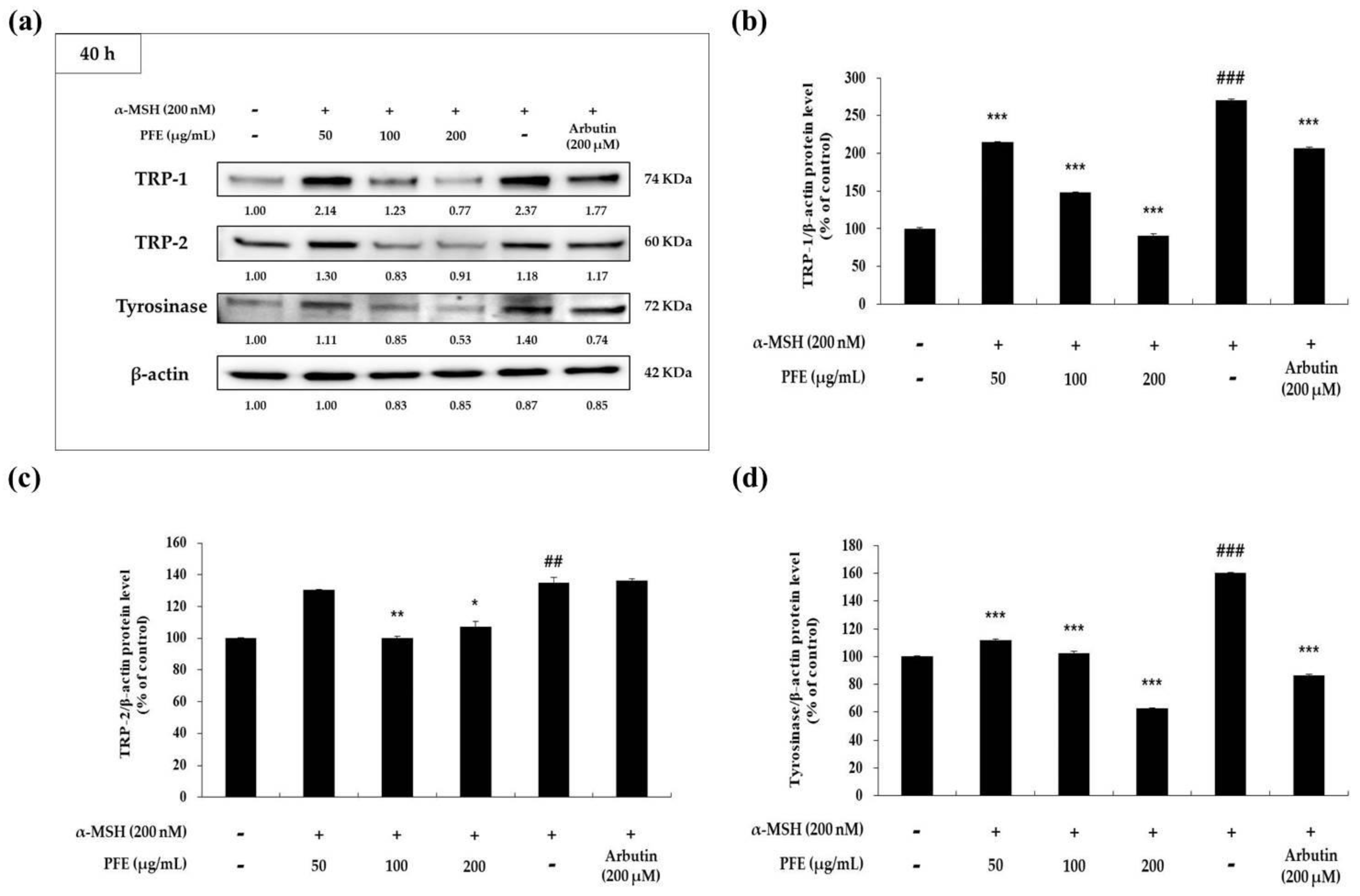
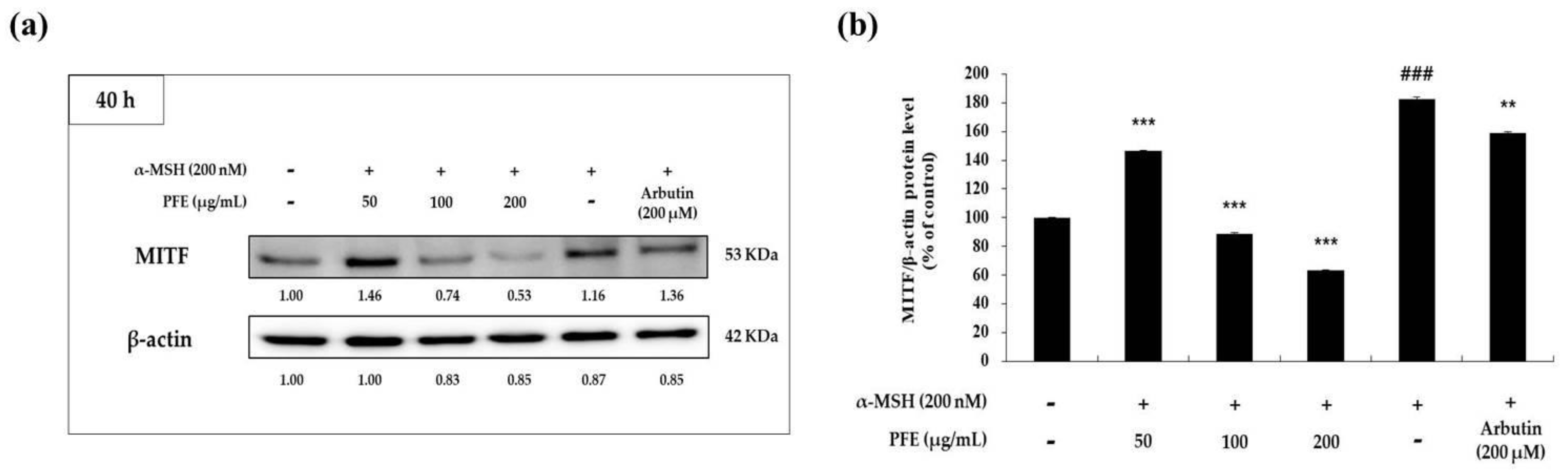
Melanogenic enzymes (TRP-1, -2, tyrosinase) have been reported to increase the production of melanin through each cycle of melanogenesis in the melanosomes. Moreover, these melanogenic enzymes are regulated by MITF expression [11,12,13,14,15,16,17]. Therefore, Western blot experiments were performed to evaluate whether PFE affects melanogenic enzymes expression. As depicted in Figure 3, TRP-1, -2, as well as tyrosinase protein levels were significantly suppressed by PFE treatment compared to those in the negative control (α-MSH-treated) group. As can be seen from the expression level of the melanogenic enzymes in Figure 3, it can be seen that the expression of MITF (Figure 4) was also inhibited depending on the concentration of treatment with PFE. In particular, the protein levels of both melanogenic enzymes and MITF in the presence of 200 μg/mL PFE was lower than that of the untreated control cells. As a result, this indicated that PFE reduced the expression level of melanogenic enzymes and MITF, thereby downregulating melanogenesis in B16F10 cells.
3.4. Effect of PFE on MAPK Phosphorylation
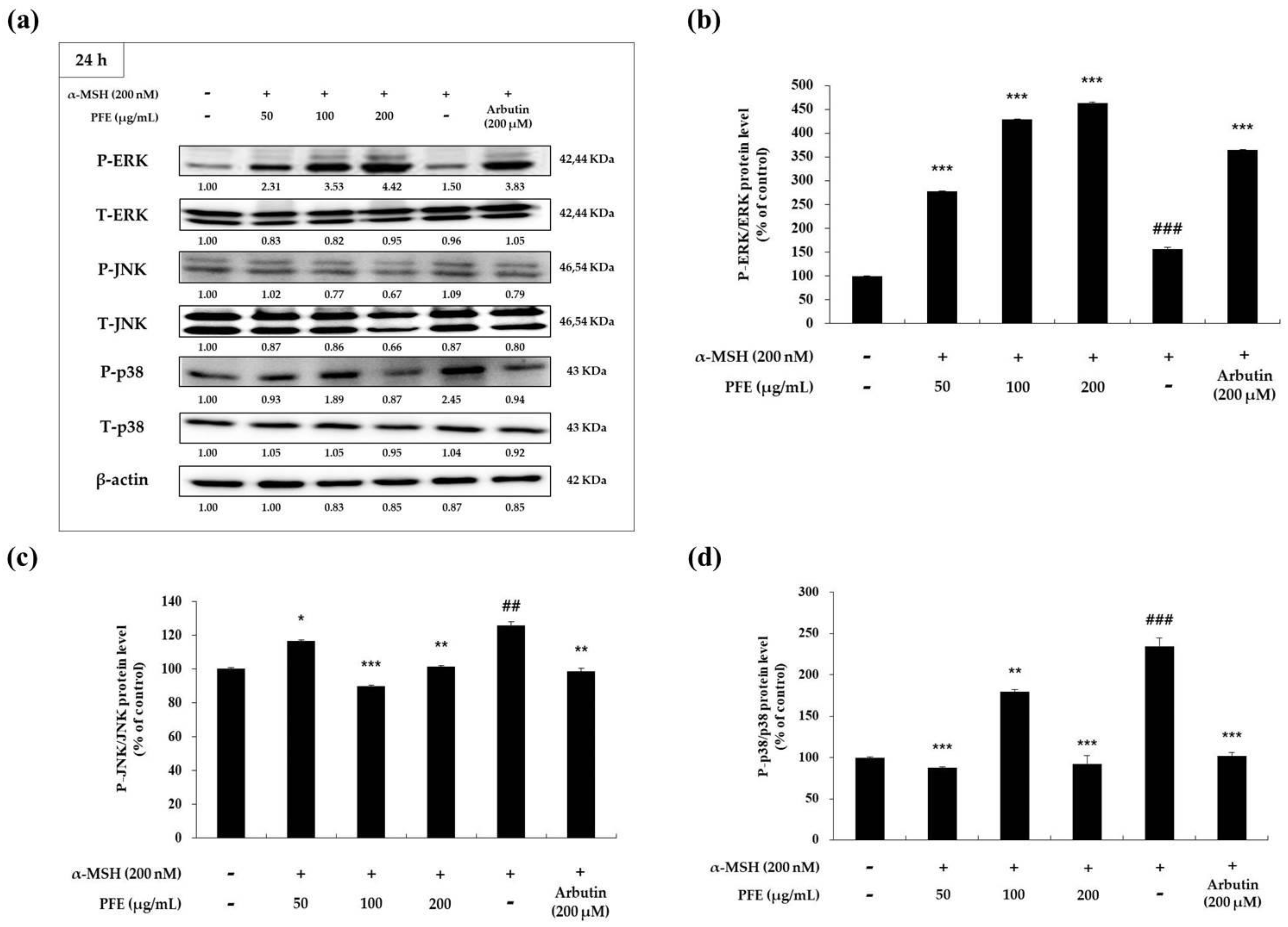
It has been observed in previous research that MITF expression is controlled by phosphorylation of mitogen-activated protein kinases (MAPK). Moreover, it has also been explored that inhibition of p38 and c-Jun N-terminal kinase (JNK), along with the activation of extracellular signal regulated kinase (ERK) phosphorylation, reduced MITF and melanogenic enzyme expression, resulting in the downregulation of melanogenesis [18,19,20,21,22,23]. As a result of confirming whether PFE treatment affects the phosphorylation expression of each MAPK family, PFE promoted the phosphorylation of ERK and inhibited the phosphorylation of JNK and p38 at each treatment concentration when compared to the negative control (Figure 5). In particular, at the highest treatment concentration of 200 μg/mL PFE, a significant change in the expression levels was observed. Therefore, these results suggest that PFE exerts anti-melanogenic effects in the melanoma cells by reducing MITF expression via the MAPK signaling pathway.
3.5. Primary Skin Irritation Test of PFE
The primary skin irritation test is an experimental method that evaluates the direct toxic action of an irritating substance. Therefore, it is used to determine a spontaneously occurring erythema or edema incident to the application of cosmetic products. In order to determine whether PFE is a biocompatibile material for use as a potential cosmetic ingredient, a human primary irritation test was conducted by placing a testing patch on human back skin. As presented in Table 3, PFE did not exhibit any specific indications of irritation—such as erythema, pruritus, or burning—at 24, 48, and 72 h after the patch was removed. These data suggest that PFE can be safely used as a potential cosmetic ingredient in whitening.
4. Conclusions
In conclusion, this study is the first to evaluate the inhibitory effect of PFE on melanin production in B16F10 melanoma cells. Moreover, PFE has been shown to exert anti-melanogenic effects in cells by inhibiting MITF and melanogenic enzymes via the activation or inhibition of phosphorylation of MAPK family in the MAPK signaling pathway. The human skin irritation test results further confirmed that PFE did not have any severe side effects. Consequently, these findings suggest that PFE may be considered a promising candidate for use as a skin whitening ingredient. However, more exhaustive research—such as high-performance liquid chromatography (HPLC)—will be necessary in order to analyze the individual components of PFE that are responsible for its whitening activity.
Author Contributions
Conceptualization, C.-G.H.; methodology, Y.C.C.; validation, C.-G.H.; formal analysis, Y.C.C.; investigation, Y.C.C.; data curation, Y.C.C.; writing—original draft preparation, Y.C.C. and C.-G.H.; writing—review and editing, Y.C.C. and C.-G.H.; contributed reagents/materials/analysis tools, J.N.L. and B.S.K.; project administration, C.-G.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Personal Care Products Council (PCPC) as well as the International Contact Dermatitis Research Group (ICDRG), and approved by the Ethics committee of the Korean Dermatology Research Institute (Approval number: KDRI-IRB-20484 and 8 July 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
This research was supported by the Ministry of Trade, Industry and Energy (MOTIE), Korea Institute for Advancement of Technology (KIAT) through the Encouragement Program for the Industries of Economic Cooperation Region (P0006063).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hearing, V.J. Biogenesis of pigment granules: A sensitive way to regulate melanocyte function. J. Dermatol. Sci. 2005, 37, 3–14. [Google Scholar] [CrossRef]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Yamaguchi, Y.; Batzer, J.; Coelho, S.G.; Zmudzka, B.Z.; Miller, S.A.; Wolber, R.; Beer, J.Z.; Hearing, V.J. Mechanisms of skin tanning in different racial/ethnic groups in response to ultraviolet radiation. J. Investig. Dermatol. 2005, 124, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, N.; Hamdani, M. Plants used to treat skin diseases. Pharmacogn. Rev. 2014, 8, 52–60. [Google Scholar] [CrossRef] [Green Version]
- D’Ischia, M.; Wakamatsu, K.; Cicoira, F.; Mauro, E.D.; Garcia-Borron, J.C.; Commo, S.; Galvan, I.; Ghanem, G.; Kenzo, K.; Meredith, P.; et al. Melanins and melanogenesis: From pigment cells to human health and technological applications. Pigment Cell Melanoma Res. 2015, 28, 520–544. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Otręba, M.; Buszman, E.; Miliński, M.; Wrześniok, D. Hypomelanoses transmitted from generation to generation. Postepy Hig Med Dosw 2014, 68, 1081–1090. [Google Scholar] [CrossRef]
- Otręba, M.; Miliński, M.; Buszman, E.; Wrześniok, D.; Beberok, A. Hereditary hypomelanocytoses: The role of PAX3, SOX10, MITF, SNAI2, KIT, EDN3 and EDNRB genes. Postepy Hig. Med. Dosw. 2013, 67, 1109–1118. [Google Scholar] [CrossRef]
- Miyamura, Y.I.; Coelho, S.G.; Wolber, R.; Miller, S.A.; Wakamatsu, K.; Zmudzka, B.Z.; Ito, S.; Smuda, C.; Passeron, T.; Choi, W. Regulation of human skin pigmentation and responses to ultraviolet radiation. Pigment Cell Res. 2007, 20, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, M. Reactivities of quinone methides versus o-quinones in catecholamine metabolism and eumelanin biosynthesis. Int. J. Mol. Sci. 2016, 17, 1576. [Google Scholar] [CrossRef] [Green Version]
- Hearing, V.J.; Jimenez, M. Mammalian tyrosinase—the critical regulatory control point in melanocyte pigmentation. Int. J. Biochem. 1987, 19, 1141–1147. [Google Scholar] [CrossRef]
- Jiménez-Cervantes, C.; Solano, F.; Kobayashi, T.; Urabe, K.; Hearing, V.J.; Lozano, J.A.; Garcia-Borrón, J.C. A new enzymatic function in the melanogenic pathway. The 5,6-dihydroxyindole-2-carboxylic acid oxidase activity of tyrosinase-related protein-1 (TRP1). J. Biol. Chem. 1994, 269, 17993–18000. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Xie, H.F.; Tang, Y.; Lin, S.Q.; Li, J.M.; Sun, S.M.; Hu, X.L.; Huang, Y.X.; Shi, W.; Jian, D.G. Protein-coupled estrogen receptor enhances melanogenesis via cAMP-protein kinase (PKA) by upregulating microphthalmia-related transcription factor-tyrosinase in melanoma. J. Steroid Biochem. Mol. Biol. 2017, 165, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Bentley, N.J.; Eisen, T.; Goding, C.R. Melanocyte-specific expression of the human tyrosinase promoter: Activation by the microphthalmia gene product and role of the initiator. Mol. Cell. Biol. 1994, 14, 7996–8006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachtenheim, J.; Borovansky, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Jin, S.H.; Kang, H.Y. LPS induces melanogenesis through p38 MAPK activation in human melanocytes. Arch. Dermatol. Res. 2008, 300, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Widlude, H.R.; Fisher, D.E. Microphthalmia-associated transcription factor: A critical regulator of pigment cell development and survival. Oncogene 2003, 22, 3035–3041. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signaling to CREB. Pigment Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.C.; Lin, Y.Y.; Yang, S.Y.; Weng, Y.; Tsai, Y. Antimelanogenic effect of c-phycocyanin through modulation of tyrosinase expression by upregulation of ERK and downregulation of p38 MAPK signaling pathways. J. Biomed. Sci. 2011, 18, 74. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.J.; Choi, B.R.; Lee, E.K.; Kim, S.H.; Yi, H.Y.; Park, H.R.; Song, C.H.; Lee, Y.J.; Ku, S.K. Inhibitory Effect of Dried Pomegranate Concentration Powder on Melanogenesis in B16F10 Melanoma Cells; Involvement of p38 and PKA Signaling Pathways. Int. J. Mol. Sci. 2015, 16, 24219–24242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.S.; Sung, J.H.; Lee, S.K. Antimelanogenesis Activity of Hydrolyzed Ginseng Extract (GINST) via Inhibition of JNK Mitogen-activated Protein Kinase in B16F10 Cells. J. Food Sci. 2016, 81, H2085–H2092. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Hong, E.; Choi, M.; Kim, K.; Lee, S. Antioxidant and Skin Whitening Effects of Rhamnus yoshinoi Extracts. Korean J. Food Sci. Anim. Resour. 2010, 42, 750–754. [Google Scholar]
- Shukla, Y.N.; Lloyd, H.A.; Morton, J.F.; Kapadia, G. Iridoid glycosides and other constituents of Paederia foetida. Phytochemistry 1976, 15, 1989–1990. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, Y.; Han, T.; Zheng, C.; Qin, L. A phytochemical, pharmacological and clinical profile of Paederia foetida and P. scandens. Nat. Prod. Commun. 2014, 9, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanda, S.; Sarethy, I.P.; De, B.; Singh, K. Paederia foetida—A promishing ethnomedicinal tribal plant of Northeast India. J. Forestry Res. 2013, 24, 801–808. [Google Scholar] [CrossRef]
- Pradhan, N.; Parbin, S.; Kausar, C.; Kar, S.; Mawatwal, S.; Das, L.; Deb, M.; Sengupta, D.; Dhiman, R.; Patra, S.K. Paederia foetida induces anticancer activity by modulating chromatin modification enzymes and altering pro-inflammatory cytokine gene expression in human prostate cancer cells. Food Chem. Toxicol. 2019, 130, 161–173. [Google Scholar] [CrossRef]
- Afroz, S.; Alamgir, M.; Khan, M.T.H.; Jabbar, S.; Nahar, N.; Choudhuri, M.S.K. Antidiarrhoeal activity of the ethanol extract of Paederia foetida Linn. (Rubiaceae). J. Ethnopharmacol. 2006, 105, 125–130. [Google Scholar] [CrossRef]
- Kumar, V.; Al-Abbasi, F.A.; Ahmed, D.; Verma, A.; Mujeeb, M.; Anwar, F. Paederia foetida Linn. Paederia foetida Linn. inhibits adjuvant induced arthritis by suppression of PGE(2) and COX-2 expression via nuclear factor-κB. Food Funct. 2015, 6, 1652–1666. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Deb, L.; Tiwari, R.K.; Singh, K.; Ahmad, S. Gastroprotective mechanism of Paederia foetida Linn. (Rubiaceae)—A popular edible plant used by the tribal community of North-East India. BMC Complement. Altern. Med. 2015, 13, 304. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Son, Y.O.; Lee, S.A.; Jeon, Y.M.; Lee, J.C. Quercetin Inhibits α-MSH-stimulated Melanogenesis in B16F10 Melanoma Cells. Phytother. Res. 2011, 25, 1166–1173. [Google Scholar] [CrossRef]
- Seong, Z.K.; Lee, S.Y.; Poudel, A.; Oh, S.R.; Lee, H.K. Constituents of Cryptotaenia japonica Inhibit Melanogenesis via CREB- and MAPK-Associated Signaling Pathways in Murine B16 Melanoma Cells. Molecules 2016, 21, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Won, K.J.; Hwang, D.I.; Park, S.M.; Kim, B.; Lee, H.M. Chemical Composition, Antioxidant and Anti-melanogenic Activities of Essential Oils from Chrysanthemum boreale Makino at Different Harvesting Stages. Chem. Biodivers. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Truong, X.T.; Park, S.H.; Lee, Y.G.; Jeong, H.Y.; Moon, J.H.; Jeon, T.I. Protocatechuic Acid from Pear Inhibits Melanogenesis in Melanoma Cells. Int. J. Mol. Sci. 2017, 18, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Cha, S.B.; Park, M.C.; Park, S.A.; Kim, H.S.; Woo, W.H.; Mun, Y.J. Scopoletin Stimulates Melanogenesis via cAMP/PKA Pathway and Partially p38 Activation. Biol. Pharm. Bull. 2017, 40, 2068–2074. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Variation in cytotoxicity of B16F10 cells to Paederia foetida L. extract (PFE) treatment concentration. Results in the graph are represented as the mean ± SD of at least triplicate experiments. *** p < 0.001 vs non-treated cells.
Figure 1.
Variation in cytotoxicity of B16F10 cells to Paederia foetida L. extract (PFE) treatment concentration. Results in the graph are represented as the mean ± SD of at least triplicate experiments. *** p < 0.001 vs non-treated cells.

Figure 2.
Variation in (a) melanin content and (b) tyrosinase activity of B16F10 cells to PFE treatment concentration. The cells, alpha-Melanocyte-stimulating hormone (α-MSH)-stimulated, were treated with the PFE and arbutin (200 μM). Results in each graph are represented as the mean ± SD of at least triplicate experiments. ### p < 0.001 vs. non-treated. ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.
Figure 2.
Variation in (a) melanin content and (b) tyrosinase activity of B16F10 cells to PFE treatment concentration. The cells, alpha-Melanocyte-stimulating hormone (α-MSH)-stimulated, were treated with the PFE and arbutin (200 μM). Results in each graph are represented as the mean ± SD of at least triplicate experiments. ### p < 0.001 vs. non-treated. ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.

Figure 3.
Variation in melanogenic enzymes expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of melanogenic enzymes and the graph by quantified protein levels of (b) tyrosinase-related protein (TRP)-1/β-actin, (c) TRP-2/β-actin, and (d) tyrosinase/β-actin. Results in each graph are represented as the mean ± SD of at least triplicate experiments. ## p < 0.01, ### p < 0.001 vs. non-treated. * p < 0.05, ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.
Figure 3.
Variation in melanogenic enzymes expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of melanogenic enzymes and the graph by quantified protein levels of (b) tyrosinase-related protein (TRP)-1/β-actin, (c) TRP-2/β-actin, and (d) tyrosinase/β-actin. Results in each graph are represented as the mean ± SD of at least triplicate experiments. ## p < 0.01, ### p < 0.001 vs. non-treated. * p < 0.05, ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.

Figure 4.
Variation in microphthalmia-associated transcription factor (MITF) expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of MITF and (b) the graph by quantified protein levels of MITF. Results in the graph are represented as the mean ± SD of at least triplicate experiments. ### p < 0.001 vs. non-treated. ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.
Figure 4.
Variation in microphthalmia-associated transcription factor (MITF) expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of MITF and (b) the graph by quantified protein levels of MITF. Results in the graph are represented as the mean ± SD of at least triplicate experiments. ### p < 0.001 vs. non-treated. ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone.

Figure 5.
Variation in mitogen-activated protein kinase (MAPK) expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of MAPK and the graph by quantified protein levels of (b) phospho-extracellular signal regulated kinase (P-ERK)/ERK, (c) phosphor-c-Jun N-terminal kinase (P-JNK)/JNK, and (d) P-p38/p38. Results in each graph are represented as the mean ± SD of at least triplicate experiments. ## p < 0.01, ### p < 0.001 vs. non-treated. * p < 0.05, ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone. P: phosphorylated; T: total.
Figure 5.
Variation in mitogen-activated protein kinase (MAPK) expression of B16F10 cells to PFE treatment concentration. The cells, α-MSH-stimulated, were treated with the PFE and arbutin (200 μM) for 40 h. (a) The band of MAPK and the graph by quantified protein levels of (b) phospho-extracellular signal regulated kinase (P-ERK)/ERK, (c) phosphor-c-Jun N-terminal kinase (P-JNK)/JNK, and (d) P-p38/p38. Results in each graph are represented as the mean ± SD of at least triplicate experiments. ## p < 0.01, ### p < 0.001 vs. non-treated. * p < 0.05, ** p < 0.01, *** p < 0.001 vs. α-MSH-treated alone. P: phosphorylated; T: total.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Skin irritation score system.
| Grade | Description of Clinical Observation |
|---|---|
| 0 (−) | No signs of inflammation, normal skin |
| 0.5 (±) | Doubtful or slight reaction |
| 1 (+) | Slight erythema |
| 2 (++) | Moderate erythema, with or without partial edema or papules |
| 3 (+++) | Moderate erythema with diffuse edema |
| 4 (++++) | Intense erythema with diffuse edema and the presence of vesicles |
Table 2.
Skin irritation judgment.
| Range of Response | Inference |
|---|---|
| 0 ≤ * R < 0.02 | No irritation |
| 0.02 ≤ R < 0.25 | Low irritation |
| 0.25 ≤ R < 1 | Slight irritation |
| 1 ≤ R < 2.5 | Moderate irritation |
| 2.5 ≤ R | Severe irritation |
* Irritation Index.
Table 3.
Results of the human skin primary irritation test.
| NO. | PFE (1%) | Blank | ||||
|---|---|---|---|---|---|---|
| Reaction | Reaction | |||||
| 24 h * | 48 h | 72 h | 24 h * | 48 h | 72 h | |
| Grade | 0.5 | 0 | 0 | 0 | 0 | 0 |
| Total number of observations | 96 | 96 | ||||
| Irritation Index | 0.005 | 0 | ||||
| Judgment | No irritation | - | ||||
* Measured 30 min after removal of the patch.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chung, Y.C.; Lee, J.N.; Kim, B.S.; Hyun, C.-G. Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation. Cosmetics 2021, 8, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010022
AMA Style
Chung YC, Lee JN, Kim BS, Hyun C-G. Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation. Cosmetics. 2021; 8(1):22. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010022
Chicago/Turabian StyleChung, You Chul, Jung No Lee, Bong Seok Kim, and Chang-Gu Hyun. 2021. "Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation" Cosmetics 8, no. 1: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.