
Opposing Immune-Metabolic Signature in Visceral Versus Subcutaneous Adipose Tissue in Patients with Adenocarcinoma of the Oesophagus and the Oesophagogastric Junction

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
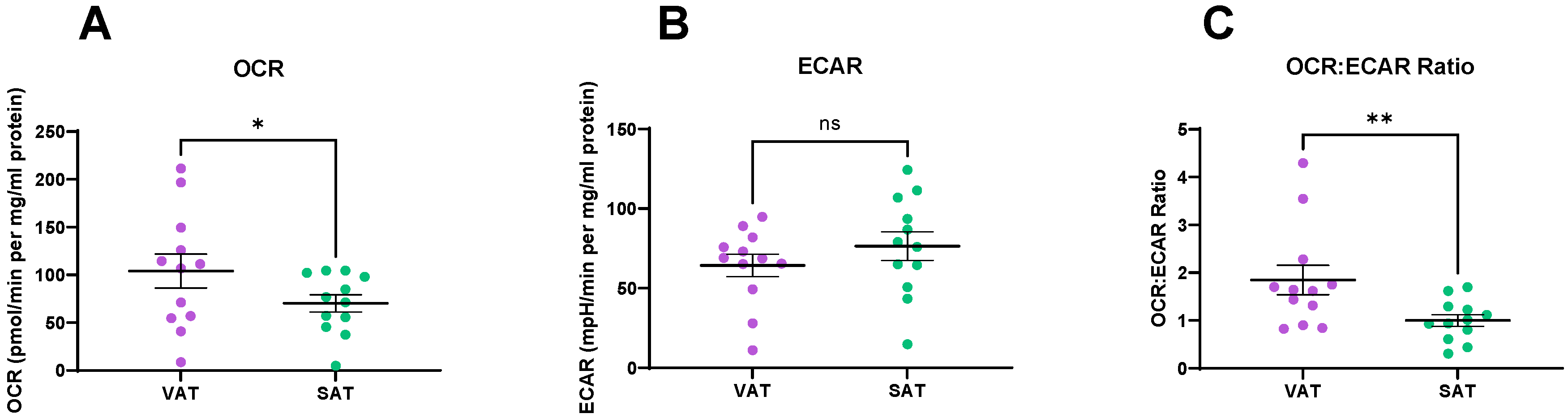
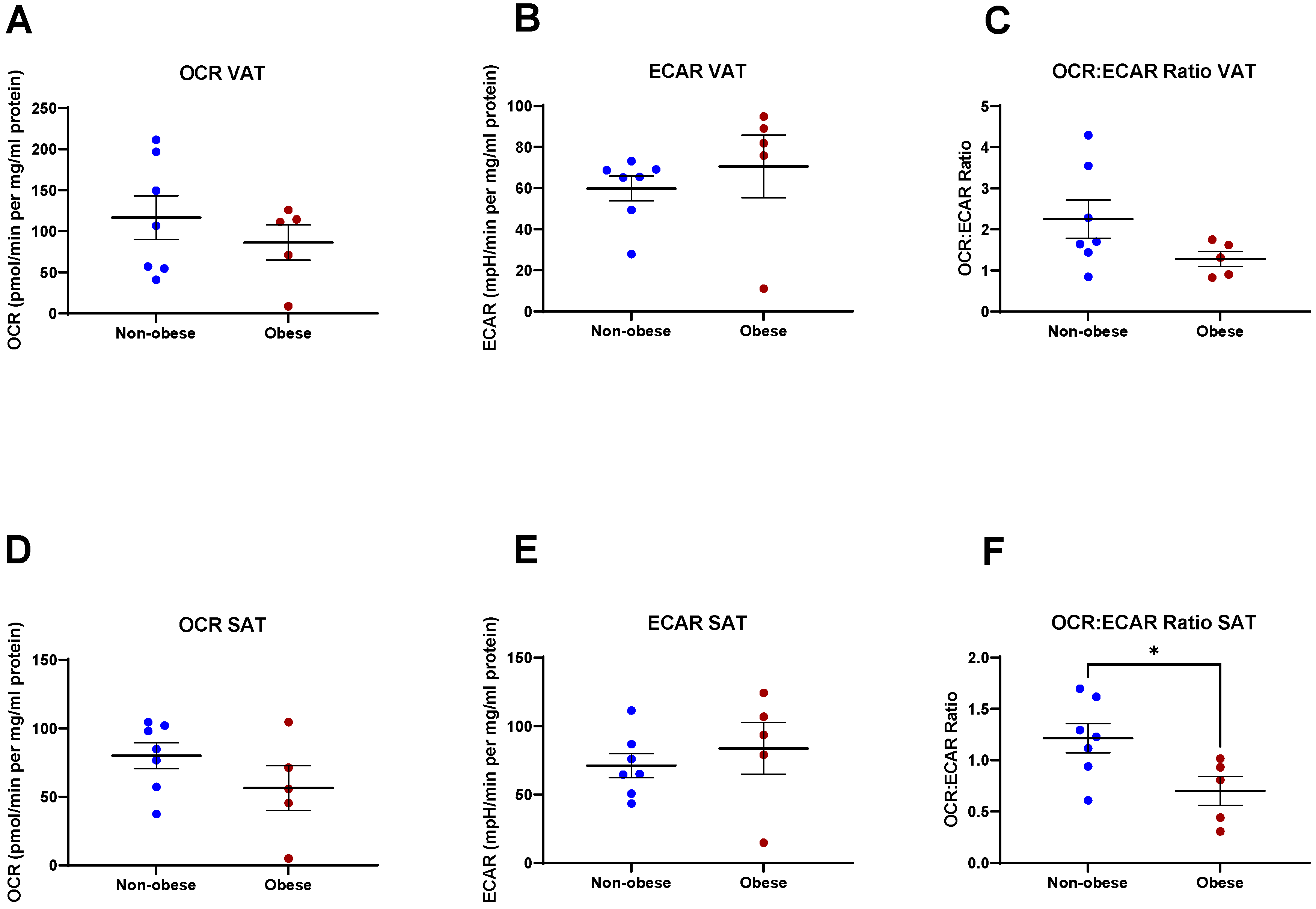
2.1. Metabolic Profiles Differ in VAT and SAT
2.2. Comparison of the Protein Secretome between VAT and SAT
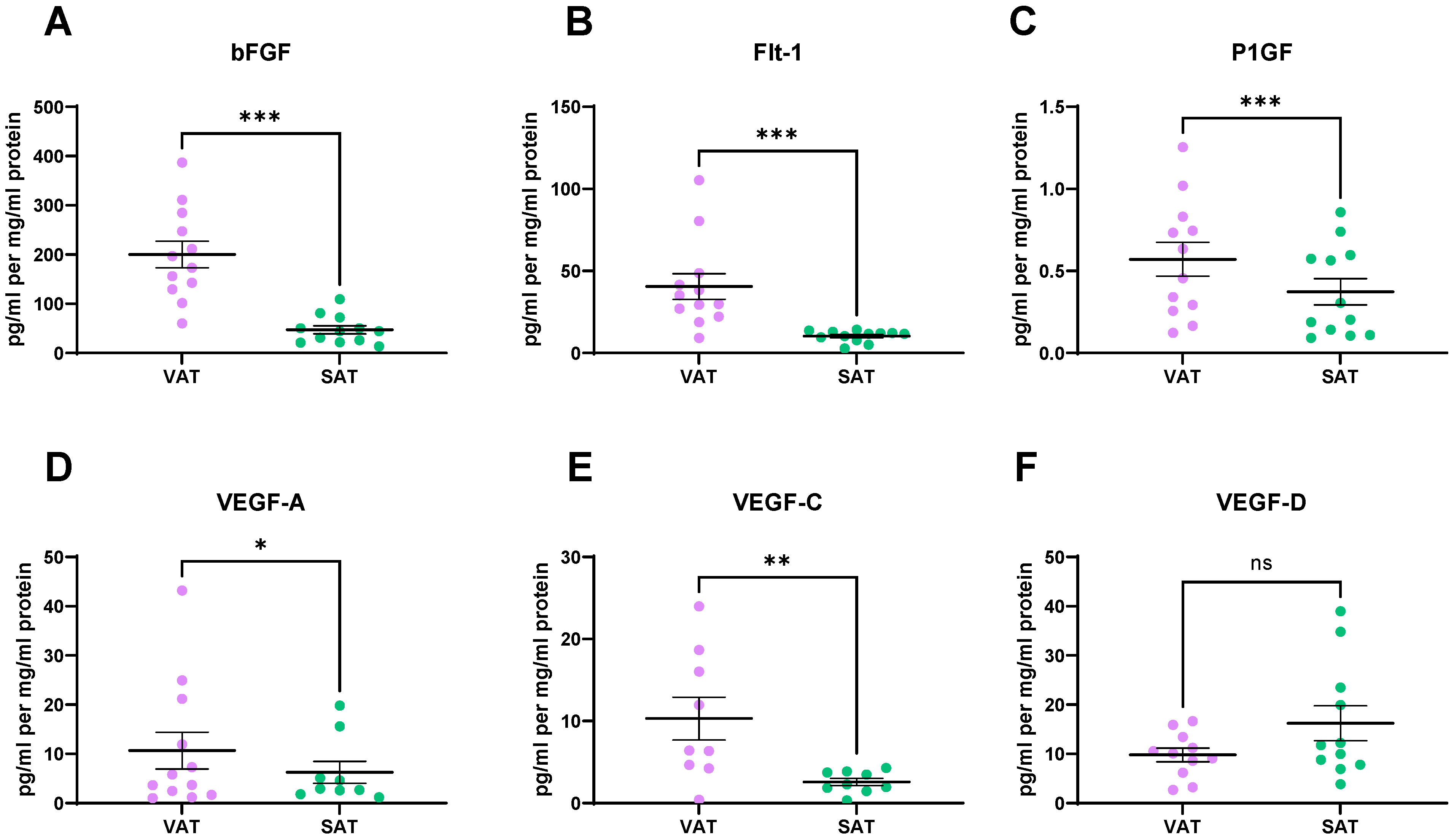
2.3. Angiogenic Factors Are Secreted at Significantly Higher Levels from VAT, When Compared to SAT
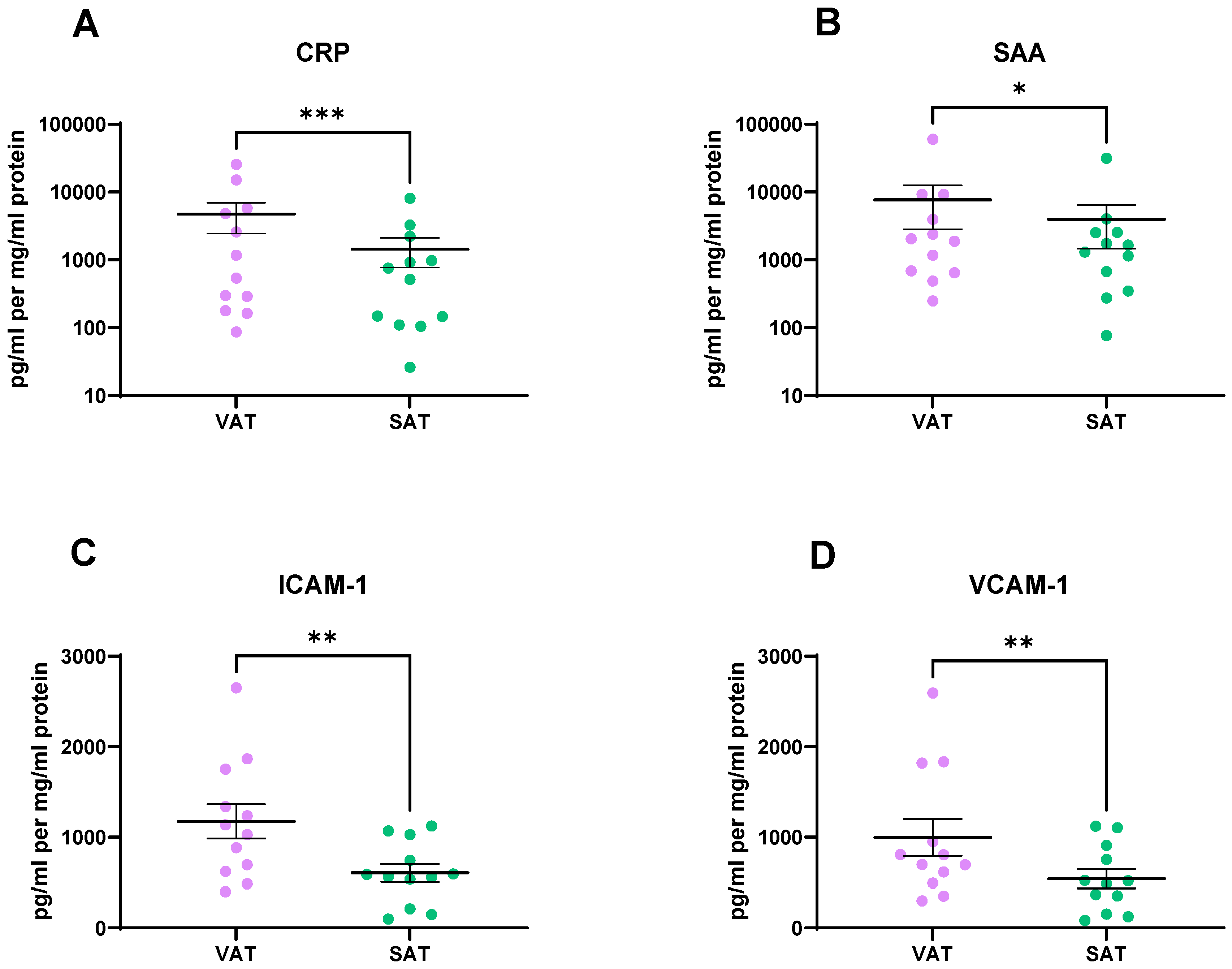
2.4. Vascular Injury Markers Were Found at Significantly Higher Levels in the VAT Secretome Compared to the SAT Secretome
2.5. VAT Secretes Higher Levels of Inflammatory Mediators, Cytokines, and Chemokines Compared to SAT
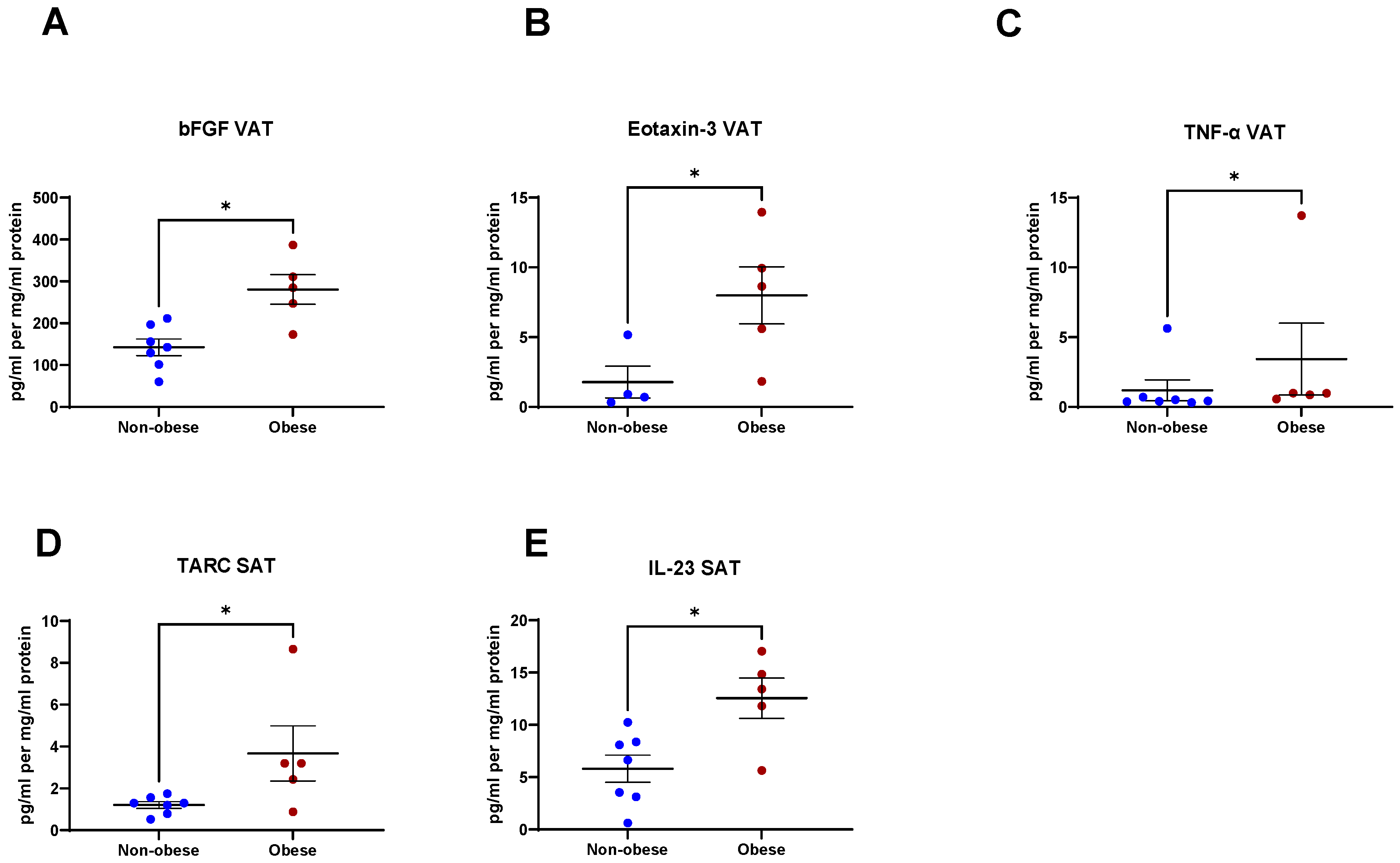
2.6. Secreted Factors from VAT and SAT Are Altered in Obese Patients
2.7. Linking Energy Metabolism to Secreted Factors in VAT and SAT to Derive an Immune-Metabolic Signature
2.8. Linking Anthropometric Parameters to Secreted Factors in VAT and SAT
3. Discussion
4. Materials and Methods
4.1. Patient Recruitment
4.2. OCR and ECAR Measurements in Visceral and Subcutaneous Adipose Tissue
4.3. Protein Isolation and Quantification from Adipose Tissue
4.4. Mesoscale Discovery (MSD) 54-Plex ELISA
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K.; International Agency for Research on Cancer Handbook Working Group. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; Ferri, L.E. Current Status of Management of Malignant Disease: Current Management of Esophageal Cancer. J. Gastrointest. Surg. 2015, 19, 964–972. [Google Scholar] [CrossRef]
- Van Hagen, P.; Hulshof, M.C.; Van Lanschot, J.; Steyerberg, E.W.; van Berge Henegouwen, M.I.; Wijnhoven, B.P.; Richel, D.J.; Nieuwenhuijzen, G.A.; Hospers, G.A.; Bonenkamp, J.J.; et al. Preoperative Chemoradiotherapy for Esophageal or Junctional Cancer. N. Engl. J. Med. 2012, 366, 2074–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum Murphy, M.; Xiao, L.; Patel, V.R.; Maru, D.M.; Correa, A.M.; Amlashi, F.G.; Liao, Z.; Komaki, R.; Lin, S.H.; Skinner, H.D.; et al. Pathological complete response in patients with esophageal cancer after the trimodality approach: The association with baseline variables and survival—The University of Texas MD Anderson Cancer Center experience. Cancer 2017, 123, 4106–4113. [Google Scholar] [CrossRef] [PubMed]
- Mongan, A.M.; Lynam-Lennon, N.; Doyle, S.L.; Casey, R.; Carr, E.; Cannon, A.; Conroy, M.J.; Pidgeon, G.P.; Brennan, L.; Lysaght, J.; et al. Visceral Adipose Tissue Modulates Radiosensitivity in Oesophageal Adenocarcinoma. Int. J. Med. Sci. 2019, 16, 519–528. [Google Scholar] [CrossRef] [Green Version]
- McQuade, J.L.; Daniel, C.R.; Hess, K.R.; Mak, C.; Wang, D.Y.; Rai, R.R.; Park, J.J.; Haydu, L.E.; Spencer, C.; Wongchenko, M.; et al. Association of body-mass index and outcomes in patients with metastatic melanoma treated with targeted therapy, immunotherapy, or chemotherapy: A retrospective, multicohort analysis. Lancet Oncol. 2018, 19, 310–322. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Aguilar, E.G.; Luna, J.I.; Dunai, C.; Khuat, L.T.; Le, C.T.; Mirsoian, A.; Minnar, C.M.; Stoffel, K.M.; Sturgill, I.R.; et al. Paradoxical effects of obesity on T cell function during tumor progression and PD-1 checkpoint blockade. Nat. Med. 2019, 25, 141–151. [Google Scholar] [CrossRef]
- Xu, H.; Cao, D.; He, A.; Ge, W. The prognostic role of obesity is independent of sex in cancer patients treated with immune checkpoint inhibitors: A pooled analysis of 4090 cancer patients. Int. Immunopharmacol. 2019, 74, 105745. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.M.; Dunne, M.R.; Lynam-Lennon, N.; Kennedy, S.A.; Cannon, A.; Reynolds, A.L.; Kennedy, B.N.; O’Sullivan, J. Pyrazinib (P3), [(E)-2-(2-Pyrazin-2-yl-vinyl)-phenol], a small molecule pyrazine compound enhances radiosensitivity in oesophageal adenocarcinoma. Cancer Lett. 2019, 447, 115–129. [Google Scholar] [CrossRef]
- Lynam-Lennon, N.; Connaughton, R.; Carr, E.; Mongan, A.-M.; O’Farrell, N.J.; Porter, R.K.; Brennan, L.; Pidgeon, G.P.; Lysaght, J.; Reynolds, J.V.; et al. Excess visceral adiposity induces alterations in mitochondrial function and energy metabolism in esophageal adenocarcinoma. BMC Cancer 2014, 14, 907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, M.; Coats, B.R.; Hisert, K.B.; Hagman, D.; Mutskov, V.; Peris, E.; Schoenfelt, K.Q.; Kuzma, J.N.; Larson, I.; Billing, P.S.; et al. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophages. Cell Metab. 2014, 20, 614–625. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.K.; Huang, S.C.-C.; Sergushichev, A.; Lampropoulou, V.; Ivanova, Y.; Loginicheva, E.; Chmielewski, K.; Stewart, K.M.; Ashall, J.; Everts, B.; et al. Network Integration of Parallel Metabolic and Transcriptional Data Reveals Metabolic Modules that Regulate Macrophage Polarization. Immunity 2015, 42, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Ralston, J.C.; Lyons, C.L.; Kennedy, E.B.; Kirwan, A.M.; Roche, H.M. Fatty Acids and NLRP3 Inflammasome–Mediated Inflammation in Metabolic Tissues. Annu. Rev. Nutr. 2017, 37, 77–102. [Google Scholar] [CrossRef]
- Kraunsøe, R.; Boushel, R.; Hansen, C.N.; Schjerling, P.; Qvortrup, K.; Støckel, M.; Mikines, K.J.; Dela, F. Mitochondrial respiration in subcutaneous and visceral adipose tissue from patients with morbid obesity. J. Physiol. 2010, 588, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Lysaght, J.; van der Stok, E.; Allott, E.; Casey, R.; Donohoe, C.; Howard, J.; McGarrigle, S.; Ravi, N.; Reynolds, J.V.; Pidgeon, G.P. Pro-inflammatory and tumour proliferative properties of excess visceral adipose tissue. Cancer Lett. 2011, 312, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Incio, J.; Tam, J.; Rahbari, N.N.; Suboj, P.; McManus, D.T.; Chin, S.M.; Vardam, T.D.; Batista, A.; Babykutty, S.; Jung, K.; et al. PlGF/VEGFR-1 Signaling Promotes Macrophage Polarization and Accelerated Tumor Progression in Obesity. Clin. Cancer Res. 2016, 22, 2993–3004. [Google Scholar] [CrossRef] [Green Version]
- Guiu, B.; Petit, J.M.; Bonnetain, F.; Ladoire, S.; Guiu, S.; Cercueil, J.P.; Krausé, D.; Hillon, P.; Borg, C.; Chauffert, B.; et al. Visceral fat area is an independent predictive biomarker of outcome after first-line bevacizumab-based treatment in metastatic colorectal cancer. Gut 2010, 59, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Kazazi-Hyseni, F.; Beijnen, J.H.; Schellens, J.H.M. Bevacizumab. Oncologist 2010, 15, 819–825. [Google Scholar] [CrossRef]
- Rupnick, M.A.; Panigrahy, D.; Zhang, C.-Y.; Dallabrida, S.M.; Lowell, B.B.; Langer, R.; Folkman, M.J. Adipose tissue mass can be regulated through the vasculature. Proc. Natl. Acad. Sci. USA 2002, 99, 10730–10735. [Google Scholar] [CrossRef] [Green Version]
- Ledoux, S.; Queguiner, I.; Msika, S.; Calderari, S.; Rufat, P.; Gasc, J.-M.; Pierre, C.; Larger, E. Angiogenesis Associated with Visceral and Subcutaneous Adipose Tissue in Severe Human Obesity. Diabetes 2008, 57, 3247–3257. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa-Hoshimoto, S.; Takahashi, K.; Bujo, H.; Hashimoto, N.; Yagui, K.; Saito, Y. Roles of degree of fat deposition and its localization on VEGF expression in adipocytes. Am. J. Physiol. Metab. 2005, 288, 1128–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gealekman, O.; Guseva, N.; Hartigan, C.; Apotheker, S.; Gorgoglione, M.; Gurav, K.; Tran, K.-V.; Straubhaar, J.; Nicoloro, S.; Czech, M.P.; et al. Depot-specific differences and insufficient subcutaneous adipose tissue angiogenesis in human obesity. Circulation 2011, 123, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cejkova, S.; Kubatova, H.; Thieme, F.; Janousek, L.; Fronek, J.; Poledne, R.; Lesna, I.K. The effect of cytokines produced by human adipose tissue on monocyte adhesion to the endothelium. Cell Adh. Migr. 2019, 13, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, I.; Pascot, A.; Prud’homme, D.; Alméras, N.; Bogaty, P.; Nadeau, A.; Bergeron, J.; Despres, J.-P. Elevated C-Reactive Protein:another component of the atherothrombotic profile of abdominal obesity. Arter. Thromb. Vasc. Biol. 2001, 21, 961–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forouhi, N.G.; Sattar, N.; McKeigue, P.M. Relation of C-reactive protein to body fat distribution and features of the metabolic syndrome in Europeans and South Asians. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1327–1331. [Google Scholar] [CrossRef] [Green Version]
- Poitou, C.; Coussieu, C.; Rouault, C.; Coupaye, M.; Cancello, R.; Bedel, J.-F.; Gouillon, M.; Bouillot, J.-L.; Oppert, J.-M.; Basdevant, A.; et al. Serum Amyloid A: A Marker of Adiposity-induced Low-grade Inflammation but Not of Metabolic Status. Obesity 2006, 14, 309–318. [Google Scholar] [CrossRef]
- Fain, J.N. Release of Inflammatory Mediators by Human Adipose Tissue Is Enhanced in Obesity and Primarily by the Nonfat Cells: A Review. Mediat. Inflamm. 2010, 2010, 513948. [Google Scholar] [CrossRef] [Green Version]
- Cancello, R.; Tordjman, J.; Poitou, C.; Guilhem, G.; Bouillot, J.L.; Hugol, D.; Coussieu, C.; Basdevant, A.; Bar Hen, A.; Bedossa, P.; et al. Increased Infiltration of Macrophages in Omental Adipose Tissue Is Associated with Marked Hepatic Lesions in Morbid Human Obesity. Diabetes 2006, 55, 1554–1561. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, R.W.; Metcalf, M.D.; White, A.E.; Madala, A.; Winters, B.R.; Maizlin, I.I.; Jobe, B.A.; Roberts, C.T., Jr.; Slifka, M.K.; Marks, D.L. Depot-specific differences in inflammatory mediators and a role for NK cells and IFN-γ in inflammation in human adipose tissue. Int. J. Obes. 2009, 33, 978–990. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Laurent, S.; Tang, Y.; Zong, H.; Vemulapalli, P.; Pessin, J.E. Adipocyte-Specific IKKβ Signaling Suppresses Adipose Tissue Inflammation through an IL-13-Dependent Paracrine Feedback Pathway. Cell Rep. 2014, 9, 1574–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darkhal, P.; Gao, M.; Ma, Y.; Liu, D. Blocking high-fat diet-induced obesity, insulin resistance and fatty liver by overexpression of Il-13 gene in mice. Int. J. Obes. 2015, 39, 1292–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Reyes, C.P.; Gómez-Arauz, A.Y.; Torres-Castro, I.; Manjarrez-Reyna, A.N.; Palomera, L.F.; Olivos-García, A.; Mendoza-Tenorio, E.; Sánchez-Medina, G.A.; Islas-Andrade, S.; Melendez-Mier, G.; et al. Serum Levels of Interleukin-13 Increase in Subjects with Insulin Resistance but Do Not Correlate with Markers of Low-Grade Systemic Inflammation. J. Diabetes Res. 2018, 2018, 7209872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacraz, G.; Rakotoarivelo, V.; Labbé, S.M.; Vernier, M.; Noll, C.; Mayhue, M.; Stankova, J.; Schwertani, A.; Grenier, G.; Carpentier, A.; et al. Deficiency of Interleukin-15 Confers Resistance to Obesity by Diminishing Inflammation and Enhancing the Thermogenic Function of Adipose Tissues. PLoS ONE 2016, 11, e0162995. [Google Scholar]
- Żelechowska, P.; Agier, J.; Kozłowska, E.; Brzezińska-Błaszczyk, E. Mast cells participate in chronic low-grade inflammation within adipose tissue. Obes. Rev. 2018, 19, 686–697. [Google Scholar] [CrossRef]
- Sumarac-Dumanovic, M.; Stevanovic, D.; Ljubic, A.; Jorga, J.; Simic, M.; Stamenkovic-Pejkovic, D.; Starcevic, V.; Trajkovic, V.; Micic, D. Increased activity of interleukin-23/interleukin-17 proinflammatory axis in obese women. Int. J. Obes. 2009, 33, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinan, E.M.; Doyle, S.L.; Bennett, A.E.; O’Neill, L.; Gannon, J.; Elliott, J.A.; O’Sullivan, J.; Reynolds, J.V.; Hussey, J. Sarcopenia during neoadjuvant therapy for oesophageal cancer: Characterising the impact on muscle strength and physical performance. Support. Care Cancer 2018, 26, 1569–1576. [Google Scholar] [CrossRef]
- Elliott, J.A.; Doyle, S.L.; Murphy, C.F.; King, S.; Guinan, E.M.; Beddy, P.; Ravi, N.; Reynolds, J.V. Sarcopenia: Prevalence, and Impact on Operative and Oncologic Outcomes in the Multimodal Management of Locally Advanced Esophageal Cancer. Ann. Surg. 2017, 266, 822–830. [Google Scholar] [CrossRef]
- Tan, B.H.; Brammer, K.; Randhawa, N.; Welch, N.T.; Parsons, S.L.; James, E.J.; Catton, J.A. Sarcopenia is associated with toxicity in patients undergoing neo-adjuvant chemotherapy for oesophago-gastric cancer. Eur. J. Surg. Oncol. 2015, 41, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Paireder, M.; Asari, R.; Kristo, I.; Rieder, E.; Tamandl, D.; Ba-Ssalamah, A.; Schoppmann, S. Impact of sarcopenia on outcome in patients with esophageal resection following neoadjuvant chemotherapy for esophageal cancer. Eur. J. Surg. Oncol. 2017, 43, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Fearon, K.C.H.; Glass, D.J.; Guttridge, D.C. Cancer Cachexia: Mediators, Signaling, and Metabolic Pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.-C.; Wang, C.-J.; Chao, Y.-J.; Chen, H.-Y.; Wang, H.-C.; Tung, H.-L.; Lin, J.-T.; Shan, Y.-S. Elevated Serum Interleukin-8 Level Correlates with Cancer-Related Cachexia and Sarcopenia: An Indicator for Pancreatic Cancer Outcomes. J. Clin. Med. 2018, 7, 502. [Google Scholar] [CrossRef] [Green Version]
- de Matos-Neto, E.M.; Lima, J.D.C.C.; de Pereira, W.O.; Figuerêdo, R.G.; Riccardi, D.M.D.R.; Radloff, K.; das Neves, R.X.; Camargo, R.G.; Maximiano, L.F.; Tokeshi, F.; et al. Systemic Inflammation in Cachexia-Is Tumor Cytokine Expression Profile the Culprit? Front. Immunol. 2015, 6, 629. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.L.; Bennett, A.M.; Donohoe, C.L.; Mongan, A.M.; Howard, J.M.; Lithander, F.E.; Pidgeon, G.P.; Reynolds, J.V.; Lysaght, J. Establishing computed tomography-defined visceral fat area thresholds for use in obesity-related cancer research. Nutr. Res. 2013, 33, 171–179. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adipose Depot | Metabolic Parameter | Factor | R Value | p Value | n |
|---|---|---|---|---|---|
| VAT | OCR | TNF-β | 0.7667 | 0.02 | 9 |
| VAT | ECAR | CRP | 0.7203 | 0.01 | 12 |
| VAT | ECAR | P1GF | 0.6294 | 0.03 | 12 |
| VAT | OCR:ECAR | TNF-β | 0.7167 | 0.03 | 9 |
| VAT | OCR:ECAR | Eotaxin | −0.8667 | 0.004 | 9 |
| SAT | OCR | IL-2 | −0.5874 | 0.04 | 12 |
| SAT | OCR | Eotaxin-3 | −0.7857 | 0.04 | 7 |
| SAT | ECAR | IFN-γ | −0.7818 | 0.01 | 10 |
| SAT | ECAR | IL-17D | −0.6294 | 0.03 | 12 |
| SAT | ECAR | ICAM-1 | 0.6154 | 0.03 | 12 |
| SAT | ECAR | IL-8 | −1 | 0.01 | 5 |
| SAT | ECAR | bFGF | 0.6294 | 0.03 | 12 |
| SAT | ECAR | VEGF-D | 0.7455 | 0.01 | 11 |
| SAT | OCR:ECAR | IP-10 | −0.5874 | 0.04 | 12 |
| SAT | OCR:ECAR | Tie-2 | −0.8857 | 0.03 | 6 |
| Anthropometric Parameter | Factor | R Value | p Value | n |
|---|---|---|---|---|
| SFA | Eotaxin | 0.8333 | 0.008 | 9 |
| Skeletal muscle | Eotaxin | −0.7167 | 0.03 | 9 |
| Skeletal muscle | MCP-1 | −0.5874 | 0.04 | 12 |
| Skeletal muscle | IL12p70 | −0.6014 | 0.04 | 12 |
| Skeletal muscle | IL-1β | −0.8571 | 0.02 | 7 |
| Skeletal muscle | IL-6 | −0.6084 | 0.03 | 12 |
| Skeletal muscle | IL-8 | −0.5874 | 0.04 | 12 |
| IMF | bFGF | 0.6182 | 0.04 | 11 |
| IMF | Eotaxin-3 | 0.7857 | 0.02 | 8 |
| IMF | IL-8 | 0.8810 | 0.007 | 8 |
| VFA | bFGF | 0.6503 | 0.02 | 12 |
| VFA | VEGF-C | 0.75 | 0.02 | 9 |
| Anthropometric Parameter | Factor | R Value | p Value | n |
|---|---|---|---|---|
| Skeletal muscle | VEGF-A | −0.7667 | 0.02 | 9 |
| IMF | TNF-α | 0.6727 | 0.02 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heeran, A.B.; McCready, J.; Dunne, M.R.; Donlon, N.E.; Nugent, T.S.; Bhardwaj, A.; Mitchelson, K.A.J.; Buckley, A.M.; Ravi, N.; Roche, H.M.; et al. Opposing Immune-Metabolic Signature in Visceral Versus Subcutaneous Adipose Tissue in Patients with Adenocarcinoma of the Oesophagus and the Oesophagogastric Junction. Metabolites 2021, 11, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110768
Heeran AB, McCready J, Dunne MR, Donlon NE, Nugent TS, Bhardwaj A, Mitchelson KAJ, Buckley AM, Ravi N, Roche HM, et al. Opposing Immune-Metabolic Signature in Visceral Versus Subcutaneous Adipose Tissue in Patients with Adenocarcinoma of the Oesophagus and the Oesophagogastric Junction. Metabolites. 2021; 11(11):768. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110768
Chicago/Turabian StyleHeeran, Aisling B., Jessica McCready, Margaret R. Dunne, Noel E. Donlon, Timothy S. Nugent, Anshul Bhardwaj, Kathleen A. J. Mitchelson, Amy M. Buckley, Narayanasamy Ravi, Helen M. Roche, and et al. 2021. "Opposing Immune-Metabolic Signature in Visceral Versus Subcutaneous Adipose Tissue in Patients with Adenocarcinoma of the Oesophagus and the Oesophagogastric Junction" Metabolites 11, no. 11: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110768