
Cord Blood Metabolome and BMI Trajectory from Birth to Adolescence: A Prospective Birth Cohort Study on Early Life Biomarkers of Persistent Obesity

 , and
, and
Abstract
:1. Introduction
2. Results
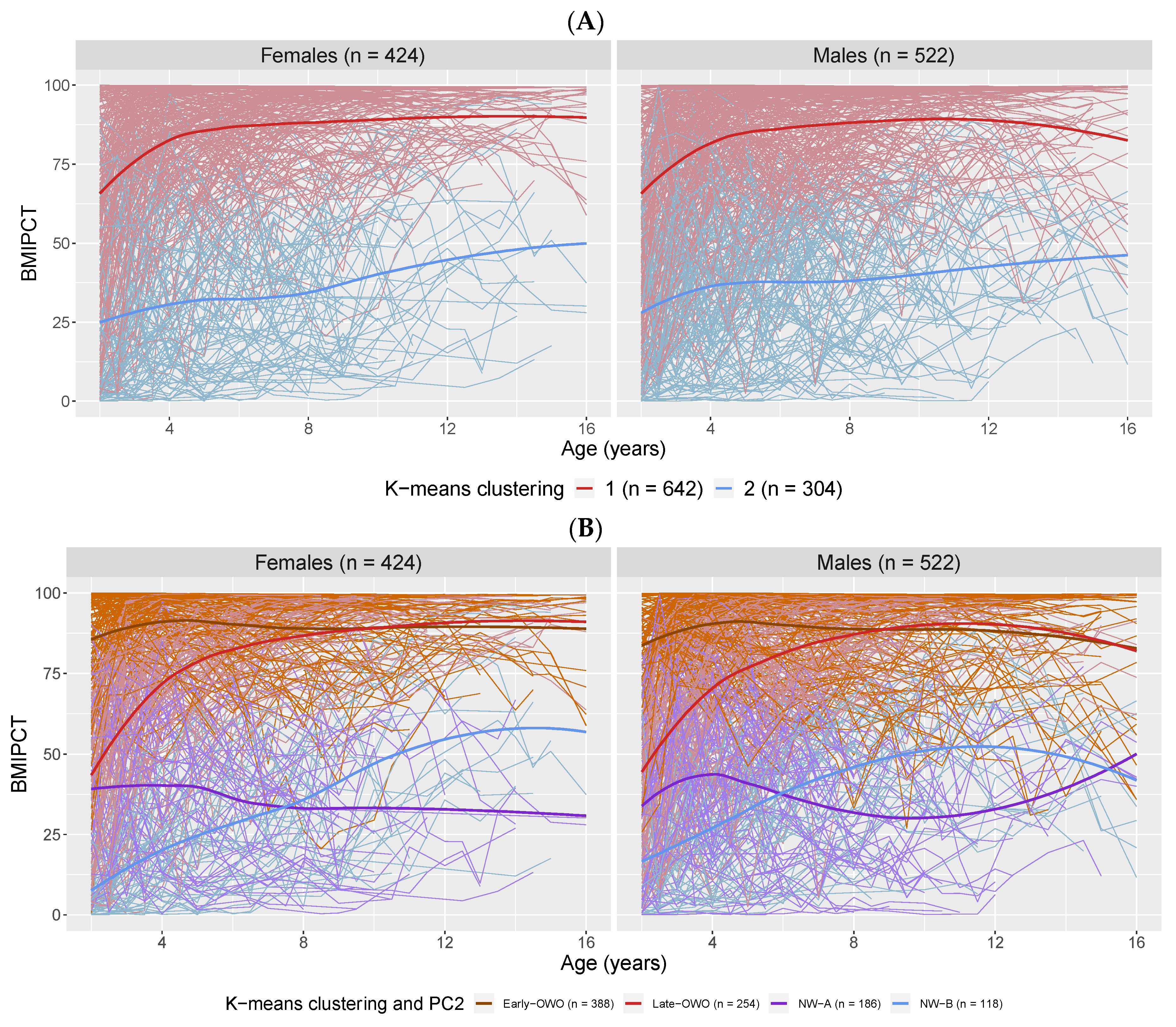
2.1. Longitudinal Trajectory Analysis: Categorizing Longitudinal BMI Trajectories
2.2. Longitudinal Trajectory Analysis: Metabolite Modules and BMI Trajectory Association
2.3. Longitudinal Trajectory Analysis: Sensitivity Analysis
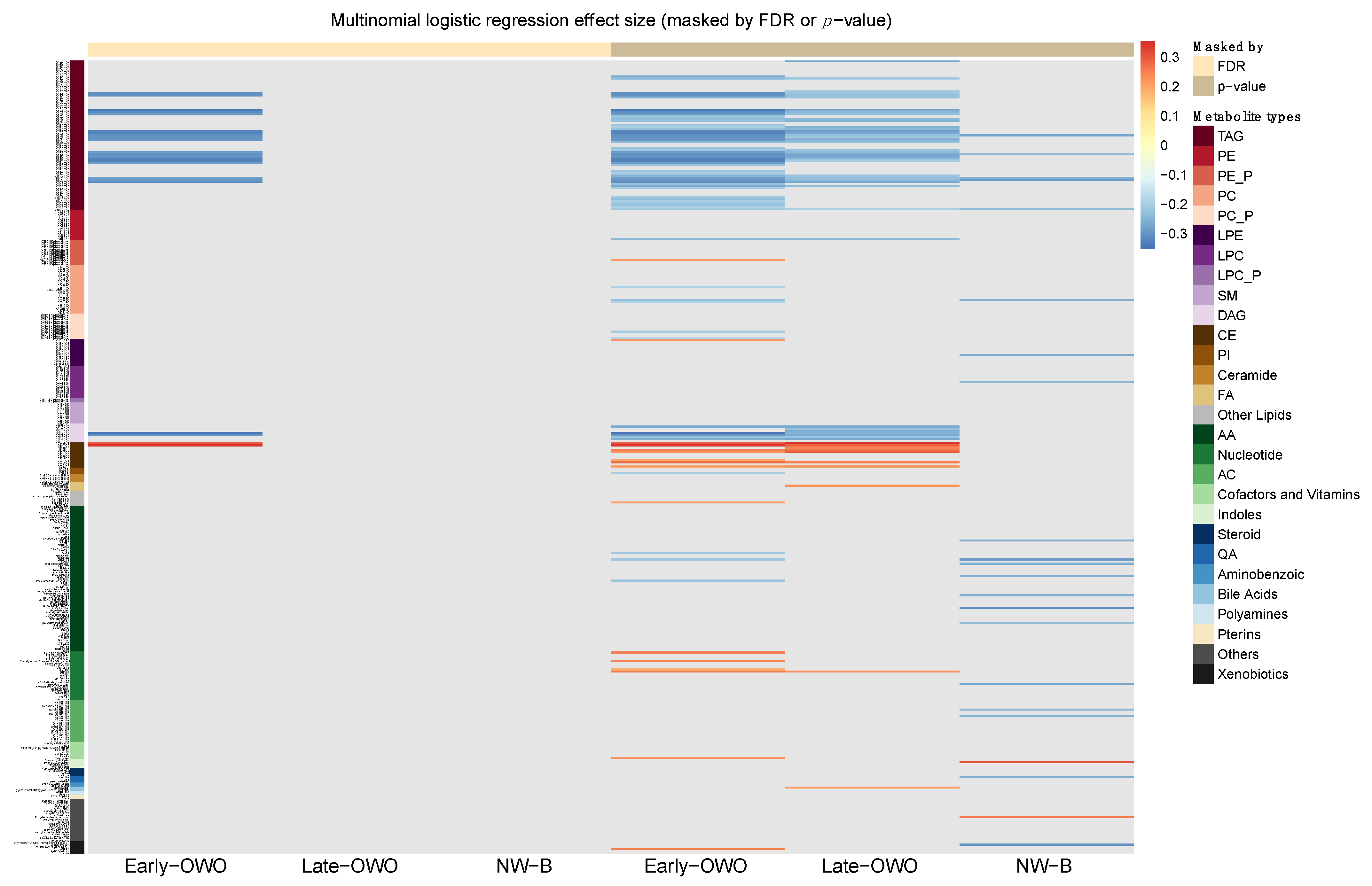
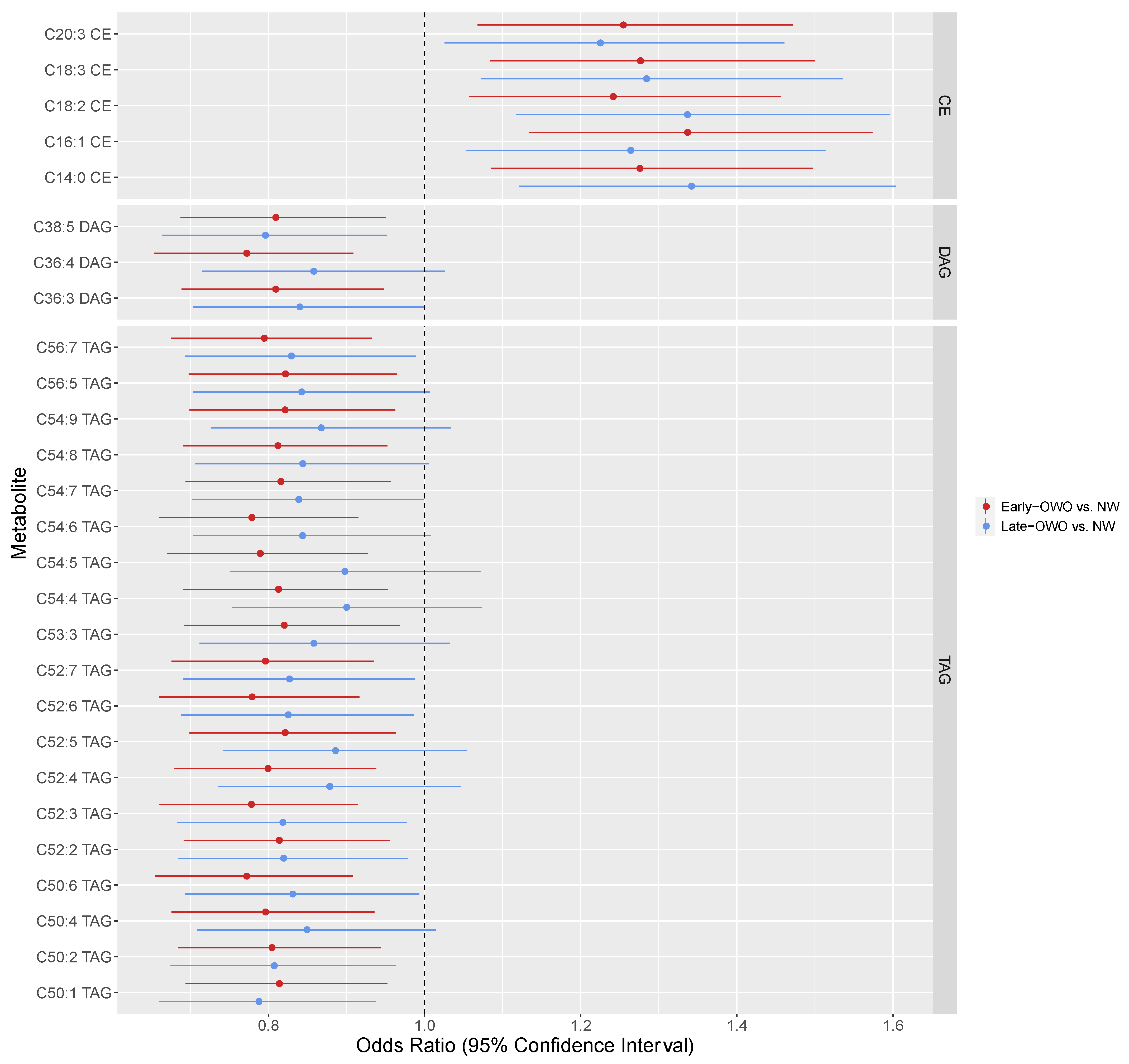
2.4. Time-Window Specific Analysis: Individual Metabolites
2.5. Time-Window Specific Analysis: Metabolite Modules
2.6. Time-Window Specific Analysis: Sensitivity Analyses
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Cord Plasma Metabolites
4.3. Statistical Analyses
4.3.1. Longitudinal Trajectory Analysis: Categorizing Longitudinal BMI Trajectories
4.3.2. Longitudinal Trajectory Analysis: Metabolite Modules and BMI Trajectory Association
4.3.3. Longitudinal Trajectory Analysis: Sensitivity Analysis
4.3.4. Time-window Specific Analysis: Individual Metabolites
4.3.5. Time-Window Specific Analysis: Metabolite Modules
4.3.6. Time-Window Specific Analysis: Sensitivity Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Disclaimer
Abbreviations
References
- Brown, C.L.; Halvorson, E.E.; Cohen, G.M.; Lazorick, S.; Skelton, J.A. Addressing Childhood Obesity: Opportunities for Prevention. Pediatr. Clin. N. Am. 2015, 62, 1241–1261. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.S.; Mulder, C.; Twisk, J.W.; van Mechelen, W.; Chinapaw, M.J. Tracking of childhood overweight into adulthood: A systematic review of the literature. Obes. Rev. 2008, 9, 474–488. [Google Scholar] [CrossRef]
- Geserick, M.; Vogel, M.; Gausche, R.; Lipek, T.; Spielau, U.; Keller, E.; Pfäffle, R.; Kiess, W.; Körner, A. Acceleration of BMI in Early Childhood and Risk of Sustained Obesity. N. Engl. J. Med. 2018, 379, 1303–1312. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, L.; Wu, Y.; Wilson, R.F.; Weston, C.; Fawole, O.; Bleich, S.N.; Cheskin, L.J.; Showell, N.N.; Lau, B.D.; et al. What childhood obesity prevention programmes work? A systematic review and meta-analysis. Obes. Rev. 2015, 16, 547–565. [Google Scholar] [CrossRef]
- Kadakia, R.; Scholtens, D.M.; Rouleau, G.W.; Talbot, O.; Ilkayeva, O.R.; George, T.; Josefson, J.L. Cord Blood Metabolites Associated with Newborn Adiposity and Hyperinsulinemia. J. Pediatr. 2018, 203, 144–149. [Google Scholar] [CrossRef]
- Kadakia, R.; Talbot, O.; Kuang, A.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Cord Blood Metabolomics: Association with Newborn Anthropometrics and C-Peptide across Ancestries. J. Clin. Endocrinol. Metab. 2019, 104, 4459–4472. [Google Scholar] [CrossRef] [PubMed]
- Perng, W.; Rifas-Shiman, S.L.; McCulloch, S.; Chatzi, L.; Mantzoros, C.; Hivert, M.-F.; Oken, E. Associations of cord blood metabolites with perinatal characteristics, newborn anthropometry, and cord blood hormones in project viva. Metabolism 2017, 76, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ivorra, C.; García-Vicent, C.; Chaves, F.J.; Monleón, D.; Morales, J.M.; Lurbe, E. Metabolomic profiling in blood from umbilical cords of low birth weight newborns. J. Transl. Med. 2012, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-P.; Reichetzeder, C.; Prehn, C.; Yin, L.-H.; Yun, C.; Zeng, S.; Chu, C.; Adamski, J.; Hocher, B. Cord Blood Lysophosphatidylcholine 16: 1 is Positively Associated with Birth Weight. Cell. Physiol. Biochem. 2018, 45, 614–624. [Google Scholar] [CrossRef]
- Isganaitis, E.; Rifas-Shiman, S.L.; Oken, E.; Dreyfuss, J.; Gall, W.; Gillman, M.W.; Patti, M.-E. Associations of cord blood metabolites with early childhood obesity risk. Int. J. Obes. 2015, 39, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrow, P.; Maguire, R.; Murphy, S.K.; Belcher, S.; Hoyo, C. Elevated metabolites of acetaminophen in cord blood of children with obesity. Pediatr. Obes. 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- La Torre, D.; Seppänen-Laakso, T.; Larsson, H.E.; Hyötyläinen, T.; Ivarsson, S.A.; Lernmark, Å.; Oresic, M.; the DiPiS Study Group. Decreased cord-blood phospholipids in young age-at-onset type 1 diabetes. Diabetes 2013, 62, 3951–3956. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weihrauch-Blüher, S.; Schwarz, P.; Klusmann, J.H. Childhood obesity: Increased risk for cardiometabolic disease and cancer in adulthood. Metabolism 2019, 92, 147–152. [Google Scholar] [CrossRef]
- Warensjö, E.; Öhrvall, M.; Vessby, B. Fatty acid composition and estimated desaturase activities are associated with obesity and lifestyle variables in men and women. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Duncan, R.E.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Triacylglycerol metabolism in adipose tissue. Future Lipidol. 2007, 2, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sulaiti, H.; Diboun, I.; Banu, S.; Al-Emadi, M.; Amani, P.; Harvey, T.M.; Dömling, A.S.; Latiff, A.; Elrayess, M.A. Triglyceride profiling in adipose tissues from obese insulin sensitive, insulin resistant and type 2 diabetes mellitus individuals. J. Transl. Med. 2018, 16, 175. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Shen, S.; Price, M.J.; Lu, J.; Sumilo, D.; Kuang, Y.; Manolopoulos, K.; Xia, H.; Qiu, X.; Cheng, K.K.; et al. Glucose, Insulin, and Lipids in Cord Blood of Neonates and Their Association with Birthweight: Differential Metabolic Risk of Large for Gestational Age and Small for Gestational Age Babies. J. Pediatr. 2020, 220, 64–72.e2. [Google Scholar] [CrossRef]
- Geraghty, A.A.; Alberdi, G.; O’Sullivan, E.J.; O’Brien, E.C.; Crosbie, B.; Twomey, P.; McAuliffe, F.M. Maternal Blood Lipid Profile during Pregnancy and Associations with Child Adiposity: Findings from the ROLO Study. PLoS ONE 2016, 11, e0161206. [Google Scholar] [CrossRef] [Green Version]
- Schaefer-Graf, U.M.; Graf, K.; Kulbacka, I.; Kjos, S.L.; Dudenhausen, J.; Vetter, K.; Herrera, E. Maternal lipids as strong determinants of fetal environment and growth in pregnancies with gestational diabetes mellitus. Diabetes Care 2008, 31, 1858–1863. [Google Scholar] [CrossRef] [Green Version]
- Wahl, S.; Yu, Z.; Kleber, M.; Singmann, P.; Holzapfel, C.; He, Y.; Mittelstrass, K.; Polonikov, A.; Prehn, C.; Römisch-Margl, W.; et al. Childhood obesity is associated with changes in the serum metabolite profile. Obes. Facts 2012, 5, 660–670. [Google Scholar] [CrossRef]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.; Delany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Stephens, F.B.; Constantin-Teodosiu, D.; Greenhaff, P. New insights concerning the role of carnitine in the regulation of fuel metabolism in skeletal muscle. J. Physiol. 2007, 581 Pt 2, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Meyer zu Heringdorf, D.; Jakobs, K.H. Lysophospholipid receptors: Signalling, pharmacology and regulation by lysophospholipid metabolism. Biochim. Biophys. Acta 2007, 1768, 923–940. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.; Ye, R.D. The lysophospholipid receptor G2A activates a specific combination of G proteins and promotes apoptosis. J. Biol. Chem. 2003, 278, 14379–14386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drzazga, A.; Sowińska, A.; Koziołkiewicz, M. Lysophosphatidylcholine and lysophosphatidylinosiol—Novel promissing signaling molecules and their possible therapeutic activity. Acta Pol. Pharm. 2014, 71, 887–899. [Google Scholar]
- Hellmuth, C.; Uhl, O.; Standl, M.; Demmelmair, H.; Heinrich, J.; Koletzko, B.; Thiering, E. Cord Blood Metabolome Is Highly Associated with Birth Weight, but Less Predictive for Later Weight Development. Obes. Facts 2017, 10, 85–100. [Google Scholar] [CrossRef]
- Sandall, J.; Tribe, R.M.; Avery, L.; Mola, G.; Visser, G.H.; Homer, C.S.; Gibbons, D.; Kelly, N.; Kennedy, H.P.; Kidanto, H.; et al. Short-term and long-term effects of caesarean section on the health of women and children. Lancet 2018, 392, 1349–1357. [Google Scholar] [CrossRef]
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Divall, S.; Radovick, S.; Paige, D.; Ning, Y.; Chen, Z.; Ji, Y.; Hong, X.; Walker, S.O.; Caruso, D.; et al. Preterm birth and random plasma insulin levels at birth and in early childhood. JAMA 2014, 311, 587–596. [Google Scholar] [CrossRef]
- Wang, G.; Sun, Q.; Liang, L.; Clash, C.; Zhang, C.; Hong, X.; Ji, Y.; Radovick, S.; Pearson, C.; Bartell, T.R.; et al. Inter-generational link of obesity in term and preterm births: Role of maternal plasma acylcarnitines. Int. J. Obes. 2019, 43, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.A.; Ordovas, J.M.; Eckfeldt, J.H.; Tracy, R.; Boerwinkle, E.; Lalouel, J.-M.; Printz, M. Guidelines of the National Heart, Lung, and Blood Institute Working Group on Blood Drawing, Processing, and Storage for Genetic Studies. Am. J. Epidemiol. 1997, 144, 437–441, Erratum in 1997, 145, 570. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted metabolomics. Curr. Protoc. Mol. Biol. 2012, 98, 30.2.1–30.2.24. [Google Scholar] [CrossRef] [PubMed]
- Rhee, E.P.; Cheng, S.; Larson, M.; Walford, G.A.; Lewis, G.D.; McCabe, E.; Yang, E.; Farrell, L.; Fox, C.S.; O’Donnell, C.J.; et al. Lipid profiling identifies a triacylglycerol signature of insulin resistance and improves diabetes prediction in humans. J. Clin. Investig. 2011, 121, 1402–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Health Statistics. CDC Growth Charts, United States. 2000. Available online: http://www.cdc.gov/growthcharts/ (accessed on 21 June 2019).
- Hong, Y.H.; Chung, S. Small for gestational age and obesity related comorbidities. Ann. Pediatr. Endocrinol. Metab. 2018, 23, 4–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early-OWO a | Late-OWO a | NW-A a | NW-B a | p-Value | |
|---|---|---|---|---|---|
| N | 388 | 254 | 186 | 118 | |
| Maternal Characteristics | |||||
| Age of delivery (years) | 28.7 ± 6.7 | 29.0 ± 6.8 | 27.4 ± 6.5 | 27.7 ± 6.3 | 0.038 |
| Race/ethnicity | 0.030 | ||||
| Black | 230 (59.3%) | 162 (63.8%) | 90 (48.4%) | 74 (62.7%) | |
| Hispanic | 95 (24.5%) | 49 (19.3%) | 49 (26.3%) | 30 (25.4%) | |
| White | 21 (5.4%) | 13 (5.1%) | 12 (6.5%) | 2 (1.7%) | |
| Others | 42 (10.8%) | 30 (11.8%) | 35 (18.8%) | 12 (10.2%) | |
| Education | 0.291 | ||||
| High school and above | 262 (67.5%) | 187 (73.6%) | 140 (75.3%) | 88 (74.6%) | |
| Less than high school | 123 (31.7%) | 65 (25.6%) | 46 (24.7%) | 30 (25.4%) | |
| Unknown | 3 (0.8%) | 2 (0.8%) | 0 (0.0%) | 0 (0.0%) | |
| Smoking during pregnancy | 0.112 | ||||
| Continuous | 52 (13.4%) | 21 (8.3%) | 14 (7.5%) | 9 (7.6%) | |
| Intermittent | 24 (6.2%) | 19 (7.5%) | 14 (7.5%) | 9 (7.6%) | |
| Never | 311 (80.2%) | 208 (81.9%) | 156 (83.9%) | 99 (83.9%) | |
| Unknown | 1 (0.3%) | 6 (2.4%) | 2 (1.1%) | 1 (0.8%) | |
| Maternal pregnancy overweight or obesity | <0.001 | ||||
| No | 151 (38.9%) | 109 (42.9%) | 109 (58.6%) | 61 (51.7%) | |
| Yes | 216 (55.7%) | 136 (53.5%) | 63 (33.9%) | 50 (42.4%) | |
| Unknown | 21 (5.4%) | 9 (3.5%) | 14 (7.5%) | 7 (5.9%) | |
| Cesarean section (N = 945) | 134 (34.6%) | 103 (40.6%) | 58 (31.2%) | 21 (17.8%) | <0.001 |
| Breastfeeding (N = 901) | 0.029 | ||||
| Both bottle-fed and breast-fed | 243 (65.5%) | 153 (62.7%) | 118 (69.4%) | 85 (73.3%) | |
| Bottle-fed | 107 (28.8%) | 68 (27.9%) | 33 (19.4%) | 21 (18.1%) | |
| Breast-fed | 21 (5.7%) | 23 (9.4%) | 19 (11.2%) | 10 (8.6%) | |
| Child’s Characteristics | |||||
| Sex: females | 173 (44.6%) | 121 (47.6%) | 76 (40.9%) | 54 (45.8%) | 0.563 |
| Birthweight (g) | 3229.6 ± 678.4 | 3068.2 ± 675.2 | 3009.8 ± 641.1 | 2837.6 ± 650.7 | <0.001 |
| Gestational age (weeks) | 38.6 ± 2.4 | 38.4 ± 2.6 | 38.7 ± 2.5 | 38.3 ± 2.7 | 0.340 |
| Preterm | 75 (19.3%) | 45 (17.7%) | 26 (14.0%) | 23 (19.5%) | 0.440 |
| Parity | 0.205 | ||||
| 0 | 163 (42.0%) | 113 (44.5%) | 90 (48.4%) | 38 (32.2%) | |
| 1 | 110 (28.4%) | 63 (24.8%) | 48 (25.8%) | 40 (33.9%) | |
| 2 | 59 (15.2%) | 44 (17.3%) | 32 (17.2%) | 23 (19.5%) | |
| 3+ | 56 (14.4%) | 34 (13.4%) | 16 (8.6%) | 17 (14.4%) | |
| Age at last visit (years) b | 9.2 (6.3–12.2) | 9.2 (7.1–11.1) | 7.8 (5.1–10.3) | 9.8 (7.2–12.7) | <0.001 |
| Height at last visit (cm) b | 138.9 (120.5–156.1) | 139.2 (122.7–150.9) | 126.2 (109.5–140.0) | 135.6 (122.4–155.9) | <0.001 |
| Weight at last visit (kg) b | 40.9 (27.3–60.8) | 40.7 (27.3–55.0) | 23.7 (17.1–31.5) | 30.2 (23.2–44.6) | <0.001 |
| BMI at last visit (kg/cm2) b | 21.3 (17.9–25.5) | 21.2 (17.8–24.5) | 15.4 (14.6–16.0) | 16.2 (15.4–18.2) | <0.001 |
| Overweight or obesity at last visit | 252 (64.9%) | 170 (66.9%) | 0 (0.0%) | 3 (2.5%) | <0.001 |
| Metabolite Module | Early-OWO vs. NW | Late-OWO vs. NW | ||||
|---|---|---|---|---|---|---|
| Odds Ratio (95% CI) | p-Value | FDR a, b | Odds Ratio (95% CI) | p-Value | FDR b | |
| red | 0.95 (0.91, 0.99) | 0.006 | 0.043 | 0.96 (0.92, 1.00) | 0.045 | 0.159 |
| brown | 0.96 (0.93, 0.99) | 0.014 | 0.049 | 0.96 (0.93, 1.00) | 0.038 | 0.159 |
| black | 0.97 (0.93, 1.01) | 0.163 | 0.352 | 0.96 (0.92, 1.00) | 0.069 | 0.161 |
| green | 1.02 (0.99, 1.06) | 0.201 | 0.352 | 1.01 (0.98, 1.05) | 0.481 | 0.728 |
| yellow | 1.02 (0.98, 1.06) | 0.275 | 0.386 | 1.00 (0.97, 1.05) | 0.808 | 0.808 |
| blue | 1.00 (0.98, 1.02) | 0.924 | 0.924 | 1.01 (0.98, 1.03) | 0.520 | 0.728 |
| turquoise | 1.00 (0.98, 1.02) | 0.829 | 0.924 | 1.01 (0.98, 1.03) | 0.685 | 0.799 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, T.; Zhao, J.; Hong, X.; Wang, G.; Hu, F.B.; Wang, X.; Liang, L. Cord Blood Metabolome and BMI Trajectory from Birth to Adolescence: A Prospective Birth Cohort Study on Early Life Biomarkers of Persistent Obesity. Metabolites 2021, 11, 739. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110739
Cao T, Zhao J, Hong X, Wang G, Hu FB, Wang X, Liang L. Cord Blood Metabolome and BMI Trajectory from Birth to Adolescence: A Prospective Birth Cohort Study on Early Life Biomarkers of Persistent Obesity. Metabolites. 2021; 11(11):739. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110739
Chicago/Turabian StyleCao, Tingyi, Jiaxuan Zhao, Xiumei Hong, Guoying Wang, Frank B. Hu, Xiaobin Wang, and Liming Liang. 2021. "Cord Blood Metabolome and BMI Trajectory from Birth to Adolescence: A Prospective Birth Cohort Study on Early Life Biomarkers of Persistent Obesity" Metabolites 11, no. 11: 739. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110739