In Vivo Metabolism of [1,6-13C2]Glucose Reveals Distinct Neuroenergetic Functionality between Mouse Hippocampus and Hypothalamus

Abstract
:1. Introduction
2. Results
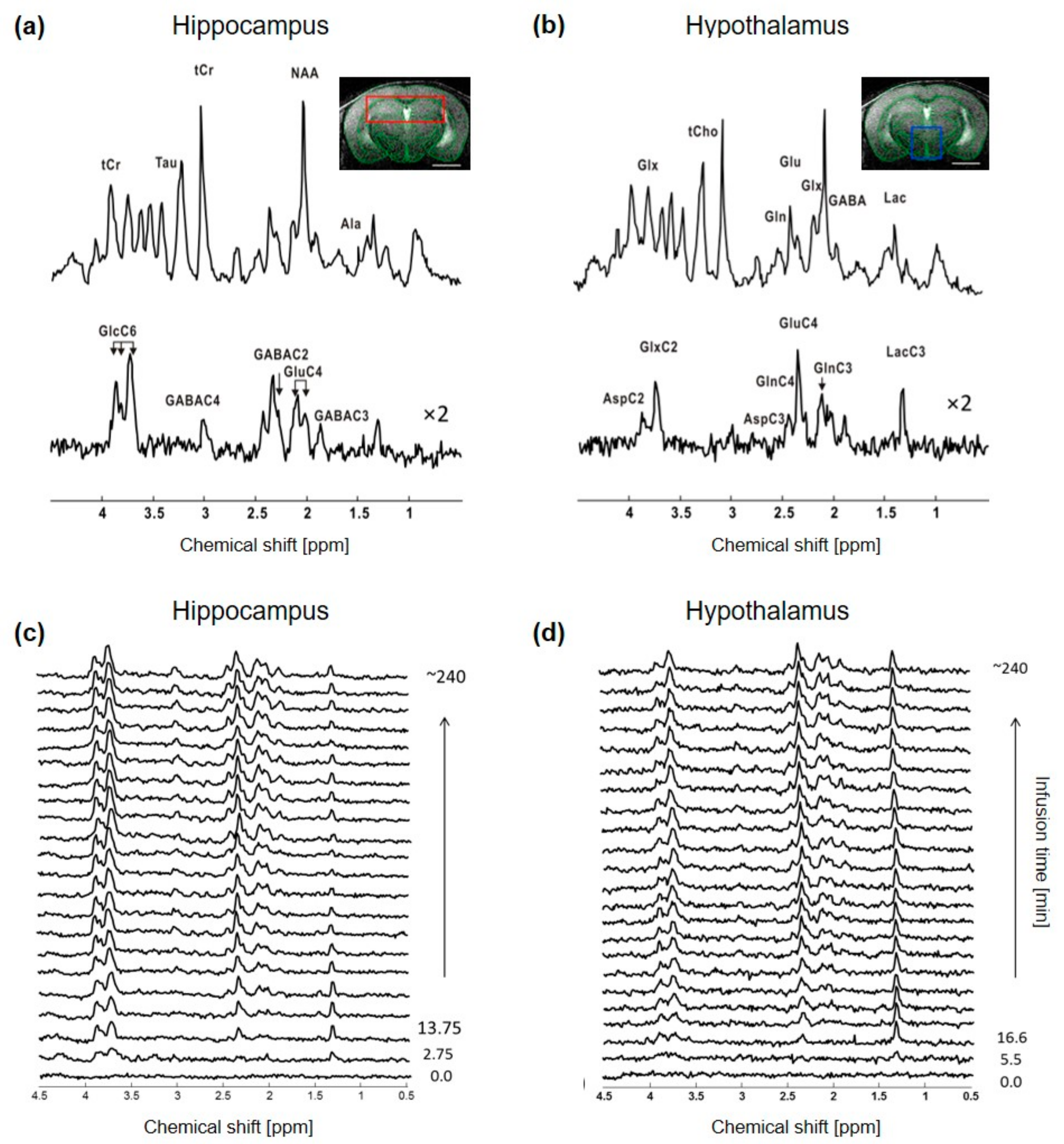
2.1. 1H-[13C]-MRS Data Comparison between Hypothalamus and Hippocampus at 14.1T
2.2. Metabolic Differences in Neurochemical Profile between Hippocampus and Hypothalamus
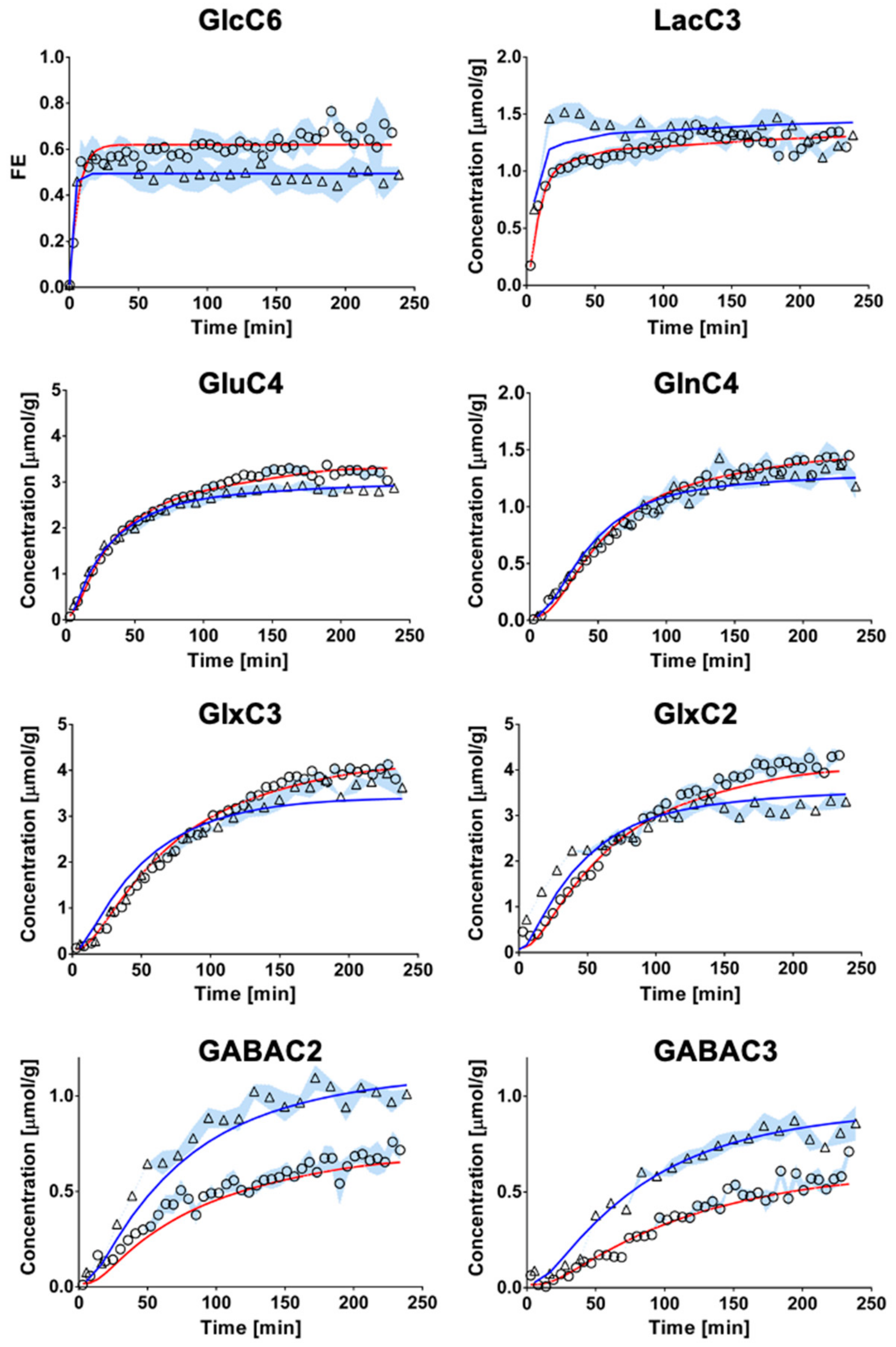
2.3. Mathematical Modeling of Hippocampal and Hypothalamic Metabolism
3. Discussion
3.1. Protocol Optimization Allows Simultaneous Assessment of Glycolytic and Oxidative Glucose Metabolism
3.2. Major Metabolic Differences with Current Literature
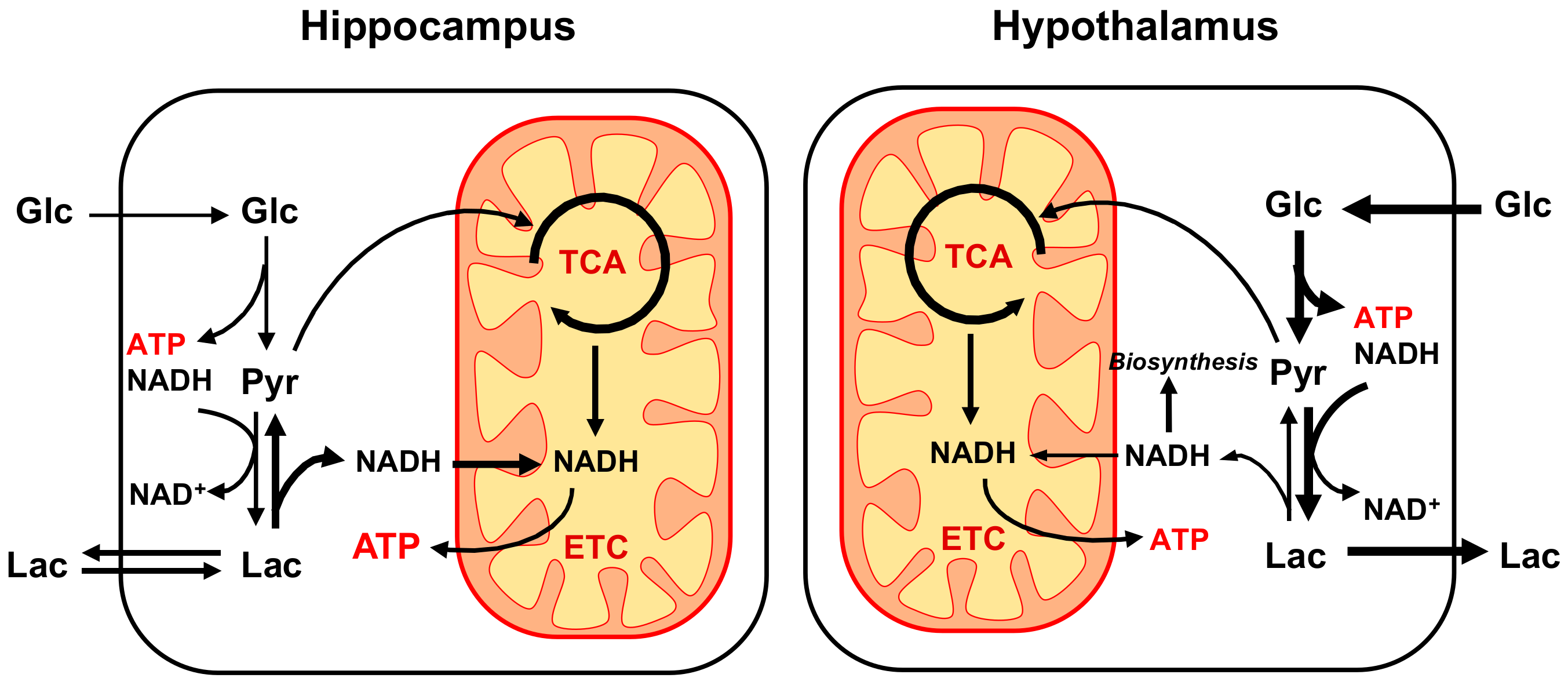
3.3. Biological Functionality Underlying Regional Differences in Energy Metabolism
3.4. General Concerns about the Protocol and Anesthesia
4. Material and Methods
4.1. Animals
4.2. MR Hardware
4.3. MR Method
4.4. 1H-[13C]-MRS Spectra Analysis
4.5. FE Measurements of Blood Samples
4.6. Mathematical Modeling of Metabolic Fluxes
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raichle, M.E.; Gusnard, D.A. Appraising the brain’s energy budget. Proc. Natl. Acad. Sci. USA 2002, 99, 10237–10239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinridders, A.; Ferris, H.A.; Reyzer, M.L.; Rath, M.; Soto, M.; Manier, M.L.; Spraggins, J.M.; Yang, Z.; Stanton, R.C.; Caprioli, R.M.; et al. Regional differences in brain glucose metabolism determined by imaging mass spectrometry. Mol. Metab. 2018, 12, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Zangrandi, L.; Schwarzer, C.; Gnaiger, E. Differences in mitochondrial function in homogenated samples from healthy and epileptic specific brain tissues revealed by high-resolution respirometry. Mitochondrion 2015, 25, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Bao, A.-M.; Meynen, G.; Swaab, D. The stress system in depression and neurodegeneration: Focus on the human hypothalamus. Brain Res. Rev. 2008, 57, 531–553. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 1999, 22, 105–122. [Google Scholar] [CrossRef] [Green Version]
- Routh, V.H.; Hao, L.; Santiago, A.M.; Sheng, Z.; Zhou, C. Hypothalamic glucose sensing: Making ends meet. Front. Syst. Neurosci. 2014, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Lizarbe, B.; Cherix, A.; Duarte, J.M.N.; Cardinaux, J.-R.; Gruetter, R. High-fat diet consumption alters energy metabolism in the mouse hypothalamus. Int. J. Obes. 2019, 43, 1295–1304. [Google Scholar] [CrossRef]
- Squire, L.R.; Genzel, L.; Wixted, J.T.; Morris, R.G. Memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021766. [Google Scholar] [CrossRef]
- Jeffery, K. The hippocampus: From memory, to map, to memory map. Trends Neurosci. 2018, 41, 64–66. [Google Scholar] [CrossRef]
- Le Merre, P.; Esmaeili, V.; Charrière, E.; Galan, K.; Salin, P.-A.; Petersen, C.C.; Crochet, S. Reward-based learning drives rapid sensory signals in medial prefrontal cortex and dorsal hippocampus necessary for goal-directed behavior. Neuron 2018, 97, 83–91.e5. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. Stress-induced remodeling of hippocampal CA3 pyramidal neurons. Brain Res. 2016, 1645, 50–54. [Google Scholar] [CrossRef]
- Sapolsky, R. Glucocorticoid toxicity in the hippocampus: Reversal by supplementation with brain fuels. J. Neurosci. 1986, 6, 2240–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlynarik, V.; Cudalbu, C.R.; Xin, L.; Gruetter, R. 1H NMR spectroscopy of rat brain in vivo at 14.1Tesla: Improvements in quantification of the neurochemical profile. J. Magn. Reson. 2008, 194, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, H.; Xin, L.; Gruetter, R.; Mlynarik, V. Localized single-voxel magnetic resonance spectroscopy, water suppression, and novel approaches for ultrashort echo-time measurements. In Magnetic Resonance Spectroscopy; Stagg, C.J., Rothman, D.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 15–30. [Google Scholar]
- Gruetter, R.; Novotny, E.J.; Boulware, S.D.; Mason, G.F.; Rothman, D.L.; Shulman, G.I.; Prichard, J.W.; Shulman, R.G. Localized 13C NMR Spectroscopy in the Human Brain of Amino Acid Labeling from d-[1-13C]Glucose. J. Neurochem. 2002, 63, 1377–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, L.; Lanz, B.; Lei, H.; Gruetter, R. Assessment of metabolic fluxes in the mouse brain in vivo using 1H-[13C] NMR spectroscopy at 14.1 Tesla. J. Cereb. Blood Flow Metab. 2015, 35, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanz, B.; Gruetter, R.; Duarte, J.M.N. Metabolic flux and compartmentation analysis in the brain in vivo. Front. Endocrinol. (Lussane) 2013, 4, 156. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.M.N.; Lanz, B.; Gruetter, R. Compartmentalized cerebral metabolism of [1,6-13C]glucose determined by in vivo13C NMR spectroscopy at 14.1 T. Front. Neuroenerg. 2011, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Lizarbe, B.; Lei, H.; Duarte, J.M.N.; Lanz, B.; Cherix, A.; Gruetter, R. Feasibility of in vivo measurement of glucose metabolism in the mouse hypothalamus by1H-[13C] MRS at 14.1T. Magn. Reson. Med. 2018, 80, 874–884. [Google Scholar] [CrossRef] [Green Version]
- Cherix, A.; Donati, G.; Lizarbe, B.; Lanz, B.; Poitry-Yamate, C.; Lei, H.; Gruetter, R. Excitatory/inhibitory neuronal metabolic balance in mouse hippocampus upon infusion of [U-13C6]glucose. J. Cereb. Blood Flow Metab. 2020, 632, 0271678X20910535. [Google Scholar] [CrossRef]
- Cherix, A.; Larrieu, T.; Grosse, J.; Rodrigues, J.; McEwen, B.; Nasca, C.; Gruetter, R.; Sandi, C. Metabolic signature in nucleus accumbens for anti-depressant-like effects of acetyl-L-carnitine. Elife 2020, 9, e50631. [Google Scholar] [CrossRef]
- Lei, H.; Poitry-Yamate, C.; Preitner, F.; Thorens, B.; Gruetter, R. Neurochemical profile of the mouse hypothalamus using in vivo 1 H MRS at 14.1T. NMR Biomed. 2010, 23, 578–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg-Johnsen, J.; Langmoen, I. Mechanisms concerned in the direct effect of isoflurane on rat hippocampal and human neocortical neurons. Brain Res. 1990, 507, 28–34. [Google Scholar] [CrossRef]
- Patel, A.B.; De Graaf, R.A.; Mason, G.F.; Rothman, D.L.; Shulman, R.G.; Behar, K.L. The contribution of GABA to glutamate/glutamine cycling and energy metabolism in the rat cortex in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 5588–5593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, M.; Lanz, B.; Poitry-Yamate, C.; Romero, J.F.; Berset, C.M.; Cudalbu, C.; Gruetter, R. In vivo 13C MRS in the mouse brain at 14.1 Tesla and metabolic flux quantification under infusion of [1,6-13C2]glucose. J. Cereb. Blood Flow Metab. 2018, 38, 1701–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuke, S.; Kubota-Sakashita, M.; Kasahara, T.; Shigeyoshi, Y.; Kato, T. Regional variation in mitochondrial DNA copy number in mouse brain. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 270–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.V.; Jakobsen, E.; Waagepetersen, H.S.; Aldana, B.I. Distinct differences in rates of oxygen consumption and ATP synthesis of regionally isolated non-synaptic mouse brain mitochondria. J. Neurosci. Res. 2019, 97, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Hertz, L.; Peng, L.; Dienel, G.A. Energy metabolism in astrocytes: High rate of oxidative metabolism and spatiotemporal dependence on glycolysis/glycogenolysis. J. Cereb. Blood Flow Metab. 2006, 27, 219–249. [Google Scholar] [CrossRef]
- McKenna, M.C.; Waagepetersen, H.S.; Schousboe, A.; Sonnewald, U. Neuronal and astrocytic shuttle mechanisms for cytosolic-mitochondrial transfer of reducing equivalents: Current evidence and pharmacological tools. Biochem. Pharmacol. 2006, 71, 399–407. [Google Scholar] [CrossRef]
- McKenna, M.C.; Tildon, J.T.; Stevenson, J.H.; Huang, X.; Kingwell, K.G. Regulation of mitochondrial and cytosolic malic enzymes from cultured rat brain astrocytes. Neurochem. Res. 1995, 20, 1491–1501. [Google Scholar] [CrossRef]
- Erö, C.; Gewaltig, M.-O.; Keller, D.; Markram, H. A cell atlas for the mouse brain. Front. Neuroinform. 2018, 12, 84. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, J. Membrane breakdown in acute and chronic neurodegeneration: Focus on choline-containing phospholipids. J. Neural Transm. 2000, 107, 1027–1063. [Google Scholar] [CrossRef] [PubMed]
- Grill, H.J.; Hayes, M.R. Hindbrain neurons as an essential hub in the neuroanatomically distributed control of energy balance. Cell Metab. 2012, 16, 296–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S.; Reagan, L.P. Glucose transporter expression in the central nervous system: Relationship to synaptic function. Eur. J. Pharmacol. 2004, 490, 13–24. [Google Scholar] [CrossRef]
- Lizarbe, B.; Cherix, A.; Gruetter, R. In vivo heteronuclear magnetic resonance spectroscopy. Methods Mol. Biol. 2018, 1718, 169–187. [Google Scholar] [CrossRef] [Green Version]
- Gruetter, R.; Tkáč, I. Field mapping without reference scan using asymmetric echo-planar techniques. Magn. Reson. Med. 2000, 43, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Mlynárik, V.; Gambarota, G.; Frenkel, H.; Gruetter, R. Localized short-echo-time proton MR spectroscopy with full signal-intensity acquisition. Magn. Reson. Med. 2006, 56, 965–970. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Mlynárik, V.; Lanz, B.; Frenkel, H.; Gruetter, R. 1H-[13C] NMR spectroscopy of the rat brain during infusion of [2-13C] acetate at 14.1 T. Magn. Reson. Med. 2010, 64, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Provencher, S.W. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001, 14, 260–264. [Google Scholar] [CrossRef]
- Sonnay, S.; Duarte, J.M.N.; Just, N.; Gruetter, R. Compartmentalised energy metabolism supporting glutamatergic neurotransmission in response to increased activity in the rat cerebral cortex: A 13C MRS study in vivo at 14.1 T. 1. J. Celeb. Blood Flow Metab. 2016, 36, 928–940. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flux | Hippocampus | Hypothalamus |
|---|---|---|
| VTCA | 0.75 ± 0.05 | 0.87 ± 0.11 |
| Vx | 0.50 ± 0.08 | 0.20 ± 0.06 **** |
| VNT | 0.14 ± 0.01 | 0.14 ± 0.05 |
| Vdilin | 0.23 ± 0.06 | 0.00 ± 0.06 **** |
| Vdilout | 0.21 ± 0.02 | 0.30 ± 0.02 **** |
| Vdilg | 0.06 ± 0.01 | 0.00 ± 0.00 **** |
| VGABA | 0.06 ± 0.01 | 0.09 ± 0.02 **** |
| VPC | 0.005 ± 0.005 | 0.010 ± 0.010 |
| Vexi | 0.01 ± 0.01 | 0.02 ± 0.02 |
| CMRGlc | 0.29 ± 0.02 | 0.47 ± 0.06 **** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cherix, A.; Sonti, R.; Lanz, B.; Lei, H. In Vivo Metabolism of [1,6-13C2]Glucose Reveals Distinct Neuroenergetic Functionality between Mouse Hippocampus and Hypothalamus. Metabolites 2021, 11, 50. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010050
Cherix A, Sonti R, Lanz B, Lei H. In Vivo Metabolism of [1,6-13C2]Glucose Reveals Distinct Neuroenergetic Functionality between Mouse Hippocampus and Hypothalamus. Metabolites. 2021; 11(1):50. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010050
Chicago/Turabian StyleCherix, Antoine, Rajesh Sonti, Bernard Lanz, and Hongxia Lei. 2021. "In Vivo Metabolism of [1,6-13C2]Glucose Reveals Distinct Neuroenergetic Functionality between Mouse Hippocampus and Hypothalamus" Metabolites 11, no. 1: 50. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010050