Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer


Division of Gastroenterology and Hepatology, Medicine, University of Illinois at Chicago, Chicago, IL 60612, USA
*
Author to whom correspondence should be addressed.
Diseases 2019, 7(1), 28; https://0-doi-org.brum.beds.ac.uk/10.3390/diseases7010028
Submission received: 13 January 2019
/
Revised: 16 February 2019
/
Accepted: 6 March 2019
/
Published: 10 March 2019
(This article belongs to the Special Issue Role of Bacterial Infection in Cancer)
{kind=link}
Abstract
:Salmonella not only causes acute infections, but can also cause patients to become chronic “asymptomatic” carriers. Salmonella has been verified as a pathogenic factor that contributes to chronic inflammation and carcinogenesis. This review summarizes the acute and chronic Salmonella infection and describes the current research progress of Salmonella infection contributing to inflammatory bowel disease and cancer. Furthermore, this review explores the underlying biological mechanism of the host signaling pathways manipulated by Salmonella effector molecules. Using experimental animal models, researchers have shown that Salmonella infection is related to host biological processes, such as host cell transformation, stem cell maintenance, and changes of the gut microbiota (dysbiosis). Finally, this review discusses the current challenges and future directions in studying Salmonella infection and its association with human diseases.
1. Introduction
Salmonella is a Gram-negative bacterial genus of the Enterobacteriaceae family with a strong pathogenicity that can cause cross-infection between humans and animals. In the United States, approximately 1.4 million people are infected with Salmonella each year [1]. Salmonella causes fever, diarrhea, gastroenteritis, and sepsis in humans, as well as intestinal damage in both humans and animals. In addition, Salmonella infections are reported to increase the risk of immune-related diseases such as arthritis [2,3] and inflammatory bowel disease (IBD) [4]. Furthermore, chronic infection of Salmonella may lead to gallbladder and colorectal cancer [5,6].
We performed an electronic literature search of papers written in English in the MEDLINE database via PubMed. Searches included combinations of the following terms: Salmonella, infection, microbiome/microbiota, inflammatory bowel disease, inflammation, cancer, nuclear factor kappa B, and stem cells. Papers without clear relevance to the role of Salmonella infection, inflammation or cancer were excluded.
Here, we review the acute and chronic Salmonella infection. We summarize the current research progress of Salmonella infection and its contribution to inflammation and cancer. We also discuss the potential mechanisms of Salmonella infection and its association with various human diseases.
2. Clinic and Epidemiologic Facts of Salmonella Infection
Foodborne diseases cause 1 in 10 persons to fall ill each year and one of every four diarrheal patients has been infected with Salmonella [7]. In the United States, Salmonella is the second most prevalent foodborne infection [8]. In Europe, there were about 94,625 Salmonella infections in 2015 [9]. The preference of “ready-to-eat”, raw or lightly cooked foods is a potential reason for the high number of infections in Western countries. Approximately 2600 serotypes have been described based on the phenotypic identification of the somatic and flagellar antigens of Salmonella [10].
Of the 2600 identified Salmonella serotypes, many cause a range of symptoms in different hosts. Based on its virulence factors, Salmonella can be divided into typhoidal and non-typhoidal Salmonella (NTS) serovars. Salmonella enterica (S. enterica) serotypes Typhi and Paratyphoid typically cause typhoid in humans, which is characterized by systemic infection, fever and gastrointestinal symptoms like diarrhea. In contrast, S. enterica serotype Typhimurium (S. Typhimurium) typically presents as enterocolitis in humans and cattle and is known to cause systemic infection in mice [11].
Typhoidal Salmonella infection typically occurs in under-developed countries due to poor sanitation practices [12], but NTS is prevalent worldwide [12,13]. In fact, there are different clinical manifestations between the typhoidal Salmonella and NTS infection. The manifestations of NTS infection are acute clinical symptoms, such as diarrhea, fever, abdominal pain, nausea, and vomiting. The symptoms are usually self-limiting as the fever usually returns to normal in 72 h and diarrheal symptoms disappear in 3–7 days [14]. Typhoidal Salmonella infection can cause systemic symptoms without the manifestation of intestinal symptoms. Systemic symptoms include a gradual onset of sustained fever, hepatosplenomegaly, and rash [15]. Because antibiotic therapy prolongs fecal excretion of Salmonella [16], it is not recommended to treat an NTS-infected patient with antibiotics unless the patient is immunocompromised. For typhoidal Salmonella-infected patients, fluoroquinolones are usually the first choice to treat the bacteria, and third generation cephalosporins can be an alternative treatment when typhoidal Salmonella is resistant to fluoroquinolones [17]. The acute manifestations are easy to identify and treat by patients and physicians but chronic asymptomatic infection may produce more serious problems, such as IBD and cancer. Thus, typhoidal Salmonella infections can cause patients to become carriers who may not only develop more severe diseases but can spread the pathogen to healthy individuals.
3. Acute Salmonella Infection
Salmonella infections begin with ingestion of the organism from contaminated food or water. Once ingested, Salmonella must overcome the acidity of the stomach. Salmonella can develop an adaptive acid-tolerance response when exposed to gastric pH, promoting its survivability and gastric invasiveness [18,19]. In the intestine, Salmonella can invade the epithelium through three distinct routes: by adhering and entering into M-cells or epithelial cells, or by infecting mononuclear phagocytes that sample the gut lumen [20].
There are many virulence factors that are involved in infection. Bacteria surface moieties, toxins, and effector proteins all play roles in invasion. Most importantly, these components can modify essential host cell-signaling pathways related to both acute and chronic infections [21,22,23]. Salmonella’s ability to invade host cells is regulated by Salmonella pathogenicity islands (SPI), which are clusters of genes that are responsible for specific virulence phenotypes and are acquired through horizontal gene transfer. SPI-1 and SPI-2 code for variants of the type 3 secretion system (T3SS) [24]. The T3SS acts as a ‘molecular syringe’ comprised of two ring-like structures at its base that span the bacterial inner and outer membrane and a needle-like structure protruding on the outside of the bacteria [25,26]. The T3SS is primarily responsible for secretion of bacterial effector proteins needed for the invasion of intestinal epithelial cells that result in intestinal secretory and inflammatory responses as well as translocating proteins for intracellular survival, replication and the establishment of systemic infection beyond the intestinal niche [27].
When bacteria interacts with the host cell, a signal is initiated inside the bacterium causing the translocators SipB and SipC to form a core complex on the host cell’s plasma membrane [21]. Effector proteins SipA and SipC will directly interact with host-cell actin, inducing cytoskeletal re-arrangements [28]. The effectors SopE, SopE2, and SopB mediate this process by activating Rho family GTPases resulting in the formation of highly organized actin structures that cause the host cell membrane to ruffle around and engulf the bacterium into a phagosome or Salmonella-containing vacuole (SCV) [29]. Once engulfed, the membrane shuffling is terminated by the bacterial effector protein SptP, causing the actin cytoskeleton to return to its original state [30]. SopB also activates AKT for efficient formation of the SCV and excludes Rab proteins from the SCV to delay lysosomal degradation and epithelial cell apoptosis [31,32]. SopB, SipA, and SopE promote acute intestinal inflammation and fluid secretion by disrupting tight junctions between epithelial cells and challenging the inositol phosphate signaling pathways that prevent adequate chloride secretion [21,33].
AvrA is an interesting Salmonella protein that plays a unique role compared to other effector molecules. We have demonstrated that AvrA inhibits the NF-κB signaling pathway and activates β–catenin signaling [34,35,36,37]. These host-bacterial interactions lead to suppression of innate immunity, inflammation, and apoptosis during infection [21,37,38]. Infection with Salmonella-expressing AvrA has been shown to promote intestinal permeability [39,40], activate the Wnt/β–catenin pathway, and subsequently increase proliferation [41,42,43]. In fact, AvrA plays a role in a few other pathways including mTOR, oxidative phosphorylation, and MAPK [44]. AvrA also modulates the Salmonella survival niche within macrophages by preventing apoptosis and bacterial propagation [45]. Bacterial effectors like AvrA provide opportunities for pathogens to “make peace” with the host and survive for extensive periods of time within host cells.
4. Persistent/Chronic Salmonella Infection in Mice and Men
Two to five percent of patients with acute Salmonella typhoid infections fail to completely clear the bacteria from the body, leading to chronic infection [46]. The course of chronic Salmonella infection has three distinct periods. A period of convalescence when carriers shed the bacteria for 3 weeks to 2 months post-infection; a period of temperance in which carriers sheds the bacteria for 3–12 months; and finally the chronic period where the organism is shed for more than a year [47]. In chronic infections, Salmonella can also use the gallbladder as a niche. It can invade the gallbladder directly from the liver or invade via the biliary tract directly from the small intestine or from system infection. [48]. It is presumed that once settled in the gallbladder, gallstone biofilms of Salmonella lead to reinfection of the intestine with subsequent transmission to new hosts via fecal shedding [49].
Studies have shown that IL-22 and IFN-γ-mediated immunity is vital for Salmonella eradication from cellular reservoirs [50]. An ineffective immune response or microbiota disruption can be implicated in the host’s inability to clear the organism [50]. Once persistence has been established, it is commonly detected by high anti-Vi titers [48]. The Vi antigen of Salmonella is coded by SPI-7 and plays a role in evading infection by masking the detection of LPS by TLR4 [50]. Therefore, the human carrier state may be associated with impaired immune function.
The most common in vivo method to study Salmonella infection is to use mouse models. S. Typhimurium instead of S. Typhi is commonly used in mouse models because the disease phenotype is similar to human typhoid as well as the fact that mice are unable to be infected by S. Typhi [51]. However, in mice, S. Typhimurium mimics the acute phase of typhoid fever. Several chronically infected mice models have been studied. We created a mouse model with persistent Salmonella infection lasting 27 weeks and investigated the physiological effects as well as the role of AvrA. We found that AvrA suppressed intestinal inflammation and inhibited the secretion of cytokines IL-12, IFN-γ, and TNF-α. AvrA expression in Salmonella enhanced its invasiveness. Liver abscesses and Salmonella translocation to the gallbladder were observed [52]. Further studies have emphasized the importance of AvrA in intestinal inflammation, bacterial translocation, and chronic infection associated with colon cancer in vivo [52,53,54,55].
C57BL/6 is a popular inbred mouse strain used in most laboratories. However, these mice express a functionally defective variant of Nramp1[56]. The Nramp1 gene encodes an ion transporter that is expressed in macrophage and dendritic cells and is a critical host factor in defense against intracellular bacteria [57]. Thus, this variant causes C57BL/6 mice to be innately susceptible to virulent strains of Salmonella but succumb to infection in 7 days. Conversely, 129SvJ mice express a wild-type Nramp1 (Nramp1wt/wt) variant that is functionally active and as a result are capable of developing a persistent infection with virulent Salmonella strains [56]. It has been shown that if infected with virulent Salmonella, the bacteria is found in macrophages and lymph nodes for up to a year in Nramp1wt/wt mice or their F1 hybrids when crossed with C57BL/6 mice [58].
Not much is known regarding the mechanisms of persistent Salmonella infection. A negative screen isolating factor required for Salmonella persistence performed by Lawley and colleagues revealed that SPI-1 was necessary for sustaining persistent infection for at least 1 month post infection [59]. It is also likely that chronic infection is maintained by the re-invasion of S. Typhimurium into epithelial tissues after its expulsion from dying cells [60,61]. T3SS effector Sse1 is also required for long-term systemic infection in mice [62]. It is secreted by Salmonella from the SCV into the host cytosol and plays a role in regulating cell migration in macrophage and dendritic cells by interfering with their ability to adhere, migrate, and communicate with other parts of the immune system [62,63]. In addition, effectors ShdA and MisL, which bind to fibronectin as well as other surface structures encoded by operons Ipf, bcf, and sth, contribute to persistence and fecal shedding of live Salmonella [64,65,66,67,68,69]. In addition, effectors that are part of SPI-2 are required for chronically infected mice to transmit S. Typhimurium to non-infected cage mates [66,70].
Lastly, a mechanism that host cells use to minimize microbial invasion is by producing reactive oxygen species (ROS) and reactive nitrogen species (RNS) [71]. In particular, neutrophils produce myeloperoxidase that can directly bind to the cell surface of Salmonella allowing for targeted killing of the organism via ROS [72]. This mechanism controls the growth of Salmonella at sites of inflammation, but when used in conjunction with RNS, can control the overall spread of infection [73]. However, studies have indicated that mice lacking the ability to produce nitric oxide synthase succumb to Salmonella infection and are not chronically infected [74,75]. However, chronic inflammation can occur in vivo due to T3SS genes sodC1 and hmp, which code for enzymes that detoxify superoxide and nitric acid respectively [70,76,77,78].
These characteristics help Salmonella to survive long-term in host cells and result in a persistent or chronic infection. Patients with chronic Salmonella infections do not display symptoms, but can infect other people because the bacteria can persist in the stool [79].
5. Salmonella Infection and Chronic Inflammatory Diseases
The intestinal mucosa contains more than 80% of activated B cells, and the intestine is the main antibody-producing tissue of the body [80]. Thus, infections in the intestine often cause gastrointestinal disorders and impaired immune functions, all of which result in intestinal or systemic autoimmune diseases like IBD (inflammatory bowel disease) [4] and arthritis [2,3]. The pathogenesis of IBD involves the following three aspects: (1) impaired intestinal mucosal barrier function, (2) gut microbiota changes, and (3) immune regulation disorder [81].
Salmonella must compete with normal gut microbiota for nutrients and intestinal epithelial attachment sites. This competitive relationship is known as colonization resistance of gut microbiota. Some studies have shown that Salmonella can alter colonization resistance in the intestine of mice, destroy normal gut microbiota, and cause colitis [82,83]. Salmonella infection increases susceptibility to intestinal inflammation and contributes to IBD [84]. However, the disorder of the host immune system caused by infection is the key factor for Salmonella-induced IBD. When the intestinal mucosal barrier is destroyed upon Salmonella infection, pro-inflammatory pathways are activated. The binding of TLRs and NOD proteins to bacterial surface moieties and toxins activates the NF-κB pathway leading to the production of cytokines and chemokines. The basolateral secretion of IL-8 recruits neutrophils and granulocytes into the subepithelial layer. Macrophages are also activated through their TLRs in the subepithelial region and induce the production of IL-1, IL-6, and IL-23, all of which drive the differentiation of Th17 cells which further induce an inflammatory state, recruiting and activating neutrophils to clear out the bacterial infection. In addition, IL-8 and IL-12 secreted from activated macrophages drive IFN-γ-dependent production of antigen specific Th1 cells [20,22,85].
In addition to promoting bacterial entry, many effector proteins encoded by SPI-1 activate inflammatory signaling. The activation of Rho family GTPases by SopE and SopB results in the activation of MAPK, ERK, JNK, and p38 signaling and subsequent induction of NF-κB encouraging a proinflammatory state [22,86]. Salmonella effector protein AvrA stabilizes the expression of tight junction proteins (e.g., ZO-1) and plays a role in reducing inflammation in Salmonella-induced colitis in in vivo models [39,40]. AvrA can inhibit epithelial cell apoptosis by regulating c-Jun N-terminal kinase to reduce epithelial cell damage [78]. In addition, Salmonella targets the tight junction protein claudin-2 to facilitate bacterial invasion by activating the EGFR pathway [87]. The S. Typhimurium lpf, bcf, stb, stc, std, and sth fimbriae operons are known to be required for intestinal persistence in mice [33]. The balance between protection and injury could be an underlying mechanism of Salmonella chronic infection.
Chronic Salmonella infection also leads to intestinal fibrosis. Mouse studies have shown that fibrosis appears at day 7, peaking at day 21, and persists to day 70 after Salmonella infection [88]. Salmonella-induced fibrosis in mice is a practical way to build an animal model needed to study intestinal fibrosis [89,90]. The underlying mechanism of intestinal fibrosis caused by Salmonella is linked to MyD88 and Cox-2 signaling. MyD88 (−/−) mice produce less collagen and have fewer fibroblasts in the submucosa compared with WT mice. The Cox-2 inhibitor rofecoxib could reverse the process of fibrogenesis in WT mice [91].
6. Salmonella Infection Increases Risk of Gallbladder Cancer and Colon Cancer
The link between viral infections and their roles in inducing cancers such as cervical and liver cancer has been confirmed, but the role in which bacteria contributes to tumorigenesis is poorly understood. The most widely accepted example of this is that Helicobacter pylori infections can contribute to gastric cancer [92]. Chronic inflammation of the gastric mucosa by Helicobacter pylori can activate the Wnt/β-catenin pathway stimulating carcinogenesis [93]. Research has shown that chronic S. Typhoid and S. Paratyphoid carriers have higher risk for cancer of the gallbladder, pancreas, colon and lung [94]. In subsequent studies, Salmonella was verified as a pathogen that can induce both gallbladder [5] and colon cancer [6]. Gallbladder cancer is the sixth most common gastrointestinal cancer and the annual incidence of gallbladder cancer is about 2 per 100,000 worldwide [95,96]. There are several common genes mutated in gallbladder cancer, such as KRAS [97], TP53 [98], and C-ERB-B2 [99].
Salmonella Typhi infection not only causes chronic inflammation and damage of the mucosa, but also produces toxins such as cytolethal distending toxins (CDT), which contribute to DNA damage and induce cell-cycle arrest [100]. CDTs are heterotrimeric toxins comprised of CdtA, CdtB, and CdtC subunits. The CdtB subunit can activate DNAse I activity, while CdtA and CdtC contribute to the binding of the holotoxin to the plasma membrane of host cells [101]. CdtB activity relies on the expression of two genes, pertussis-like toxin A (pltA) and pertussis-like toxin B (pltB), which can form a CdtB–PltA–PltB tripartite complex that induces DNA damage and cell-cycle arrest, thereby inducing carcinogenesis in the gallbladder [102,103]. It has been shown that both S. Typhi and non-typhoidal Salmonella both contribute to gallbladder carcinogenesis. Reports have indicated that TP53 mutations and Salmonella chronic infection models in gallbladder cancer are analogous to the pathogenic role of Helicobacter pylori in gastric cancer [103]. Butin-Israeli et al stated that neutrophil-derived miR-23a and miR-155 downregulate LB1 and RAD51 leading to increased inflammation and genomic instability [104]. RAD51 plays a key role in DNA homologous recombination and can be downregulated by p53 [105,106]. In our previous study, we found that Salmonella Typhimurium infection increases p53 acetylation via AvrA in intestinal epithelial cells. P53 acetylation was able to induce cell cycle arrest at G0/G1 and decrease inflammation. Though AvrA increases p53 acetylation, total p53 levels were reduced [107]. We assume that decreased p53 leads to upregulation of RAD51, thereby promoting the DNA damage repair process. This process may confer a survival advantage to infected cells by increasing the activation of the DNA repair system allowing the cells to eliminate DNA mutations.
Recently, a report indicated that severe Salmonella infection contributes to increased risk of human colon cancer with tumors developing mainly in the ascending colon and the transverse colon [6]. Salmonella chronic infection and subsequent immune disorders are potential risks of colon cancer due to increased risk of salmonellosis in colon cancer patients with IBD [6]. IBD is an independent risk factor of colon cancer with about 20% of IBD patients developing colon cancer in 30 years [108]. Salmonella infection is a potential factor contributing to the transformation of host cells from a colitic to cancerous state through the activation of its T3SS effectors. AvrA’s deubiquitinase activity can block degradation of IκBα and β-catenin and suppress the NF-κB pathway, thereby inhibiting the host inflammatory responses [34]. It is well known that the NF-κB and Wnt/β-catenin pathways are closely related to the occurrence and development of cancer [109,110]. The expression of AvrA is reported to be higher in cancer adjacent tissues than in normal human colon and cancer tissues [53]. Therefore, Salmonella AvrA may lead to colon cancer by regulating the NF-κB pathway and Wnt/β-catenin. In fact, a further study showed that AvrA activates the Wnt/β-catenin signaling pathway by preventing β-catenin ubiquitination and degradation, resulting in the upregulation of Myc and cyclin D1 expression and the promotion of colon tumorigenesis [54]. The STAT3 signaling pathway is a key player both in IBD and colitis-associated cancer [111] as it can inhibit inflammation and promote proliferation. Furthermore, we have found that AvrA can activate the STAT3 signaling pathway by upregulating IL-6 [112]. Thereby, secretion of AvrA by Salmonella can contribute to colitis-associated cancer by upregulating STAT3, NF-κB, and Wnt/β-catenin signaling pathways.
Other bacterial effectors could also play a role in carcinogenesis. As previously stated, SopB is a T3SS effector that induces sustained activation of Akt/protein kinase B, a pro-survival kinas; activates the Akt signaling pathway; and subsequently protects epithelial cells from apoptosis [113]. Based on this report, the anti-apoptotic role of SopB could be an underlying mechanism of Salmonella-associated colon cancer.
7. Mechanisms: Transformation, Stem Cells, Microbiota
Salmonella infection plays an important role in cellular phenomena, such as host cell transformation, stem cell growth, inflammation, and gut microbiota regulation. Salmonella can induce cellular transformation by activating the Wnt/β-catenin signaling pathway. Studies report that Salmonella can induce follicular-associated epithelial cell transformation into M cells by epithelial-mesenchymal transition (EMT), thereby promoting its colonization and invasion [114]. GSK3β is a key complex in Wnt/β-catenin signaling, which interacts with β-catenin, causing its phosphorylation and degradation. SopB can inhibit GSK3 by activating Akt, [113] which phosphorylates β-catenin increasing its transcriptional activity and ultimately activating the Wnt/β-catenin pathway [115]. Wnt/β-catenin activation increases the expression of EMT transcriptional activators (snail, slug, and twist) and down-regulates the expression of the tumor suppressor E-cadherin [116]. T3SS is also important for neoplastic transformation of epithelial cells. It has been shown that Apc+/min mice (with c-Myc overexpression) [117] infected with WT S. Typhimurium developed cancer in their colon, but Apc+/min mice infected with ΔprgH mutant Salmonella (lacking functional T3SS) did not [118]. Mouse embryonic fibroblasts (MEFs) with genetic susceptibility can transform after WT Salmonella infection because effectors of the T3SS increase AKT and MAP kinase activities, whereas mutant Salmonella lacking effectors SopB, SopE, SopE2, and SptP cannot activate the AKT and MAP kinase [119]. The transformation to cancerous cells persists even after Salmonella has been eradicated from the host. Studies have also shown that during Salmonella infection, the organism can modify the transcriptome of the host cell and induce epigenetic changes [118]. In our previous studies, we demonstrated that Salmonella protein AvrA activates the Wnt/β-catenin pathway, which is essential for the maintenance of the intestinal stem cell niche [43]. Salmonella infection can also regulate the intestinal stem cell markers, Lgr5 and Bmi1 [120].
The gut microbiota is a stable and closed system because of its colonization resistance. In most cases, foreign microbes are unable to colonize a healthy gut and disrupt the gut microbiota [121]. Salmonella is capable of disrupting the gut flora but once the pathogen has been cleared from the gut, the microbiota population can revert back to normal [82]. Salmonella can colonize the gut microbiota through multiple mechanisms. Upon infection, Salmonella can stimulate the C-type lectin RegIIIβ which can destroy indigenous bacteria, such as E. coli and Yersinia spp., resulting in dysbiosis [122,123]. Salmonella can utilize ethanolamine as a nutrient to proliferate and colonize via tetrathionate respiration inside of the inflamed intestine [124]. Salmonella can also compete with indigenous microbiota for intestinal epithelial attachment sites and nutrients thereby disrupting gut microbiota.
In summary, Salmonella infection may induce carcinogenesis through manipulating multiple factors (Figure 1). Salmonella can cause persistent intestinal infection, gut microbiota imbalance and chronic inflammation, which further induces DNA damage resulting in chromosome instability or epigenetic modification. Cancer-related signaling pathways are activated by Salmonella effector proteins. During chronic infection, Salmonella activates the Wnt/β-catenin signaling pathway, leading to host cell transformation. Leaky gut, microbiota imbalance, and inflammation are induced by bacterial proteins and contribute to the development of cancer.
8. Conclusion and Future Directions
Salmonella is a leading bacterial cause of acute gastroenteritis and constitutes a huge health burden in both developing and developed countries. Symptoms of acute infection (e.g., diarrhea, fever, abdominal pain, nausea, and vomiting) are easy to identify and treat using antibiotics or through natural clearance of the organism. However, Salmonella infection can exist in host cells persistently causing patients to chronically carry the pathogen resulting in further spread of the bacteria. Persistent infection can also lead to the development of other severe diseases such as IBD and cancer. Lastly, chronic Salmonella infection plays a role in a number of biological processes, such as stem cell maintenance, host cell transformation, and gut dysbiosis. The mechanism by which Salmonella leads to colitis-associated colon cancer include (1) impaired intestinal mucosal barrier function by acute infection; (2) the effectors of the T3SS activating essential host cell pathways causing immune regulation disorders and finally; (3) Salmonella-associated dysbiosis. Salmonella can act as an extremely pathogenic organism and cause extensive damage to host cells contributing to the onset of severe diseases.
Salmonella has benign properties that can be used to diagnose or treat cancer. It has been reported that live attenuated bacteria can destroy cancer cells in vitro and in vivo [125,126]. Specifically, recombinant attenuated Salmonella vaccines (RASV) have been developed to treat neuroblastoma [127]. Salmonella can be used as a vector to deliver chemotherapeutic drugs to cancer cells and eradicate them with precision. However, much research is needed before these practices can be used as sufficient cancer therapies as the safety of this treatment strategy has yet to be evaluated.
Salmonella-associated cancer involves the interplay of multiple elements, such as environmental factors, host genetic background, the gut microbiome, and host cell immunity. It should be noted that Salmonella infection and its chronic effects are not the same as chemically-induced biological events. Salmonella infection may not act in a dose-dependent manner. In the event a small number of bacteria managed to survive in the host, the opportunity could still arise for the organism to multiply and translocate to other organs. Studies using human samples and mouse models will help us further understand the relationship between Salmonella infections and the risk of colon cancer.
Author Contributions
J.S. designed and directed the project. L.Z., S.G. and J.S. contributed to the writing of the manuscript.
Funding
We would like to acknowledge the support from UIC Cancer Center, the NIDDK grant R01 DK105118 and NIDDK R01 R01DK114126 to Jun Sun.
Acknowledgments
We would like to thank Hongyu Zhang from the First Affiliated Hospital of Chongqing Medical University, China, for the helpful discussion on colon cancer and infection and Danika Bakke for helping with proofreading.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Rohekar, S.; Tsui, F.W.; Tsui, H.W.; Xi, N.; Riarh, R.; Bilotta, R.; Inman, R.D. Symptomatic acute reactive arthritis after an outbreak of salmonella. J. Rheumatol. 2008, 35, 1599–1602. [Google Scholar] [PubMed]
- Li, S.; Yu, Y.; Yue, Y.; Zhang, Z.; Su, K. Microbial Infection and Rheumatoid Arthritis. J. Clin. Cell. Immunol. 2013, 4. [Google Scholar] [CrossRef]
- Axelrad, J.E.; Olen, O.; Askling, J.; Lebwohl, B.; Khalili, H.; Sachs, M.C.; Ludvigsson, J.F. Gastrointestinal Infection Increases Odds of Inflammatory Bowel Disease in a Nationwide Case-Control Study. Clin. Gastroenterol. Hepatol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Koshiol, J.; Wozniak, A.; Cook, P.; Adaniel, C.; Acevedo, J.; Azocar, L.; Hsing, A.W.; Roa, J.C.; Pasetti, M.F.; Miquel, J.F.; et al. Salmonella enterica serovar Typhi and gallbladder cancer: A case-control study and meta-analysis. Cancer Med. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Mughini-Gras, L.; Schaapveld, M.; Kramers, J.; Mooij, S.; Neefjes-Borst, E.A.; Pelt, W.V.; Neefjes, J. Increased colon cancer risk after severe Salmonella infection. PLoS ONE 2018, 13, e0189721. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2013. EFSA J. 2015, 13, 3991. [Google Scholar] [CrossRef] [Green Version]
- Grimont, P.A.; Weill, F.-X. Antigenic formulae of the Salmonella serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–166. [Google Scholar]
- Grassl, G.A.; Finlay, B.B. Pathogenesis of enteric Salmonella infections. Curr. Opin. Gastroenterol. 2008, 24, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, E.; Schwartz, E. Enteric fever: A travel medicine oriented view. Curr. Opin. Infect. Dis. 2010, 23, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease “Burden of Illness” Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Wang, Y.; Su, L.H.; Chiu, C.H. Nontyphoid salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Stuart, B.M.; Pullen, R.L. Typhoid: Clinical analysis of three hundred and sixty cases. Arch. Intern. Med. 1946, 78, 629–661. [Google Scholar] [CrossRef]
- Aserkoff, B.; Bennett, J.V. Effect of antibiotic therapy in acute salmonellosis on the fecal excretion of salmonellae. N. Engl. J. Med. 1969, 281, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Ordóñez, A.; Begley, M.; Prieto, M.; Messens, W.; López, M.; Bernardo, A.; Hill, C. Salmonella spp. survival strategies within the host gastrointestinal tract. Microbiology 2011, 157, 3268–3281. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Ordóñez, A.; Prieto, M.; Bernardo, A.; Hill, C.; López, M. The acid tolerance response of Salmonella spp.: An adaptive strategy to survive in stressful environments prevailing in foods and the host. Food Res. Int. 2012, 45, 482–492. [Google Scholar] [CrossRef]
- Murphy, K.; Travers, P.; Walport, M. Janeway’s Immunology; Garland Science: New York, NY, USA, 2012. [Google Scholar]
- Ramos-Morales, F. Impact of Salmonella enterica type III secretion system effectors on the eukaryotic host cell. ISRN Cell Biol. 2012, 2012, 1–36. [Google Scholar] [CrossRef]
- Patel, S.; McCormick, B.A. Mucosal Inflammatory Response to Salmonella typhimurium Infection. Front. Immunol. 2014, 5, 311. [Google Scholar] [CrossRef] [PubMed]
- Coburn, B.; Sekirov, I.; Finlay, B.B. Type III secretion systems and disease. Clin. Microbiol. Rev. 2007, 20, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, V.; Hensel, M. Cellular microbiology of intracellular Salmonella enterica: Functions of the type III secretion system encoded by Salmonella pathogenicity Island 2. Cell. Mol. Life Sci. CMLS 2004, 61, 2812–2826. [Google Scholar] [CrossRef] [PubMed]
- Hueck, C.J. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Biol. Rev. 1998, 62, 379–433. [Google Scholar] [PubMed]
- Galán, J.E.; Wolf-Watz, H. Protein delivery into eukaryotic cells by type III secretion machines. Nature 2006, 444, 567. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Jain, S.K. Role of antigens and virulence factors of Salmonella enterica serovar Typhi in its pathogenesis. Microbiol. Res. 2012, 167, 199–210. [Google Scholar] [CrossRef] [PubMed]
- McGhie, E.J.; Hayward, R.D.; Koronakis, V. Cooperation between actin-binding proteins of invasive Salmonella: SipA potentiates SipC nucleation and bundling of actin. EMBO J. 2001, 20, 2131–2139. [Google Scholar] [CrossRef] [Green Version]
- Hardt, W.-D.; Chen, L.-M.; Schuebel, K.E.; Bustelo, X.R.; Galán, J.E.S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 1998, 93, 815–826. [Google Scholar] [CrossRef]
- Fu, Y.; Galan, J.E. A salmonella protein antagonizes Rac-1 and Cdc42 to mediate host-cell recovery after bacterial invasion. Nature 1999, 401, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Terebiznik, M.R.; Vieira, O.V.; Marcus, S.L.; Slade, A.; Yip, C.M.; Trimble, W.S.; Meyer, T.; Finlay, B.B.; Grinstein, S. Elimination of host cell PtdIns (4, 5) P 2 by bacterial SigD promotes membrane fission during invasion by Salmonella. Nat. Cell Biol. 2002, 4, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Bakowski, M.A.; Braun, V.; Lam, G.Y.; Yeung, T.; Do Heo, W.; Meyer, T.; Finlay, B.B.; Grinstein, S.; Brumell, J.H. The phosphoinositide phosphatase SopB manipulates membrane surface charge and trafficking of the Salmonella-containing vacuole. Cell Host Microbe 2010, 7, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Boyle, E.C.; Brown, N.F.; Finlay, B.B. Salmonella enterica serovar Typhimurium effectors SopB, SopE, SopE2 and SipA disrupt tight junction structure and function. Cell. Microbiol. 2006, 8, 1946–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.; Petrof, E.O.; Boone, D.; Claud, E.C.; Sun, J. Salmonella effector AvrA regulation of colonic epithelial cell inflammation by deubiquitination. Am. J. Pathol. 2007, 171, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. Pathogenic Bacterial Proteins and their Anti-Inflammatory Effects in the Eukaryotic Host. Antiinflamm. Antiallergy Agents Med. Chem. 2009, 8, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Hobert, M.E.; Rao, A.S.; Neish, A.S.; Madara, J.L. Bacterial activation of beta-catenin signaling in human epithelia. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G220–G227. [Google Scholar] [CrossRef] [PubMed]
- Collier-Hyams, L.S.; Zeng, H.; Sun, J.; Tomlinson, A.D.; Bao, Z.Q.; Chen, H.; Madara, J.L.; Orth, K.; Neish, A.S. Cutting edge: Salmonella AvrA effector inhibits the key proinflammatory, anti-apoptotic NF-kappa B pathway. J. Immunol. 2002, 169, 2846–2850. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.M.; Wu, H.; Wentworth, C.; Luo, L.; Collier-Hyams, L.; Neish, A.S. Salmonella AvrA Coordinates Suppression of Host Immune and Apoptotic Defenses via JNK Pathway Blockade. Cell Host Microbe 2008, 3, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, A.P.; Petrof, E.O.; Kuppireddi, S.; Zhao, Y.; Xia, Y.; Claud, E.C.; Sun, J. Salmonella type III effector AvrA stabilizes cell tight junctions to inhibit inflammation in intestinal epithelial cells. PLoS ONE 2008, 3, e2369. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhang, Y.G.; Xia, Y.; Xu, X.; Jiao, X.; Sun, J. Salmonella enteritidis Effector AvrA Stabilizes Intestinal Tight Junctions via the JNK Pathway. J. Biol. Chem. 2016, 291, 26837–26849. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, S.; Xia, Y.; Li, X.E.; Xia, Y.; Zhou, Z.D.; Sun, J. Wingless homolog Wnt11 suppresses bacterial invasion and inflammation in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G992–G1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Lu, R.; Wu, S.; Zhang, Y.G.; Xia, Y.; Sartor, R.B.; Sun, J. Wnt2 inhibits enteric bacterial-induced inflammation in intestinal epithelial cells. Inflamm. Bowel Dis. 2012, 18, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Lu, R.; Wu, S.; Sun, J. Salmonella regulation of intestinal stem cells through the Wnt/beta-catenin pathway. FEBS Lett. 2010, 584, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, R.; Xia, Y.; Wu, S.; Sun, J. Eukaryotic signaling pathways targeted by Salmonella effector protein AvrA in intestinal infection in vivo. BMC Microbiol. 2010, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Jones, R.M.; Neish, A.S. The Salmonella effector AvrA mediates bacterial intracellular survival during infection in vivo. Cell. Microbiol. 2012, 14, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M.; Black, R.E.; Lanata, C. Precise estimation of the numbers of chronic carriers of Salmonella typhi in Santiago, Chile, an endemic area. J. Infect. Dis. 1982, 146, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Parry, C.M.; Hien, T.T.; Dougan, G.; White, N.J.; Farrar, J.J. Typhoid fever. N. Engl. J. Med. 2002, 347, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.W.; Rosales-Reyes, R.; Ramirez-Aguilar Mde, L.; Chapa-Azuela, O.; Alpuche-Aranda, C.; Gunn, J.S. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. USA 2010, 107, 4353–4358. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M. Helicobacter and salmonella persistent infection strategies. Cold Spring Harb. Perspect. Med. 2013, 3, a010348. [Google Scholar] [CrossRef] [PubMed]
- Mathur, R.; Oh, H.; Zhang, D.; Park, S.G.; Seo, J.; Koblansky, A.; Hayden, M.S.; Ghosh, S. A mouse model of Salmonella typhi infection. Cell 2012, 151, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Wu, S.; Liu, X.; Xia, Y.; Zhang, Y.G.; Sun, J. Chronic effects of a Salmonella type III secretion effector protein AvrA in vivo. PLoS ONE 2010, 5, e10505. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Bosland, M.; Xia, Y.; Zhang, Y.G.; Kato, I.; Sun, J. Presence of Salmonella AvrA in colorectal tumor and its precursor lesions in mouse intestine and human specimens. Oncotarget 2017, 8, 55104–55115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Wu, S.; Zhang, Y.G.; Xia, Y.; Liu, X.; Zheng, Y.; Chen, H.; Schaefer, K.L.; Zhou, Z.; Bissonnette, M.; et al. Enteric bacterial protein AvrA promotes colonic tumorigenesis and activates colonic beta-catenin signaling pathway. Oncogenesis 2014, 3, e105. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Liu, X.; Wu, S.; Xia, Y.; Zhang, Y.G.; Petrof, E.O.; Claud, E.C.; Sun, J. Consistent activation of the beta-catenin pathway by Salmonella type-three secretion effector protein AvrA in chronically infected intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1113–G1125. [Google Scholar] [CrossRef] [PubMed]
- Johanns, T.M.; Ertelt, J.M.; Rowe, J.H.; Way, S.S. Regulatory T cell suppressive potency dictates the balance between bacterial proliferation and clearance during persistent Salmonella infection. PLoS Pathog. 2010, 6, e1001043, Published 2010 Aug 12. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.M.; Malo, D.; Vogan, K.; Skamene, E.; Gros, P. Natural resistance to infection with intracellular parasites: Isolation of a candidate for Bcg. Cell 1993, 73, 469–485. [Google Scholar] [CrossRef]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNγ neutralization. J. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Lawley, T.D.; Chan, K.; Thompson, L.J.; Kim, C.C.; Govoni, G.R.; Monack, D.M. Genome-wide screen for Salmonella genes required for long-term systemic infection of the mouse. PLoS Pathog. 2006, 2, e11. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Vallance, B.A.; Celli, J.; Winfree, S.; Hansen, B.; Montero, M.; Steele-Mortimer, O. Dissemination of invasive Salmonella via bacterial-induced extrusion of mucosal epithelia. Proc. Natl. Acad. Sci. USA 2010, 107, 17733–17738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawley, T.D.; Bouley, D.M.; Hoy, Y.E.; Gerke, C.; Relman, D.A.; Monack, D.M. Host transmission of Salmonella enterica serovar Typhimurium is controlled by virulence factors and indigenous intestinal microbiota. Infect. Immun. 2008, 76, 403–416. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, L.M.; Govoni, G.R.; Gerke, C.; Gopinath, S.; Peng, K.; Laidlaw, G.; Chien, Y.H.; Jeong, H.W.; Li, Z.; Brown, M.D.; et al. The Salmonella SPI2 effector SseI mediates long-term systemic infection by modulating host cell migration. PLoS Pathog. 2009, 5, e1000671. [Google Scholar] [CrossRef] [PubMed]
- Worley, M.J.; Nieman, G.S.; Geddes, K.; Heffron, F. Salmonella typhimurium disseminates within its host by manipulating the motility of infected cells. Proc. Natl. Acad. Sci. USA 2006, 103, 17915–17920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsley, R.A.; Van Amsterdam, K.; Kramer, N.; Bäumler, A.J. The shdA Gene Is Restricted to Serotypes ofSalmonella enterica Subspecies I and Contributes to Efficient and Prolonged Fecal Shedding. Infect. Immun. 2000, 68, 2720–2727. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Keestra, A.M.; de Zoete, M.R.; Baumler, A.J. The ShdA adhesin binds to the cationic cradle of the fibronectin 13FnIII repeat module: Evidence for molecular mimicry of heparin binding. Mol. Microbiol. 2004, 52, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Humphries, A.D.; Weening, E.H.; De Zoete, M.R.; Winter, S.; Papaconstantinopoulou, A.; Dougan, G.; Baumler, A.J. Molecular and phenotypic analysis of the CS54 island of Salmonella enterica serotype typhimurium: Identification of intestinal colonization and persistence determinants. Infect. Immun. 2003, 71, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Abi Ghanem, D.; Puebla-Osorio, N.; Keestra, A.M.; Berghman, L.; Baumler, A.J. Fibronectin binding to the Salmonella enterica serotype Typhimurium ShdA autotransporter protein is inhibited by a monoclonal antibody recognizing the A3 repeat. J. Bacteriol. 2004, 186, 4931–4939. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, C.W.; Laarakker, M.C.; Humphries, A.D.; Weening, E.H.; Baumler, A.J. Salmonella enterica serotype Typhimurium MisL is an intestinal colonization factor that binds fibronectin. Mol. Microbiol. 2005, 57, 196–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weening, E.H.; Barker, J.D.; Laarakker, M.C.; Humphries, A.D.; Tsolis, R.M.; Baumler, A.J. The Salmonella enterica serotype Typhimurium lpf, bcf, stb, stc, std, and sth fimbrial operons are required for intestinal persistence in mice. Infect. Immun. 2005, 73, 3358–3366. [Google Scholar] [CrossRef] [PubMed]
- Ruby, T.; McLaughlin, L.; Gopinath, S.; Monack, D. Salmonella’s long-term relationship with its host. FEMS Microbiol. Rev. 2012, 36, 600–615. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Fang, F.C. Oxygen-dependent anti-Salmonella activity of macrophages. Trends Microbiol. 2001, 9, 29–33. [Google Scholar] [CrossRef]
- Schürmann, N.; Forrer, P.; Casse, O.; Li, J.; Felmy, B.; Burgener, A.V.; Ehrenfeuchter, N.; Hardt, W.D.; Recher, M.; Hess, C.; et al. Myeloperoxidase targets oxidative host attacks to Salmonella and prevents collateral tissue damage. Nat. Microbiol. 2017, 2, 16268. [Google Scholar] [CrossRef] [PubMed]
- Burton, N.A.; Schürmann, N.; Casse, O.; Steeb, A.K.; Claudi, B.; Zankl, J.; Schmidt, A.; Bumann, D. Disparate Impact of Oxidative Host Defenses Determines the Fate of Salmonella during Systemic Infection in Mice. Cell Host Microbe 2014, 15, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastroeni, P.; Simmons, C.; Fowler, R.; Hormaeche, C.; Dougan, G. Igh-6−/−(B-Cell-Deficient) Mice Fail To Mount Solid Acquired Resistance to Oral Challenge with Virulent Salmonella enterica Serovar Typhimurium and Show Impaired Th1 T-Cell Responses toSalmonella Antigens. Infect. Immun. 2000, 68, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Mastroeni, P.; Vazquez-Torres, A.; Fang, F.C.; Xu, Y.; Khan, S.; Hormaeche, C.E.; Dougan, G. Antimicrobial actions of the NADPH phagocyte oxidase and inducible nitric oxide synthase in experimental salmonellosis. II. Effects on microbial proliferation and host survival in vivo. J. Exp. Med. 2000, 192, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C.; DeGroote, M.A.; Foster, J.W.; Bäumler, A.J.; Ochsner, U.; Testerman, T.; Bearson, S.; Giárd, J.-C.; Xu, Y.; Campbell, G. Virulent Salmonella typhimurium has two periplasmic Cu, Zn-superoxide dismutases. Proc. Natl. Acad. Sci. USA 1999, 96, 7502–7507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnakumar, R.; Craig, M.; Imlay, J.A.; Slauch, J.M. Differences in enzymatic properties allow SodCI but not SodCII to contribute to virulence in Salmonella enterica serovar Typhimurium strain 14028. J. Bacteriol. 2004, 186, 5230–5238. [Google Scholar] [CrossRef] [PubMed]
- Bang, I.S.; Liu, L.; Vazquez-Torres, A.; Crouch, M.L.; Stamler, J.S.; Fang, F.C. Maintenance of nitric oxide and redox homeostasis by the salmonella flavohemoglobin hmp. J. Biol. Chem. 2006, 281, 28039–28047. [Google Scholar] [CrossRef] [PubMed]
- Crum-Cianflone, N.F. Salmonellosis and the gastrointestinal tract: More than just peanut butter. Curr. Gastroenterol. Rep. 2008, 10, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. Mucosal immunity: Induction, dissemination, and effector functions. Scand. J. Immunol. 2009, 70, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Gradel, K.O.; Nielsen, H.L.; Schonheyder, H.C.; Ejlertsen, T.; Kristensen, B.; Nielsen, H. Increased short- and long-term risk of inflammatory bowel disease after salmonella or campylobacter gastroenteritis. Gastroenterology 2009, 137, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Barman, M.; Unold, D.; Shifley, K.; Amir, E.; Hung, K.; Bos, N.; Salzman, N. Enteric salmonellosis disrupts the microbial ecology of the murine gastrointestinal tract. Infect. Immun. 2008, 76, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.B.; Gill, N.; Willing, B.P.; Antunes, L.C.; Russell, S.L.; Croxen, M.A.; Finlay, B.B. The intestinal microbiota plays a role in Salmonella-induced colitis independent of pathogen colonization. PLoS ONE 2011, 6, e20338. [Google Scholar] [CrossRef] [PubMed]
- Schultz, B.M.; Salazar, G.A.; Paduro, C.A.; Pardo-Roa, C.; Pizarro, D.P.; Salazar-Echegarai, F.J.; Torres, J.; Riedel, C.A.; Kalergis, A.M.; Alvarez-Lobos, M.M.; et al. Persistent Salmonella enterica serovar Typhimurium Infection Increases the Susceptibility of Mice to Develop Intestinal Inflammation. Front. Immunol. 2018, 9, 1166. [Google Scholar] [CrossRef] [PubMed]
- Godinez, I.; Raffatellu, M.; Chu, H.; Paixao, T.A.; Haneda, T.; Santos, R.L.; Bevins, C.L.; Tsolis, R.M.; Baumler, A.J. Interleukin-23 orchestrates mucosal responses to Salmonella enterica serotype Typhimurium in the intestine. Infect. Immun. 2009, 77, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, C.V.; Mercado-Lubo, R.; Hallstrom, K.; McCormick, B.A. Salmonella effector proteins and host-cell responses. Cell. Mol. Life Sci. 2011, 68, 3687–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.G.; Wu, S.; Xia, Y.; Sun, J. Salmonella infection upregulates the leaky protein claudin-2 in intestinal epithelial cells. PLoS ONE 2013, 8, e58606. [Google Scholar] [CrossRef] [PubMed]
- Grassl, G.A.; Valdez, Y.; Bergstrom, K.S.; Vallance, B.A.; Finlay, B.B. Chronic enteric salmonella infection in mice leads to severe and persistent intestinal fibrosis. Gastroenterology 2008, 134, 768–780. [Google Scholar] [CrossRef] [PubMed]
- McCormick, B.A. Using Salmonella enterica serotype typhimurium to model intestinal fibrosis. Gastroenterology 2008, 134, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Rodansky, E.S.; Moons, D.S.; Larsen, S.D.; Neubig, R.R.; Higgins, P.D.R. Optimisation of Intestinal Fibrosis and Survival in the Mouse S. Typhimurium Model for Anti-fibrotic Drug Discovery and Preclinical Applications. J. Crohns Colitis 2017, 11, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Mansson, L.E.; Montero, M.; Zarepour, M.; Bergstrom, K.S.; Ma, C.X.; Huang, T.; Man, C.; Grassl, G.A.; Vallance, B.A. MyD88 signaling promotes both mucosal homeostatic and fibrotic responses during Salmonella-induced colitis. Am. J. Physiol.-Gastr. L 2012, 303, G311–G323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polk, D.B.; Peek, R.M., Jr. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.T.; Israel, D.A.; Washington, M.K.; Krishna, U.; Fox, J.G.; Rogers, A.B.; Neish, A.S.; Collier-Hyams, L.; Perez-Perez, G.I.; Hatakeyama, M.; et al. Activation of beta-catenin by carcinogenic Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2005, 102, 10646–10651. [Google Scholar] [CrossRef] [PubMed]
- Caygill, C.P.; Hill, M.J.; Braddick, M.; Sharp, J.C. Cancer mortality in chronic typhoid and paratyphoid carriers. Lancet 1994, 343, 83–84. [Google Scholar] [CrossRef]
- Hundal, R.; Shaffer, E.A. Gallbladder cancer: Epidemiology and outcome. Clin. Epidemiol. 2014, 6, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, K.L.; Gupta, A.; Yadav, A.; Kumar, A. Gallbladder cancer epidemiology, pathogenesis and molecular genetics: Recent update. World J. Gastroenterol. 2017, 23, 3978–3998. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Hoshi, T.; Ogawa, K. K-ras codon 12 mutations in biliary tract tumors detected by polymerase chain reaction denaturing gradient gel electrophoresis. Cancer 1994, 73, 2727–2733. [Google Scholar] [CrossRef]
- Billo, P.; Marchegiani, C.; Capella, C.; Sessa, F. Expression of p53 in gallbladder carcinoma and in dysplastic and metaplastic lesions of the surrounding mucosa. Pathologica 2000, 92, 249–256. [Google Scholar] [PubMed]
- Kiguchi, K.; Carbajal, S.; Chan, K.; Beltran, L.; Ruffino, L.; Shen, J.; Matsumoto, T.; Yoshimi, N.; DiGiovanni, J. Constitutive expression of ErbB-2 in gallbladder epithelium results in development of adenocarcinoma. Cancer Res. 2001, 61, 6971–6976. [Google Scholar] [PubMed]
- Song, J.; Gao, X.; Galan, J.E. Structure and function of the Salmonella Typhi chimaeric A(2)B(5) typhoid toxin. Nature 2013, 499, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Scuron, M.D.; Boesze-Battaglia, K.; Dlakic, M.; Shenker, B.J. The Cytolethal Distending Toxin Contributes to Microbial Virulence and Disease Pathogenesis by Acting As a Tri-Perditious Toxin. Front. Cell. Infect. Microbiol. 2016, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, E.G.; Cavallo, I.; Pontone, M.; Toma, L.; Ensoli, F. Biofilm Producing Salmonella Typhi: Chronic Colonization and Development of Gallbladder Cancer. Int. J. Mol. Sci. 2017, 18, 1887. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.; Barreto, S.G.; Sahoo, B.; Chandrani, P.; Ramadwar, M.R.; Shrikhande, S.V.; Dutt, A. Non-typhoidal Salmonella DNA traces in gallbladder cancer. Infect. Agent Cancer 2016, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Butin-Israeli, V.; Bui, T.M.; Wiesolek, H.L.; Mascarenhas, L.; Lee, J.J.; Mehl, L.C.; Knutson, K.R.; Adam, S.A.; Goldman, R.D.; Beyder, A.; et al. Neutrophil-induced genomic instability impedes resolution of inflammation and wound healing. J. Clin. Investig. 2019, 129, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Buchhop, S.; Gibson, M.K.; Wang, X.W.; Wagner, P.; Sturzbecher, H.W.; Harris, C.C. Interaction of p53 with the human Rad51 protein. Nucleic Acids Res. 1997, 25, 3868–3874. [Google Scholar] [CrossRef] [PubMed]
- Arias-Lopez, C.; Lazaro-Trueba, I.; Kerr, P.; Lord, C.J.; Dexter, T.; Iravani, M.; Ashworth, A.; Silva, A. p53 modulates homologous recombination by transcriptional regulation of the RAD51 gene. EMBO Rep. 2006, 7, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Ye, Z.; Liu, X.; Zhao, Y.; Xia, Y.; Steiner, A.; Petrof, E.O.; Claud, E.C.; Sun, J. Salmonella typhimurium infection increases p53 acetylation in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G784–G794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feagins, L.A.; Souza, R.F.; Spechler, S.J. Carcinogenesis in IBD: Potential targets for the prevention of colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-κB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.; Karin, E.; Terzic, J.; Mucida, D.; Yu, G.Y.; Vallabhapurapu, S.; Scheller, J.; Rose-John, S.; Cheroutre, H.; Eckmann, L.; et al. IL-6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell 2009, 15, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Wu, S.; Zhang, Y.G.; Xia, Y.; Zhou, Z.; Kato, I.; Dong, H.; Bissonnette, M.; Sun, J. Salmonella Protein AvrA Activates the STAT3 Signaling Pathway in Colon Cancer. Neoplasia 2016, 18, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Finlay, B.B.; Steele-Mortimer, O. The Salmonella effector protein SopB protects epithelial cells from apoptosis by sustained activation of Akt. J. Biol. Chem. 2005, 280, 9058–9064. [Google Scholar] [CrossRef] [PubMed]
- Tahoun, A.; Mahajan, S.; Paxton, E.; Malterer, G.; Donaldson, D.S.; Wang, D.; Tan, A.; Gillespie, T.L.; O’Shea, M.; Roe, A.J.; et al. Salmonella transforms follicle-associated epithelial cells into M cells to promote intestinal invasion. Cell Host Microbe 2012, 12, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Zavadil, J.; Bottinger, E.P. TGF-beta and epithelial-to-mesenchymal transitions. Oncogene 2005, 24, 5764–5774. [Google Scholar] [CrossRef] [PubMed]
- DSe Wind, N.; Dekker, M.; van Rossum, A.; van der Valk, M.; te Riele, H. Mouse models for hereditary nonpolyposis colorectal cancer. Cancer Res. 1998, 58, 248–255. [Google Scholar] [PubMed]
- Scanu, T.; Spaapen, R.M.; Bakker, J.M.; Pratap, C.B.; Wu, L.E.; Hofland, I.; Broeks, A.; Shukla, V.K.; Kumar, M.; Janssen, H.; et al. Salmonella Manipulation of Host Signaling Pathways Provokes Cellular Transformation Associated with Gallbladder Carcinoma. Cell Host Microbe 2015, 17, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Chen, L.M.; Hernandez, L.; Shears, S.B.; Galán, J.E. A Salmonella inositol polyphosphatase acts in conjunction with other bacterial effectors to promote host cell actin cytoskeleton rearrangements and bacterial internalization. Mol. Microbiol. 2001, 39, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.G.; Wu, S.; Xia, Y.; Sun, J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiol. Rep. 2014, 2, e12147. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessein, R.; Gironella, M.; Vignal, C.; Peyrin-Biroulet, L.; Sokol, H.; Secher, T.; Gratadoux, J.-J.; Lafont, F.; Dagorn, J.-C.; Ryffel, B. TLR2 is critical for induction of REG3β expression and intestinal clearance of Yersinia pseudotuberculosis. Gut 2009, 58, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Stelter, C.; Käppeli, R.; König, C.; Krah, A.; Hardt, W.-D.; Stecher, B.; Bumann, D. Salmonella-induced mucosal lectin RegIIIβ kills competing gut microbiota. PLoS ONE 2011, 6, e20749. [Google Scholar] [CrossRef] [PubMed]
- Thiennimitr, P.; Winter, S.E.; Winter, M.G.; Xavier, M.N.; Tolstikov, V.; Huseby, D.L.; Sterzenbach, T.; Tsolis, R.M.; Roth, J.R.; Baumler, A.J. Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 17480–17485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.T.; Shabahang, S.; Timiryasova, T.; Zhang, Q.; Beltz, R.; Gentschev, I.; Goebel, W.; Szalay, A. Visualization of Tumors and Metastases in Live Animals with Injected Bacteria and Vaccinia Virus Encoding Light-Emitting Proteins; AACR: Philadelphia, PA, USA, 2005. [Google Scholar]
- Adkins, I.; Holubova, J.; Kosova, M.; Sadilkova, L. Bacteria and their toxins tamed for immunotherapy. Curr. Pharm. Biotechnol. 2012, 13, 1446–1473. [Google Scholar] [CrossRef] [PubMed]
- Hand, N. Development of a Recombinant Attenuated Salmonella Cancer Vaccine. Ph.D. Thesis, The George Washington University, Washington, DC, USA, 2018. [Google Scholar]
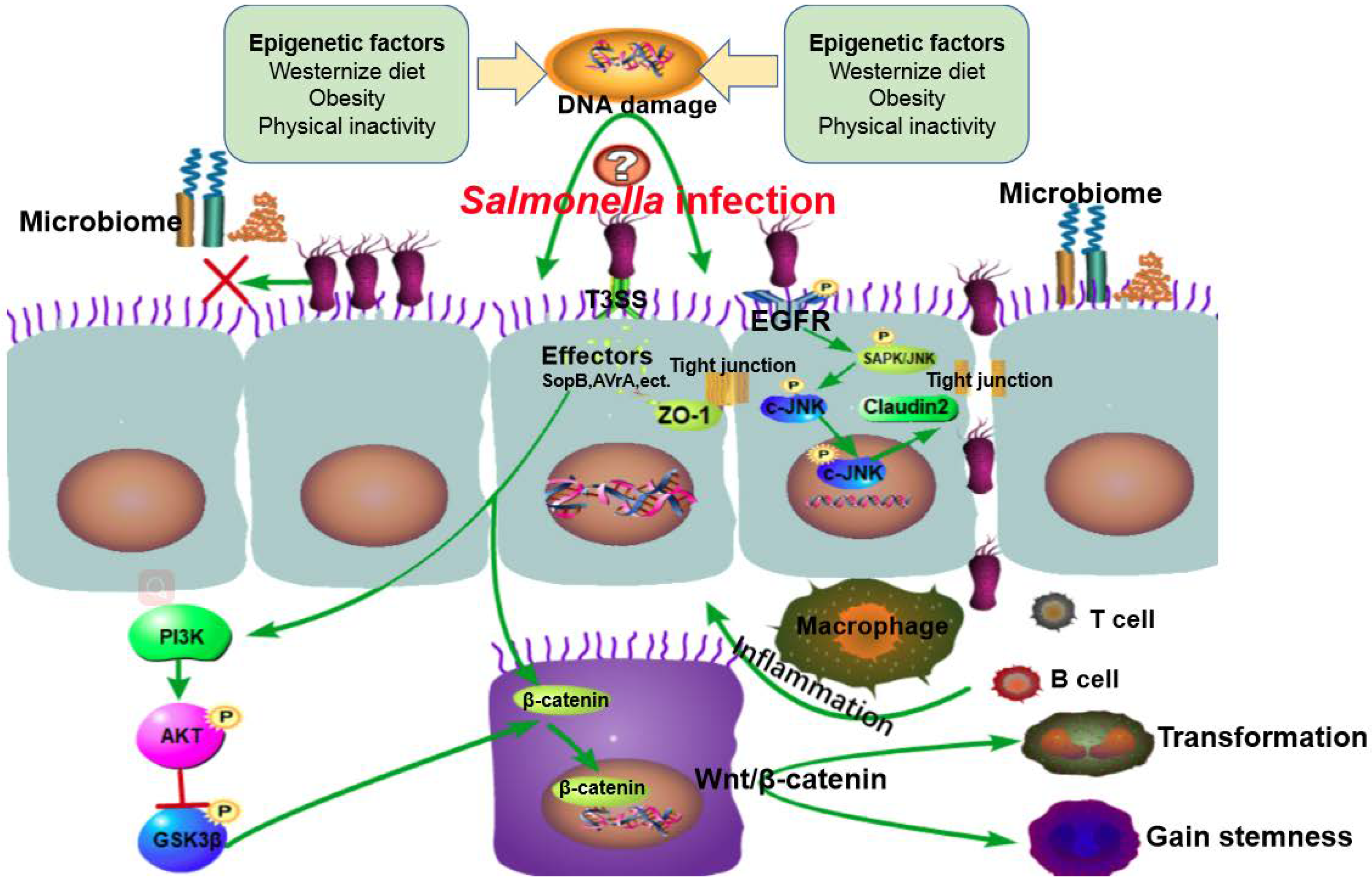
Figure 1.
The working model summarizing the roles of Salmonella infection in cancer. Salmonella infection induces carcinogenesis potentially through four paths: (1) the host is preconditioned by DNA damage caused by epigenetic factors (Western diet, obesity and physical inactivity) and genetic factors (tumor susceptibility); (2) Salmonella effector protein AvrA stabilizes the expression of tight junction proteins (e.g., ZO-1) and decreases inflammation. Meanwhile, Salmonella targets the “leaky protein” claudin-2 to facilitate bacterial invasion. The balance between protection and injury contributes to chronic infection and leads to sustained epithelial cell injury and DNA mutation; (3) Salmonella competes with indigenous microbiota for intestinal epithelial attachment sites and nutrients, thereby disrupting the gut microbiome and overcoming colonization resistance; and (4) T3SS effectors enter epithelial cells and activates signaling pathways leading to chronic inflammation, host cell transformation, and carcinogenesis.
Figure 1.
The working model summarizing the roles of Salmonella infection in cancer. Salmonella infection induces carcinogenesis potentially through four paths: (1) the host is preconditioned by DNA damage caused by epigenetic factors (Western diet, obesity and physical inactivity) and genetic factors (tumor susceptibility); (2) Salmonella effector protein AvrA stabilizes the expression of tight junction proteins (e.g., ZO-1) and decreases inflammation. Meanwhile, Salmonella targets the “leaky protein” claudin-2 to facilitate bacterial invasion. The balance between protection and injury contributes to chronic infection and leads to sustained epithelial cell injury and DNA mutation; (3) Salmonella competes with indigenous microbiota for intestinal epithelial attachment sites and nutrients, thereby disrupting the gut microbiome and overcoming colonization resistance; and (4) T3SS effectors enter epithelial cells and activates signaling pathways leading to chronic inflammation, host cell transformation, and carcinogenesis.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zha, L.; Garrett, S.; Sun, J. Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer. Diseases 2019, 7, 28. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases7010028
AMA Style
Zha L, Garrett S, Sun J. Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer. Diseases. 2019; 7(1):28. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases7010028
Chicago/Turabian StyleZha, Lang, Shari Garrett, and Jun Sun. 2019. "Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer" Diseases 7, no. 1: 28. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases7010028
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.