Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions and Their Implications in Malaria Control

 and
and
Abstract
:1. Introduction
2. Methods
2.1. Systematic Review Protocols
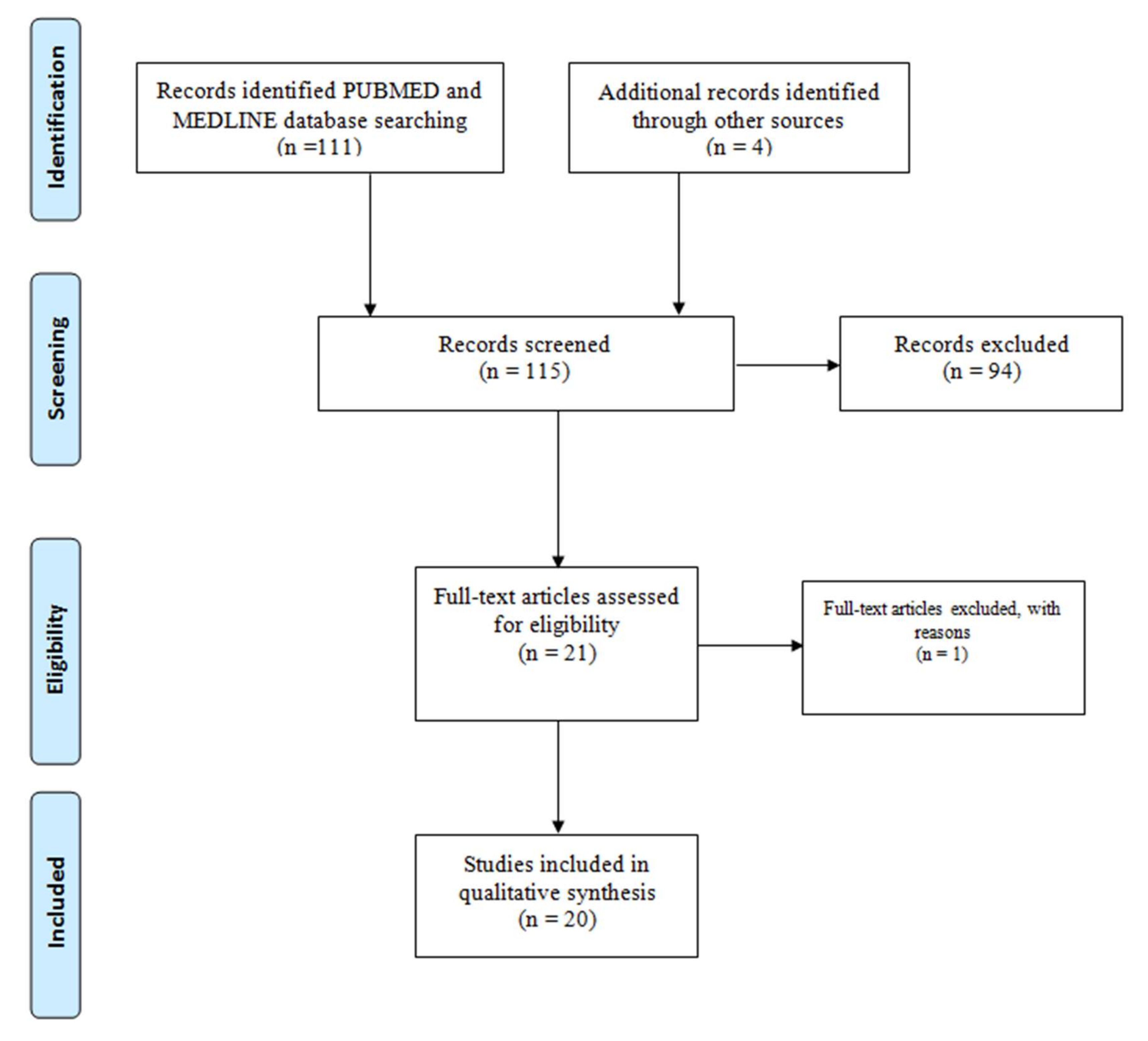
2.2. Search Strategy and Inclusion/Exclusion Criteria
2.3. Statistical Analysis
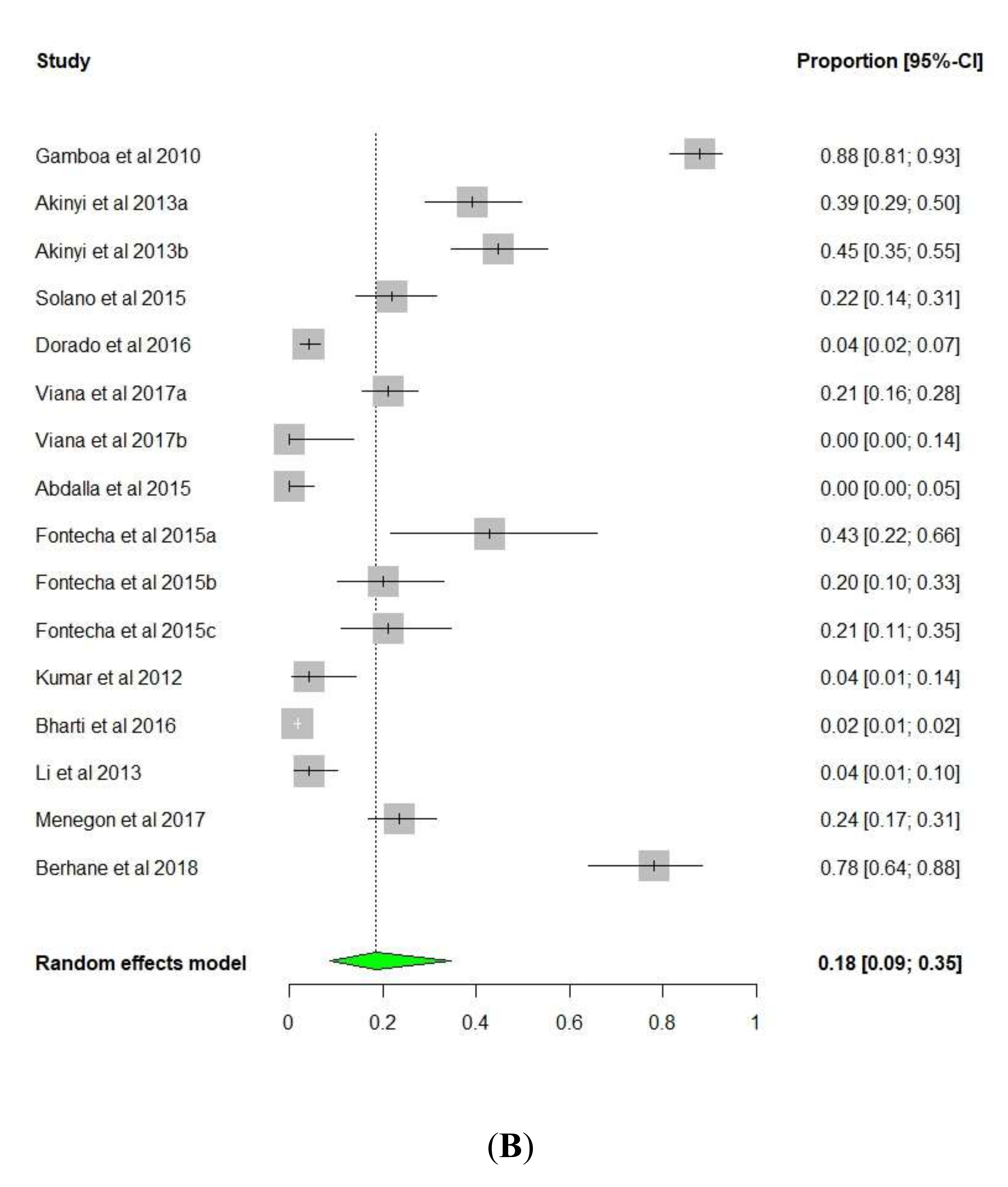
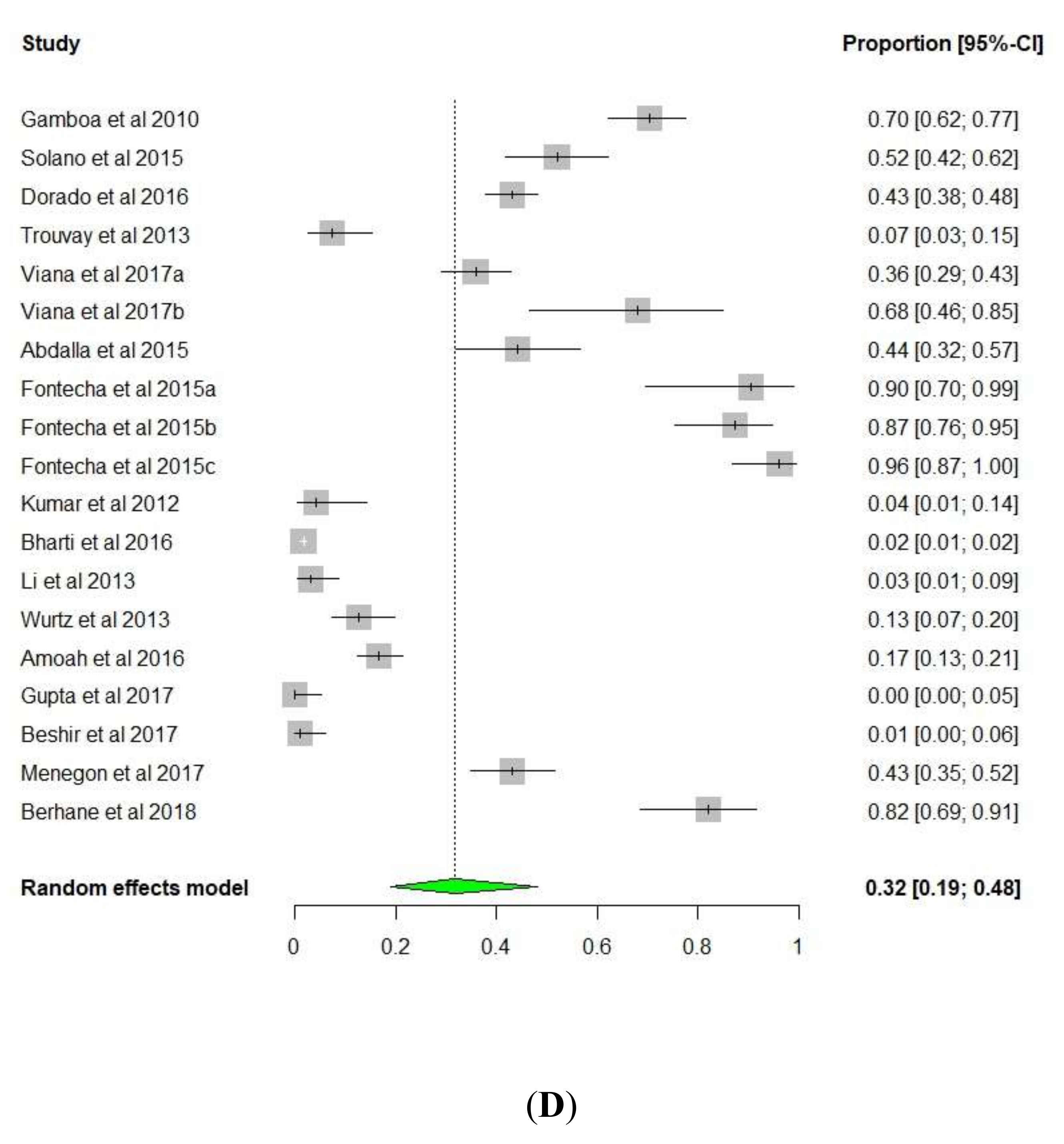
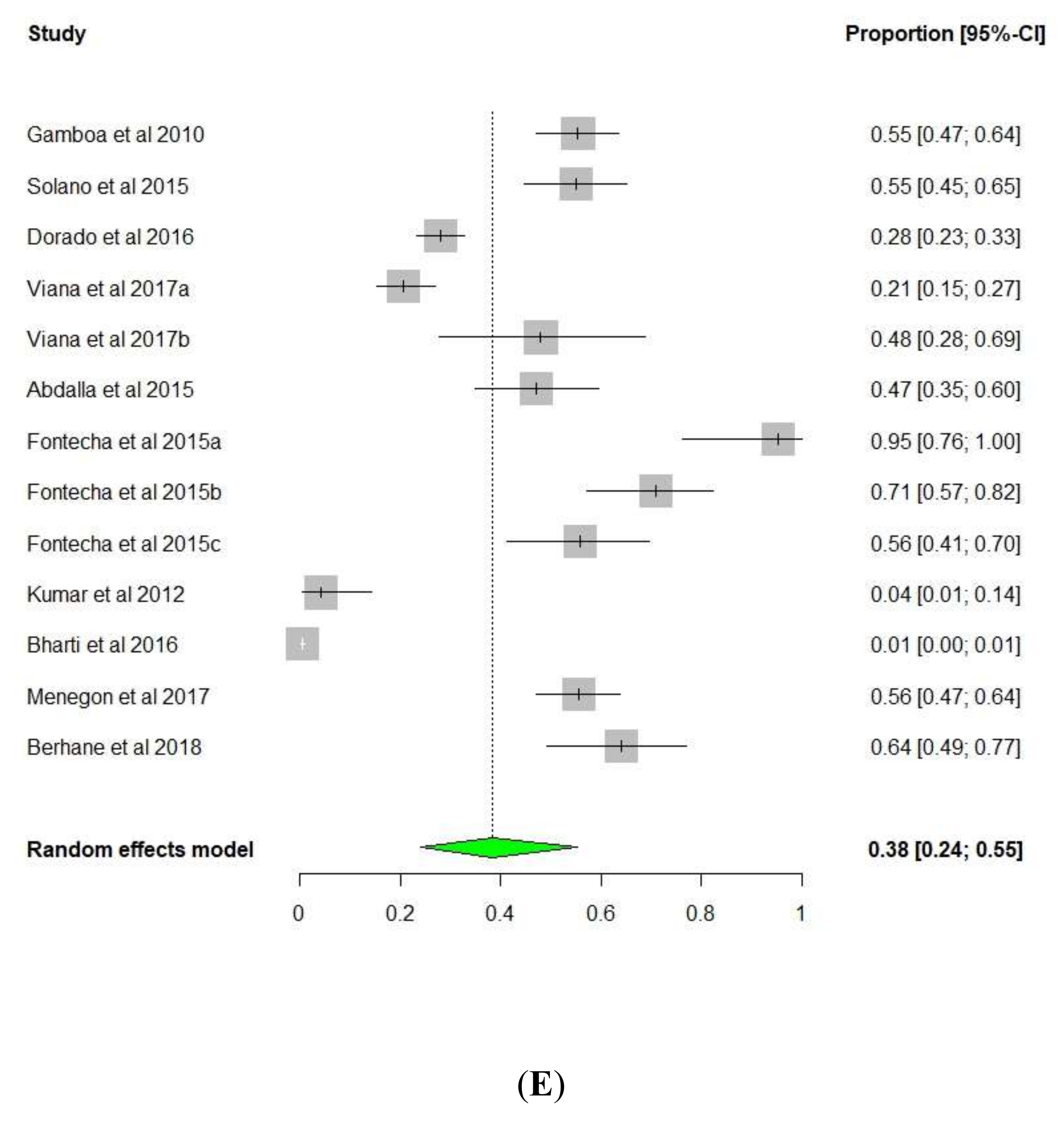
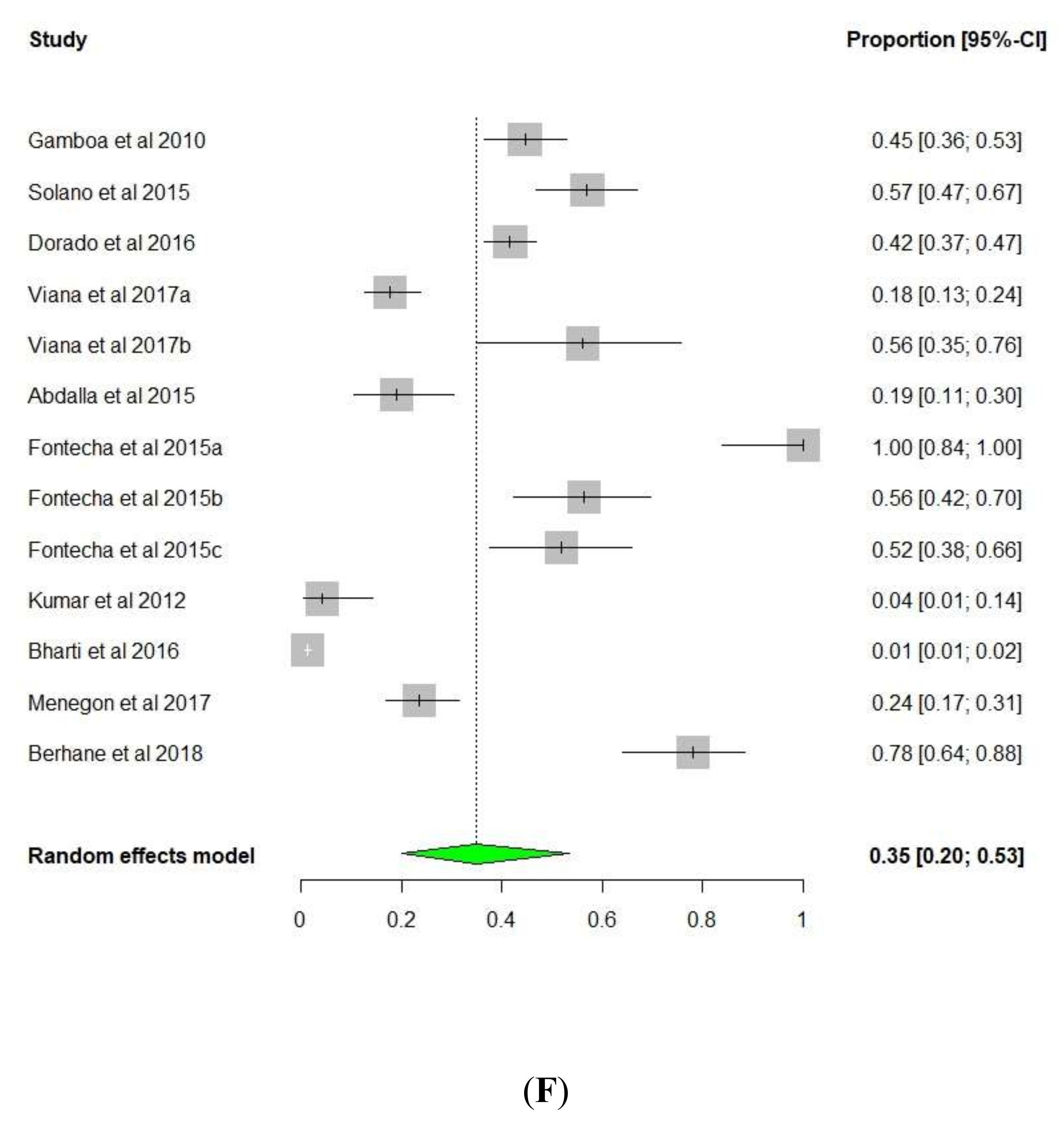
3. Results and Discussion
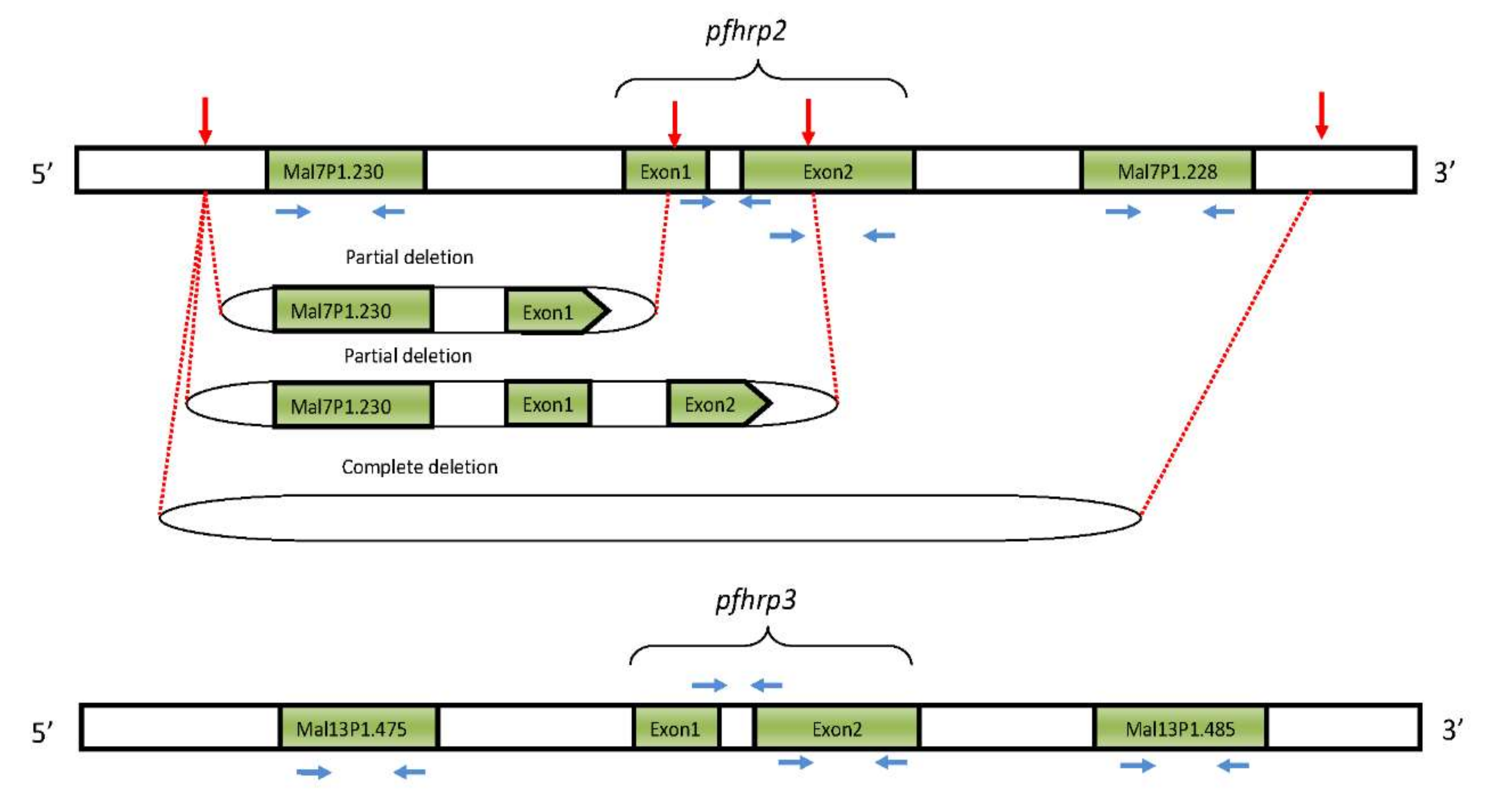
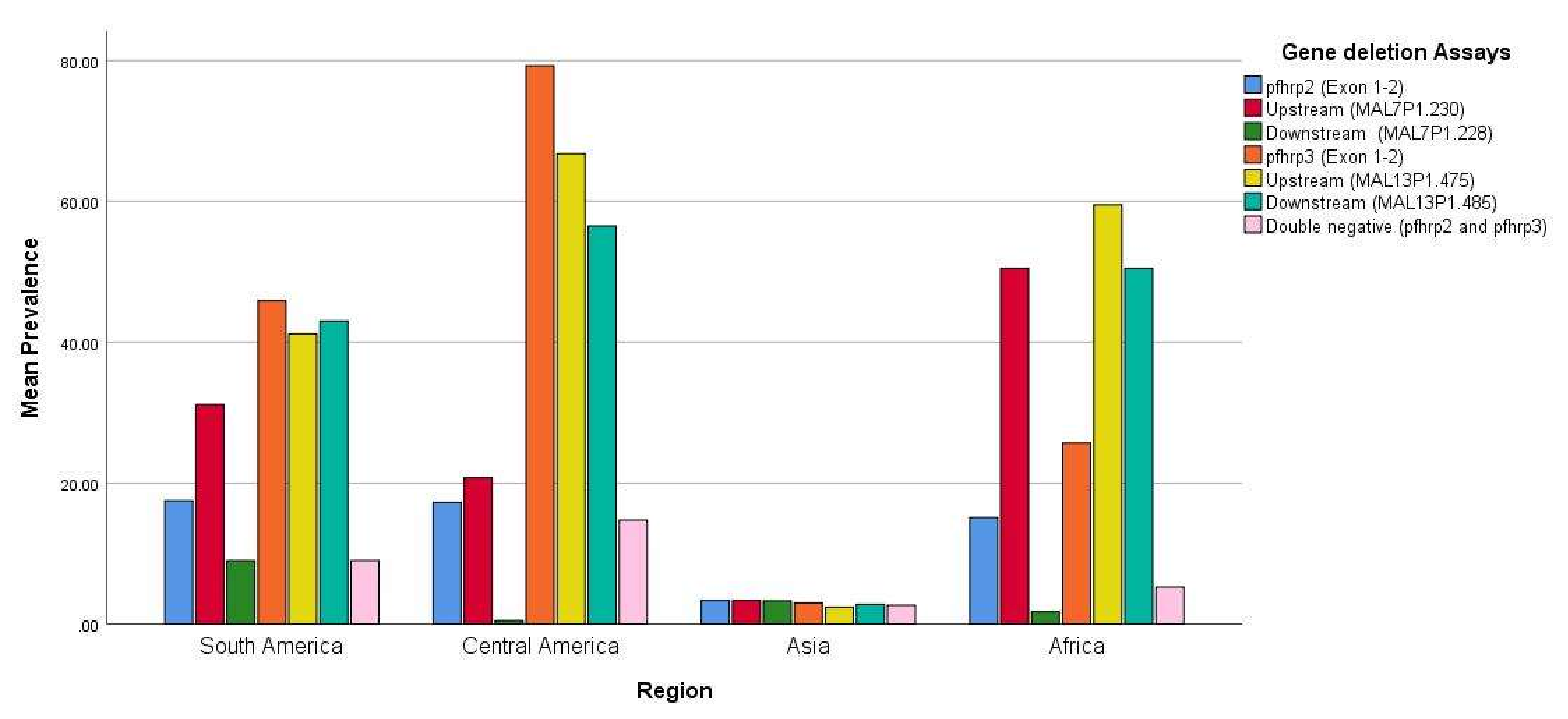
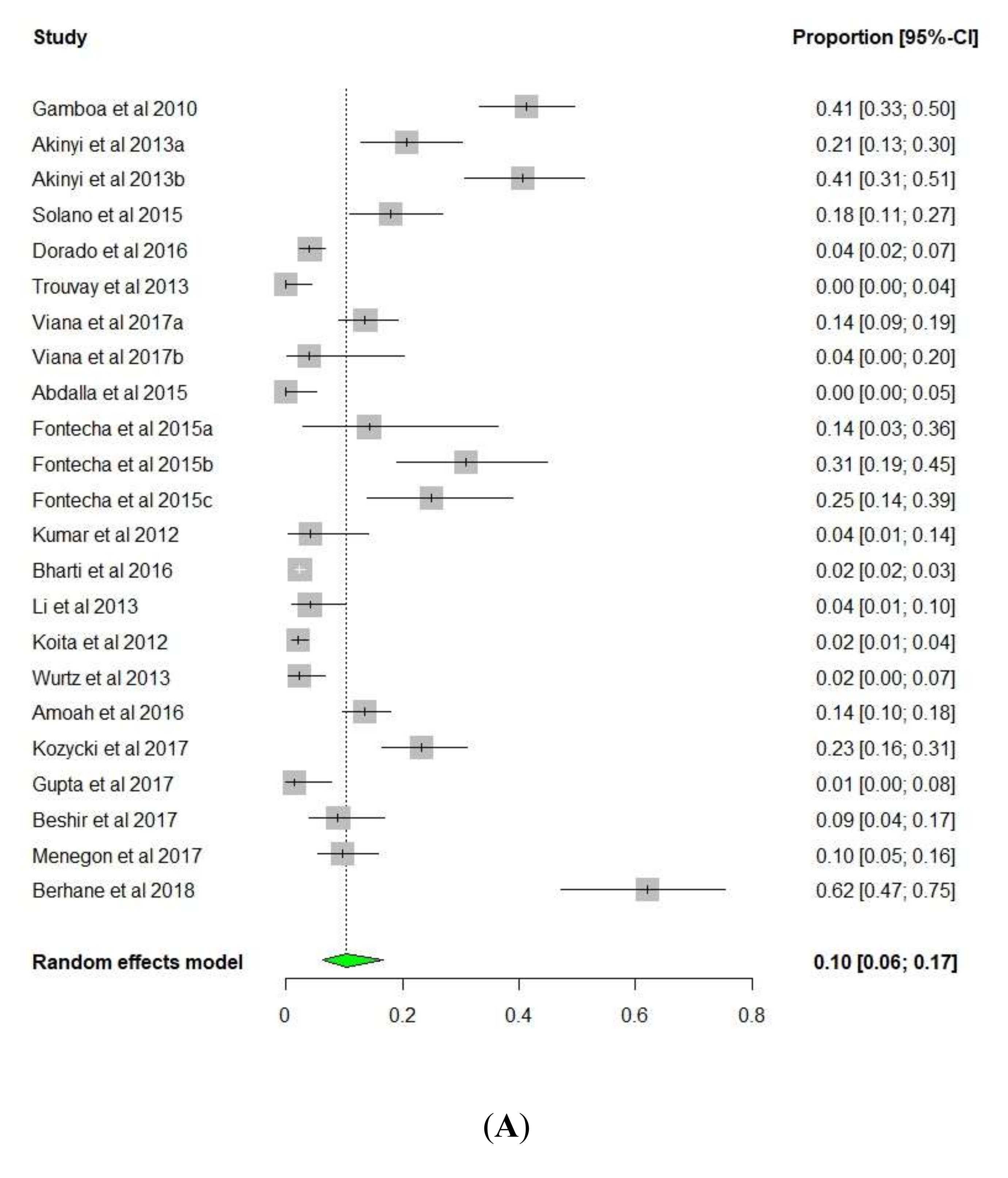
3.1. pfhrp2/pfhrp3 Deletions
3.2. Implication of pfhrp2/pfhrp3 Deletions on RDT Use
Funding
Acknowledgments
Conflicts of Interest
References
- Centers for Dises Control and Prevention. World Malaria Day 2018. Features; Centers for Dises Control and Prevention: Atlanta, GA, USA, 2018; Available online: https://www.cdc.gov/features/worldmalariaday/index.html (accessed on 12 October 2018).
- WHO. World Malaria Report 2016; World Health Organization: Geneva, Switzerland, 2016; Available online: https://www.who.int/malaria/publications/world-malaria-report-2016/report/en/ (accessed on 12 October 2018).
- WHO. WHO News Release. Available online: https://www.who.int/mediacentre/news/releases/2011/malaria_report_20111213/en/ (accessed on 22 October 2018).
- WHO. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019; Available online: https://www.who.int/publications-detail/world-malaria-report-2019 (accessed on 12 October 2018).
- WHO. Guidlines for the Treatment of Malaria; World Health Organization: Geneva, Switzerland, 2015; Available online: https://www.who.int/malaria/publications/atoz/9789241549127/en/ (accessed on 6 July 2019).
- Tangpukdee, N.; Duangdee, C.; Wilairatana, P.; Krudsood, S. Malaria diagnosis: A brief review. Korean J. Parasitol. 2009, 47, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Jirk, M.; Petrželková, K.J.; Zuzana, H.; Modrý, D.; Lukeš, J. Detection of Plasmodium spp. in Human Feces. Emerg. Infect. Dis. 2012, 18, 634–636. [Google Scholar]
- Putaporntip, C.; Jongwutiwes, S.; Sakihama, N.; Ferreira, M.U.; Kho, W.G.; Kaneko, A.; Kanbara, H.; Hattori, T.; Tanabe, K. Mosaic organization and heterogeneity in frequency of allelic recombination of the Plasmodium vivax merozoite surface protein-1 locus. Proc. Natl. Acad. Sci. USA 2002, 99, 16348–16353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongsrichanalai, C.; Barcus, M.J.; Muth, S.; Sutamihardja, A.; Wernsdorfer, W.H. A Review of Malaria Diagnostic Tools: Microscopy and Rapid Diagnostic Test (RDT). Am. J. Trop. Med. Hyg. 2007, 77, 119–127. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK1695/ (accessed on 22 October 2018). [CrossRef] [PubMed] [Green Version]
- Murphy, S.C.; Shott, J.P.; Parikh, S.; Etter, P.; Prescott, W.R.; Stewart, V.A. Review article: Malaria diagnostics in clinical trials. Am. J. Trop. Med. Hyg. 2013, 89, 824–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirdha, B.R.; Samantaray, J.C.; Mishra, B. Laboratory diagnosis of malaria. J. Clin. Pathol. 1997, 50, 356. [Google Scholar] [CrossRef] [Green Version]
- Beshir, K.B.; Sepúlveda, N.; Bharmal, J.; Robinson, A.; Mwanguzi, J.; Busula, A.O.; De Boer, J.G.; Sutherland, C.; Cunningham, J.; Hopkins, H. Plasmodium falciparum parasites with histidine-rich protein 2 (pfhrp2) and pfhrp3 gene deletions in two endemic regions of Kenya. Sci. Rep. 2017, 7, 14718. [Google Scholar] [CrossRef]
- Khairnar, K.; Martin, D.; Lau, R.; Ralevski, F.; Pillai, D.R. Multiplex real-time quantitative PCR, microscopy and rapid diagnostic immuno-chromatographic tests for the detection of Plasmodium spp: Performance, limit of detection analysis and quality assurance. Malar. J. 2009, 8, 284. [Google Scholar] [CrossRef] [Green Version]
- Ohrt, C.; Sutamihardja, M.A.; Tang, D.; Kain, K.C. Impact of Microscopy Error on Estimates of Protective Efficacy in Malaria-Prevention Trials. J. Infect. Dis. 2002, 186, 540–546. [Google Scholar] [CrossRef]
- Payne, D. Use and limitations of light microscopy for diagnosing malaria at the primary health care level. Bull. World Health Organ. 1988, 66, 621–626. [Google Scholar]
- Snounou, G.; Viriyakosol, S.; Jarra, W.; Thaithong, S.; Brown, K.N. Identification of the four human malaria parasite species in field samples by the polymerase chain reaction and detection of a high prevalence of mixed infections. Mol. Biochem. Parasitol. 1993, 58, 283–292. [Google Scholar] [CrossRef]
- Erdman, L.K.; Kain, K.C. Molecular diagnostic and surveillance tools for global malaria control. Travel Med. Infect. Dis. 2008, 6, 82–99. [Google Scholar] [CrossRef]
- Zhao, J.; Lama, M.; Korenromp, E.; Aylward, P.; Shargie, E.; Filler, S.; Komatsu, R.; Atun, R. Adoption of rapid diagnostic tests for the diagnosis of malaria, a preliminary analysis of the global fund program data, 2005 to 2010. PLoS ONE 2012, 7, e43549. [Google Scholar] [CrossRef] [PubMed]
- WHO. Malaria Rapid Diagnostic Test Performance: Summary Results of WHO Product Testing of Malaria RDTs: Round 1-5 (2008–2013); WHO: Geneva, Switzerland, 2014; Available online: https://www.who.int/malaria/publications/atoz/9789241507554/en/ (accessed on 6 July 2019).
- Meier, B.; Döbeli, H.; Certa, U. Stage-specific expression of aldolase isoenzymes in the rodent malaria parasite Plasmodium berghei. Mol. Biochem. Parasitol. 1992, 52, 15–27. [Google Scholar] [CrossRef]
- Makler, M.T.; Hinrichs, D.J. Measurement of the lactate dehydrogenase activity of Plasmodium falciparum as an assessment of parasitemia. Am. J. Trop. Med. Hyg. 1993, 48, 205–210. [Google Scholar] [CrossRef]
- Wiwanitkit, V. Plasmodium and Host Lactate Dehydrogenase Molecular Function and Biological Pathways: Implication for Antimalarial Drug Discovery. Chem. Biol. Drug Des. 2007, 69, 280–283. [Google Scholar] [CrossRef]
- Ouattara, A.; Doumbo, S.; Saye, R.; Beavogui, A.H.; Traoré, B.; Djimdé, A.; Niangaly, A.; Kayentao, K.; Diallo, M.; Doumbo, O.K.; et al. Use of a pLDH-based dipstick in the diagnostic and therapeutic follow-up of malaria patients in Mali. Malar. J. 2011, 10, 345. [Google Scholar] [CrossRef] [Green Version]
- Dalrymple, U.; Arambepola, R.; Gething, P.W.; Cameron, E. How long do rapid diagnostic tests remain positive after anti-malarial treatment? Malar. J. 2018, 17, 228. [Google Scholar] [CrossRef] [Green Version]
- Chiodini, P.L.; Bowers, K.; Jorgensen, P.; Barnwell, J.W.; Grady, K.K.; Luchavez, J.; Moody, A.H.; Cenizal, A.; Bell, D. The heat stability of Plasmodium lactate dehydrogenase-based and histidine-rich protein 2-based malaria rapid diagnostic tests. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 331–337. [Google Scholar] [CrossRef]
- Singh, N.; Bharti, P.K.; Singh, M.P.; Mishra, S.; Shukla, M.M.; Sharma, R.K.; Singh, R.K. Comparative Evaluation of Bivalent Malaria Rapid Diagnostic Tests versus Traditional Methods in Field with Special Reference to Heat Stability Testing in Central India. PLoS ONE 2013, 8, e58080. [Google Scholar] [CrossRef] [Green Version]
- Rock, E.P.; Marsh, K.; Saul, A.J.; Wellems, T.E.; Taylor, D.W.; Maloy, W.L.; Howard, R.J. Comparative analysis of the Plasmodium falciparum histidine-rich proteins HRP-I, HRP-II and HRP-III in malaria parasites of diverse origin. Parasitology 1987, 95, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.D. Genomic organization, structure and possible function of histidine-rich proteins of malaria parasites. Int. J. Biochem. 1988, 20, 471–477. [Google Scholar] [CrossRef]
- Gruenberg, J.; Allred, D.R.; Sherman, I.W. Scanning electron microscope-analysis of the protrusions (knobs) present on the surface of Plasmodium falciparum-infected erythrocytes. J. Cell Biol. 1983, 97, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Kilejian, A. Characterization of a protein correlated with the production of knob-like protrusions on membranes of erythrocytes infected with Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1979, 76, 4650–4653. [Google Scholar] [CrossRef] [Green Version]
- Luse, S.A.; Miller, L.H. Plasmodium falciparum malaria. Ultrastructure of parasitized erythrocytes in cardiac vessels. Am. J. Trop. Med. Hyg. 1971, 20, 655–660. [Google Scholar] [CrossRef]
- Manning, L.; Laman, M.; Stanisic, D.; Rosanas-Urgell, A.; Bona, C.; Teine, D.; Siba, P.; Mueller, I.; Davis, T.M.E. Plasma Plasmodium falciparum histidine-rich protein-2 concentrations do not reflect severity of malaria in Papua New Guinean children. Clin. Infect. Dis. 2011, 52, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Parra, M.E.; Evans, C.B.; Taylor, D.W. Identification of Plasmodium falciparum Histidine-Rich Protein 2 in the Plasma of Humans with Malaria. J. Clin. Microbiol. 1991, 29, 1629–1634. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-del Valle, M.; Quakyi, I.A.; Amuesi, J.; Quaye, J.T.; Nkrumah, F.; Taylor, D.W. Detection of antigen and antibodies and in urine of humans with Plasmodium falciparum malaria. J. Clin. Microbiol. 1991, 29, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Howard, R.; Uni, S.; Aley, S.B.; Taylorll, W.; Diseases, I.; Francisco, S.; Hospital, G.; Francisco, S. Plasmodium falciparum-infected. J. Cell Biol. 1986, 103, 1269–1277. [Google Scholar] [CrossRef] [Green Version]
- Desakorn, V.; Dondorp, A.M.; Silamut, K.; Pongtavornpinyo, W.; Sahassananda, D.; Chotivanich, K.; Pitisuttithum, P.; Smithyman, A.M.; Day, N.P.J.; White, N.J. Stage-dependent production and release of histidine-rich protein 2 by Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 517–524. [Google Scholar] [CrossRef]
- Moody, A. Rapid diagnostic tests for malaria parasites. Clin. Microbiol. Rev. 2002, 15, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leech, J.H.; Aley, S.B.; Miller, L.H.; Howard, R.J. Plasmodium falciparum malaria: Cytoadherence of infected erythrocytes to endothelial cells and associated changes in the erythrocyte membrane. Prog. Clin. Biol. Res. 1984, 155, 63–77. [Google Scholar] [PubMed]
- Abdallah, J.F.; Okoth, S.A.; Fontecha, G.A.; Mejia Torres, R.E.; Banegas, E.I.; Matute, M.L.; Bucheli, S.T.M.; Goldman, I.F.; De Oliveira, A.M.; Barnwell, J.W.; et al. Prevalence of pfhrp2 and pfhrp3 gene deletions in Puerto Lempira, Honduras. Malar. J. 2015, 14, 19. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, D.; Ho, M.F.; Bendezu, J.; Torres, K.; Chiodini, P.L.; Barnwell, J.W.; Incardona, S.; Perkins, M.; Bell, D.; McCarthy, J.; et al. A large proportion of P. falciparum isolates in the Amazon region of Peru lack pfhrp2 and pfhrp3: Implications for malaria rapid diagnostic tests. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoth, S.A.; Abdallah, J.F.; Ceron, N.; Adhin, M.R.; Chandrabose, J.; Krishnalall, K.; Huber, C.S.; Goldman, I.F.; De Oliveira, A.M.; Barnwell, J.W.; et al. Variation in Plasmodium falciparum histidine-rich protein 2 (Pfhrp2) and Plasmodium falciparum histidine-rich protein 3 (Pfhrp3) gene deletions in Guyana and Suriname. PLoS ONE 2015, 10, e0126805. [Google Scholar]
- Viana, G.M.R.; Okoth, S.A.; Silva-Flannery, L.; Barbosa, D.R.L.; De Oliveira, A.M.; Goldman, I.F.; Morton, L.C.; Huber, C.; Anez, A.; Machado, R.L.D.; et al. Histidine-rich protein 2 (pfhrp2) and pfhrp3 gene deletions in Plasmodium falciparum isolates from select sites in Brazil and Bolivia. PLoS ONE 2017, 12, e0171150. [Google Scholar]
- Walker-Jonah, A.; Dolan, S.A.; Gwadz, R.W.; Panton, L.J.; Wellems, T.E. An RFLP map of the Plasmodium falciparum genome, recombination rates and favored linkage groups in a genetic cross. Mol. Biochem. Parasitol. 1992, 51, 313–320. [Google Scholar] [CrossRef]
- Pologe, L.G.; Ravetch, J.V. Large deletions result from breakage and healing of P. falciparum chromosomes. Cell 1988, 55, 869–874. [Google Scholar] [CrossRef]
- Dharia, N.V.; Plouffe, D.; Bopp, S.E.R.; González-Páez, G.E.; Lucas, C.; Salas, C.; Soberon, V.; Bursulaya, B.; Kochel, T.J.; Bacon, D.J.; et al. Genome scanning of Amazonian Plasmodium falciparum shows subtelomeric instability and clindamycin-resistant parasites. Genome Res. 2010, 20, 1534–1544. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; McCarthy, J.; Gatton, M.; Kyle, D.E.; Belizario, V.; Luchavez, J.; Bell, D.; Cheng, Q. Genetic Diversity of Plasmodium falciparum Histidine-Rich Protein 2 (PfHRP2) and Its Effect on the Performance of PfHRP2-Based Rapid Diagnostic Tests. J. Infect. Dis. 2005, 192, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Bharti, P.K.; Chandel, H.S.; Krishna, S.; Nema, S. Sequence variation in Plasmodium falciparum Histidine Rich Proteins 2 and 3 in Indian isolates: Implications for Malaria Rapid Diagnostic Test Performance. Sci. Rep. 2017, 7, 1308. [Google Scholar]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Gatton, M.L.; Barnwell, J.; Chiodini, P.; McCarthy, J.; Bell, D.; Cunningham, J. Plasmodium falciparum parasites lacking histidine-rich protein 2 and 3: A review and recommendations for accurate reporting. Malar. J. 2014, 13, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinyi, S.; Hayden, T.; Gamboa, D.; Torres, K.; Bendezu, J.; Abdallah, J.F.; Griffing, S.M.; Quezada, W.M.; Arrospide, N.; De Oliveira, A.M.E.; et al. Multiple genetic origins of histidine-rich protein 2 gene deletion in Plasmodium falciparum parasites from Peru. Sci. Rep. 2013, 3, 2797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo Solano, C.; Akinyi Okoth, S.; Abdallah, J.F.; Pava, Z.; Dorado, E.; Incardona, S.; Huber, C.S.; Macedo de Oliveira, A.; Bell, D.; Udhayakumar, V.; et al. Deletion of Plasmodium falciparum Histidine-Rich Protein 2 (pfhrp2) and Histidine-Rich Protein 3 (pfhrp3) Genes in Colombian Parasites. PLoS ONE 2015, 10, e0131576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorado, E.J.; Okoth, S.A.; Montenegro, L.M.; Diaz, G.; Barnwell, J.W.; Udhayakumar, V.; Solano, C.M. Genetic characterisation of Plasmodium falciparum isolates with deletion of the pfhrp2 and/or pfhrp3 genes in Colombia: The amazon region, a challenge for malaria diagnosis and control. PLoS ONE 2016, 11, e0163137. [Google Scholar] [CrossRef]
- Trouvay, M.; Palazon, G.; Berger, F.; Volney, B.; Blanchet, D.; Faway, E.; Donato, D.; Legrand, E.; Carme, B.; Musset, L. High Performance of Histidine-Rich Protein 2 Based Rapid Diagnostic Tests in French Guiana are Explained by the Absence of pfhrp2 Gene Deletion in P. falciparum. PLoS ONE 2013, 8, e74269. [Google Scholar] [CrossRef]
- Fontecha, G.; Mejía, R.E.; Banegas, E.; Ade, M.P.; Mendoza, L.; Ortiz, B.; Sabillón, I.; Alvarado, G.; Matamoros, G.; Pinto, A. Deletions of pfhrp2 and pfhrp3 genes of Plasmodium falciparum from Honduras, Guatemala and Nicaragua. Malar. J. 2018, 17, 320. [Google Scholar] [CrossRef]
- Kumar, N.; Pande, V.; Bhatt, R.M.; Shah, N.K.; Mishra, N.; Srivastava, B.; Valecha, N.; Anvikar, A.R. Genetic deletion of hrp2 and hrp3 in Indian Plasmodium falciparum population and false negative malaria rapid diagnostic test. Acta Trop. 2013, 125, 119–121. [Google Scholar] [CrossRef]
- Bharti, P.K.; Chandel, H.S.; Ahmad, A.; Krishna, S.; Udhayakumar, V.; Singh, N. Prevalence of pfhrp2 and/or pfhrp3 gene deletion in Plasmodium falciparum population in eight highly endemic states in India. PLoS ONE 2016, 11, e0157949. [Google Scholar] [CrossRef]
- Pati, P.; Dhangadamajhi, G.; Bal, M.; Ranjit, M. High proportions of pfhrp2 gene deletion and performance of HRP2-based rapid diagnostic test in Plasmodium falciparum field isolates of Odisha. Malar. J. 2018, 17, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Xing, H.; Zhao, Z.; Yang, Z.; Cao, Y.; Yan, G.; Sattabongkot, J.; Cui, L.; Fan, Q. Genetic diversity of Plasmodium falciparum histidine-rich protein 2 in the China-Myanmar border area. Acta Trop. 2015, 152, 26–31. [Google Scholar] [PubMed] [Green Version]
- Nima, M.K.; Hougard, T.; Hossain, M.E.; Kibria, M.G.; Mohon, A.N.; Johora, F.T.; Rahman, R.; Haque, R.; Alam, M.S. Case Report: A Case of Plasmodium falciparum hrp2 and hrp3 Gene Mutation in Bangladesh. Am. J. Trop. Med. Hyg. 2017, 97, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Koita, O.A.; Doumbo, O.K.; Ouattara, A.; Tall, L.K.; Konaré, A.; Diakité, M.; Diallo, M.; Sagara, I.; Masinde, G.L.; Doumbo, S.N.; et al. False-negative rapid diagnostic tests for malaria and deletion of the histidine-rich repeat region of the hrp2 gene. Am. J. Trop. Med. Hyg. 2012, 86, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurtz, N.; Fall, B.; Bui, K.; Pascual, A.; Fall, M.; Camara, C.; Diatta, B.; Fall, K.B.; Mbaye, P.S.; Diémé, Y.; et al. Pfhrp2 and pfhrp3 polymorphisms in Plasmodium falciparum isolates from Dakar, Senegal: Impact on rapid malaria diagnostic tests. Malar. J. 2013, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amoah, L.E.; Abankwa, J.; Oppong, A. Plasmodium falciparum histidine rich protein-2 diversity and the implications for PfHRP 2: Based malaria rapid diagnostic tests in Ghana. Malar. J. 2016, 15, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozycki, C.T.; Umulisa, N.; Rulisa, S.; Mwikarago, E.I.; Musabyimana, J.P.; Habimana, J.P.; Karema, C.; Krogstad, D.J. False-negative malaria rapid diagnostic tests in Rwanda: Impact of Plasmodium falciparum isolates lacking hrp2 and declining malaria transmission. Malar. J. 2017, 16, 123. [Google Scholar] [CrossRef] [Green Version]
- Gupta, H.; Matambisso, G.; Galatas, B.; Cisteró, P.; Nhamussua, L.; Simone, W.; Cunningham, J.; Rabinovitch, N.R.; Alonso, P.; Saute, F.; et al. Molecular surveillance of pfhrp2 and pfhrp3 deletions in Plasmodium falciparum isolates from Mozambique. Malar. J. 2017, 16, 416. [Google Scholar] [CrossRef] [Green Version]
- Menegon, M.; L’Episcopia, M.; Nurahmed, A.M.; Talha, A.A.; Nour, B.Y.M.; Severini, C. Identification of Plasmodium falciparum isolates lacking histidine-rich protein 2 and 3 in Eritrea. Infect. Genet. Evol. 2017, 55, 131–134. [Google Scholar] [CrossRef]
- Berhane, A.; Anderson, K.; Mihreteab, S.; Gresty, K.; Rogier, E.; Mohamed, S.; Hagos, F.; Embaye, G.; Chinorumba, A.; Zehaie, A.; et al. Major threat to malaria control programs by Plasmodium falciparum lacking histidine-rich protein 2, Eritrea. Emerg. Infect. Dis. 2018, 24, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Echeverry, D.F.; Nair, S.; Osorio, L.; Menon, S.; Murillo, C.; Anderson, T.J.C. Long term persistence of clonal malaria parasite Plasmodium falciparum lineages in the Colombian Pacific region. BMC Genet. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laban, N.M.; Kobayashi, T.; Hamapumbu, H.; Sullivan, D.; Mharakurwa, S.; Thuma, P.E.; Shiff, C.J.; Moss, W.J. Comparison of a PfHRP2-based rapid diagnostic test and PCR for malaria in a low prevalence setting in rural southern Zambia: Implications for elimination. Malar. J. 2015, 14, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parr, J.B.; Anderson, O.; Juliano, J.J.; Meshnick, S.R. Streamlined, PCR-based testing for pfhrp2- and pfhrp3-negative Plasmodium falciparum. Malar. J. 2018, 17, 137. [Google Scholar] [CrossRef]
- Berhane, A.; Russom, M.; Bahta, I.; Hagos, F.; Ghirmai, M.; Uqubay, S. Rapid diagnostic tests failing to detect Plasmodium falciparum infections in Eritrea: An investigation of reported false negative RDT results. Malar. J. 2017, 16, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Protocol for Estimating the Prevalence of pfhrp2/pfhrp3 Gene Deletions among Symptomatic Falciparum Patients with False-Negative RDT Results; World Health Organization: Geneva, Switzerland, 2018; Available online: http://www.who.int/malaria/publications/atoz/hrp2-deletion-protocol/en/ (accessed on 12 October 2018).
- Fontecha, G.; Pinto, A.; Escobar, D.; Matamoros, G.; Ortiz, B. Genetic variability of Plasmodium falciparum histidine-rich proteins 2 and 3 in Central America. Malar. J. 2019, 18, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.; Baker, J.; Andrews, K.T.; Gatton, M.L.; Bell, D.; Cheng, Q.; McCarthy, J. Effect of sequence variation in Plasmodium falciparum histidine-rich protein 2 on binding of specific monoclonal antibodies: Implications for rapid diagnostic tests for malaria. J. Clin. Microbiol. 2006, 44, 2773–2778. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; Ho, M.; Pelecanos, A.; Gatton, M.; Chen, N.; Abdullah, S.; Albertini, A.; Ariey, F.; Barnwell, J.; Bell, D.; et al. Global sequence variation in the histidine-rich proteins 2 and 3 of Plasmodium falciparum: Implications for the performance of malaria rapid diagnostic tests. Malar. J. 2010, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Gendrot, M.; Fawaz, R.; Dormoi, J.; Madamet, M.; Pradines, B. Genetic diversity and deletion of Plasmodium falciparum histidine-rich protein 2 and 3: A threat to diagnosis of P. falciparum malaria. Clin. Microbiol. Infect. 2018, 25, 580–585. [Google Scholar] [CrossRef]
- Ramutton, T.; Hendriksen, I.C.; Mwanga-Amumpaire, J.; Mtove, G.; Olaosebikan, R.; Tshefu, A.K.; Onyamboko, M.A.; Karema, C.; Maitland, K.; Gomes, E.; et al. Sequence variation does not confound the measurement of plasma PfHRP2 concentration in African children presenting with severe malaria. Malar. J. 2012, 11, 276. [Google Scholar] [CrossRef] [Green Version]
- WHO. T3: Test. Treat. Track. Scaling up Diagnostic Testing, Treatment and Surveillance for Malaria; WHO: Geneva, Switzerland, 2013; Available online: http://www.who.int/malaria/publications/atoz/t3_brochure/en/ (accessed on 12 October 2018).
- Watson, O.J.; Slater, H.C.; Verity, R.; Parr, J.B.; Mwandagalirwa, M.K.; Tshefu, A.; Meshnick, S.R.; Ghani, A.C. Modelling the drivers of the spread of Plasmodium falciparum hrp2 gene deletions in sub-Saharan Africa. Elife 2017, 6, e25008. [Google Scholar] [CrossRef]
- Shakely, D.; Elfving, K.; Aydin-Schmidt, B.; Msellem, M.I.; Morris, U.; Omar, R.; Weiping, X.; Petzold, M.; Greenhouse, B.; Baltzell, K.A.; et al. The usefulness of rapid diagnostic tests in the new context of low malaria transmission in Zanzibar. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranadive, N.; Kunene, S.; Darteh, S.; Ntshalintshali, N.; Nhlabathi, N.; Dlamini, N.; Chitundu, S.; Saini, M.; Murphy, M.; Soble, A.; et al. Limitations of rapid diagnostic testing in patients with suspected malaria: A diagnostic accuracy evaluation from Swaziland, a low-endemicity country aiming for malaria elimination. Clin. Infect. Dis. 2017, 64, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Agaba, B.B.; Yeka, A.; Nsobya, S.; Arinaitwe, E.; Nankabirwa, J.; Opigo, J.; Mbaka, P.; Lim, C.S.; Kalyango, J.N.; Karamagi, C.; et al. Systematic review of the status of pfhrp2 and pfhrp3 gene deletion, approaches and methods used for its estimation and reporting in Plasmodium falciparum populations in Africa: Review of published studies 2010–2019. Malar. J. 2019, 18, 355. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
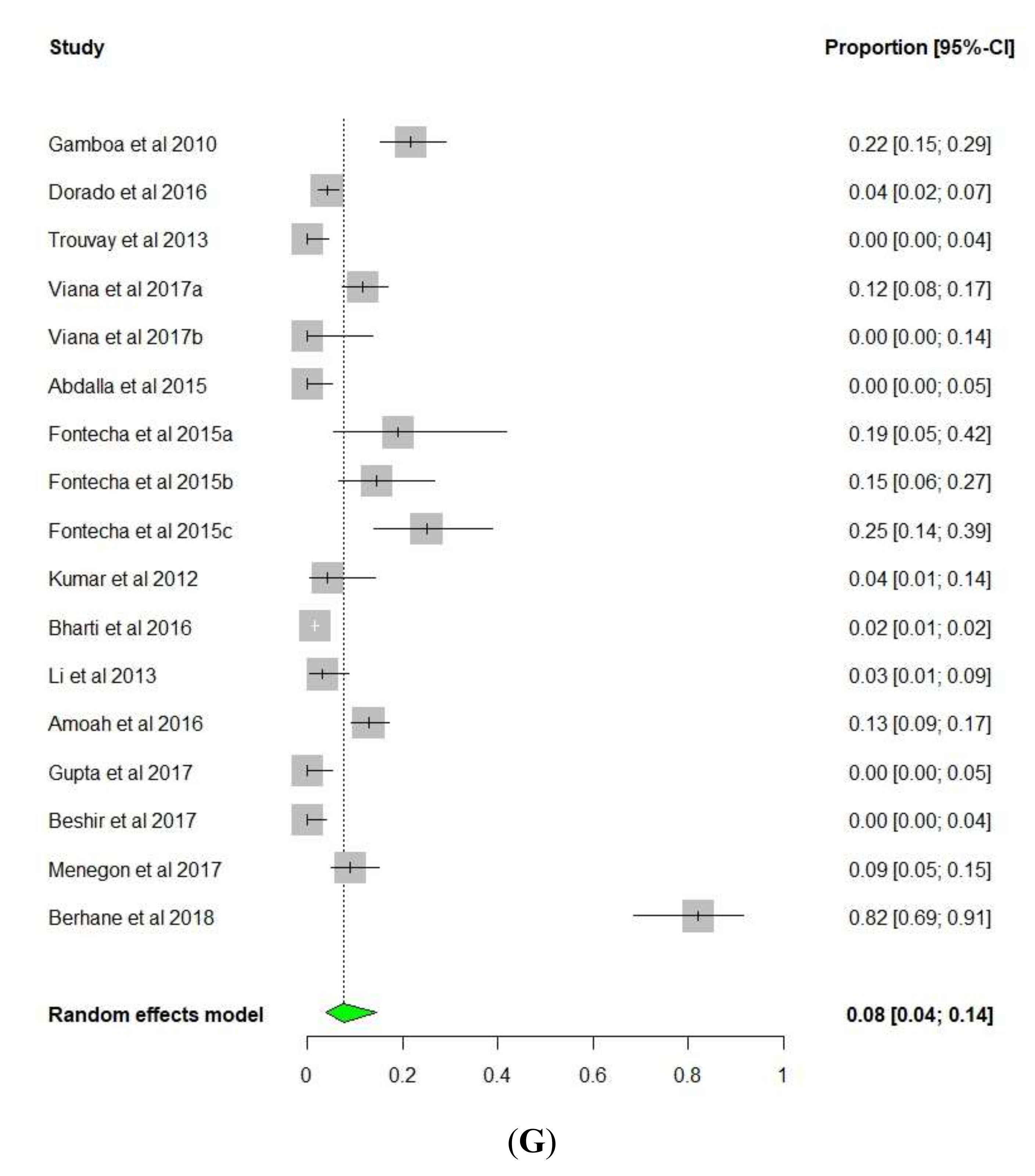{kind=link}
| Region | Country (Year of Sample Collection) | Initial Evidence | Gene Deletion Assays | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| RDT | Microscopy | pfhrp2-(Exon 1–2) | Upstream MAL7P1.230 | Downstream MAL7P1.228 | pfhrp3-(Exon 1–2) | Upstream MAL13P1.475 | Downstream MAL13P1.485 | Double Negative (pfhrp2 and pfhrp3) | Reference | ||
| South America | Peru (2003–2007) | D | D | 41% (61/148) | 88% (8/9) | 0 | 70% (103/148) | 55.6% (5/9) | 44.4% (4/9) | 21.6% (32/148) | [40] |
| Peru (1998–2001) | ND | ND | 20.7% (19/92) | 39.1% (36/92) | 14.1% (13/92) | ND | ND | ND | ND | [50] | |
| Peru (2003–2005) | ND | ND | 40.6(36/96) | 44.8(43/96) | 11.5% (11/96) | ND | ND | ND | ND | ||
| Colombia (1999–2009) | D | D | 18% (18/100) | 22% (22/100) | 1% (1/100) | 52% (52/100) | 55% (55/100) | 57% (57/100) | ND | [51] | |
| Colombia (2003–2012) | D | D | 4.1% (15/365) | 4.1% (15/365) | 0 | 43.0% (157/365) | 28.0% (102/365) | 41.6% (152/365) | 4.1% (15/365) | [52] | |
| French Guiana (2010–2011) | D | D | 0 | ND | ND | 7.4% (6/81) | ND | ND | 0 | [53] | |
| Brazil (2010–2012) | D | D | 13.6% (27/198) | 21.2% (42/198) | 0.5% (1/198) | 35.9% (71/198) | 20.7% (41/198) | 17.7% (35/198) | 11.6% (23/198) | [42] | |
| Bolivia (2010) | D | D | 4.0% (1/25) | 0 | 36.0% (9/25) | 68.0% (17/25) | 48.0% (12/25) | 56.0% (14/25) | NS | ||
| Central America | Honduras (2008–2009) | ND | D | 0 | 0 | 0 | 44.1% (30/68) | 47.1% (32/68) | 19.1% (13/68) | 0 | [39] |
| Guatemala, 2015 | ND | D | 14.3% (3/21) | 42.8% (9/21) | 0 | 90.5% (19/21) | 95.2% (20/21) | 100% (21/21) | 20% (11/55) | [54] | |
| Nicaragua, 2015 | ND | D | 30.9% (17/55) | 20.0% (11/55) | 0 | 87.3% (48/55) | 70.9% (39/55) | 56.4% (31/55) | 14.3% (3/21) | ||
| Honduras (2011–2017) | ND | D | 25.0% (13/52) | 21.1% (11/52) | 1.9% (1/52) | 96.2% (50/52) | 55.8% (29/52) | 51.9% (27/52) | 25% (13/52) | ||
| Asia | India 2010 | D | D | 4.2% (2/48) | 4.2% (2/48) | 4.2% (2/48) | 4.2% (2/48) | 4.2% (2/48) | 4.2% (2/48) | 4.2% (2/48) | [55] |
| India 2014 | D | D | 2.4% (36/1571) | 1.7% (28/1571) | 1.6% (26/1571) | 1.7% (27/1571) | 0.6% (10/1571) | 1.4% (22/1571) | 1.6% (25/1571) | [56] | |
| India (2018) | D | D | 3.6% (38/1058) | 0.5% (5/1058) | 0.4% (4/1058) | 2.3% (24/1058) | 0.1% (1/1058) | 0.3% (3/1058) | 1.6% (17/1058) | [57] | |
| China–Myanmar (2011–2012) | D | D | 4.1% (4/97) | 4.1% (4/97) | 4.1% (4/97) | 3.1% (3/97) | ND | ND | 3.1% (3/97) | [58] | |
| Bangladesh 2017 | D | NS | CS | CS | CS | CS | CS | CS | CS | [59] | |
| Africa | Mali (1996) | D | D | 2.1% (10/480) | ND | ND | ND | ND | ND | ND | [60] |
| Senegal (2009–2012) | D | D | 2.4% (3/125) | ND | ND | 12.8% (16/125) | ND | ND | NS | [61] | |
| Ghana (2015) | D | D | 13.5% (29/288) | C | ND | 16.7% (48/288) | ND | ND | 12.9% (37/288) | [62] | |
| Rwanda (2014–2015) | D | D | 23.1% (32/138) | ND | ND | ND | ND | ND | ND | [63] | |
| Mozambique (2010–2016) | D | D | 1.5% (1/69) | ND | ND | 0 | ND | ND | 0 | [64] | |
| Kenya | D | D | 9% (8/89) | ND | ND | 1.1% (1/89) | ND | ND | 0 | [12] | |
| Eritrea (2013–2014) | ND | D | 9.7% (914/144) | 23.6% (34/144) | 3.5% (5/144) | 43% (62/144) | 55.6% (80/155) | 23.6% (34/144) | 9% (13/144) | [65] | |
| Eritrea (2016) | D | D | 62% (31/50) | 78% (39/50) | 0 | 82% (41/50) | 64% (32/50) | 78% (39/50) | 82% (41/50) | [66] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyataya, J.; Waitumbi, J.; Mobegi, V.A.; Noreddin, A.; El Zowalaty, M.E. Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions and Their Implications in Malaria Control. Diseases 2020, 8, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases8020015
Nyataya J, Waitumbi J, Mobegi VA, Noreddin A, El Zowalaty ME. Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions and Their Implications in Malaria Control. Diseases. 2020; 8(2):15. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases8020015
Chicago/Turabian StyleNyataya, Josphat, John Waitumbi, Victor A. Mobegi, Ayman Noreddin, and Mohamed E. El Zowalaty. 2020. "Plasmodium falciparum Histidine-Rich Protein 2 and 3 Gene Deletions and Their Implications in Malaria Control" Diseases 8, no. 2: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/diseases8020015