
Effect of Cadmium and Copper Exposure on Growth, Physio-Chemicals and Medicinal Properties of Cajanus cajan L. (Pigeon Pea)


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Plant Growth Attritubutes
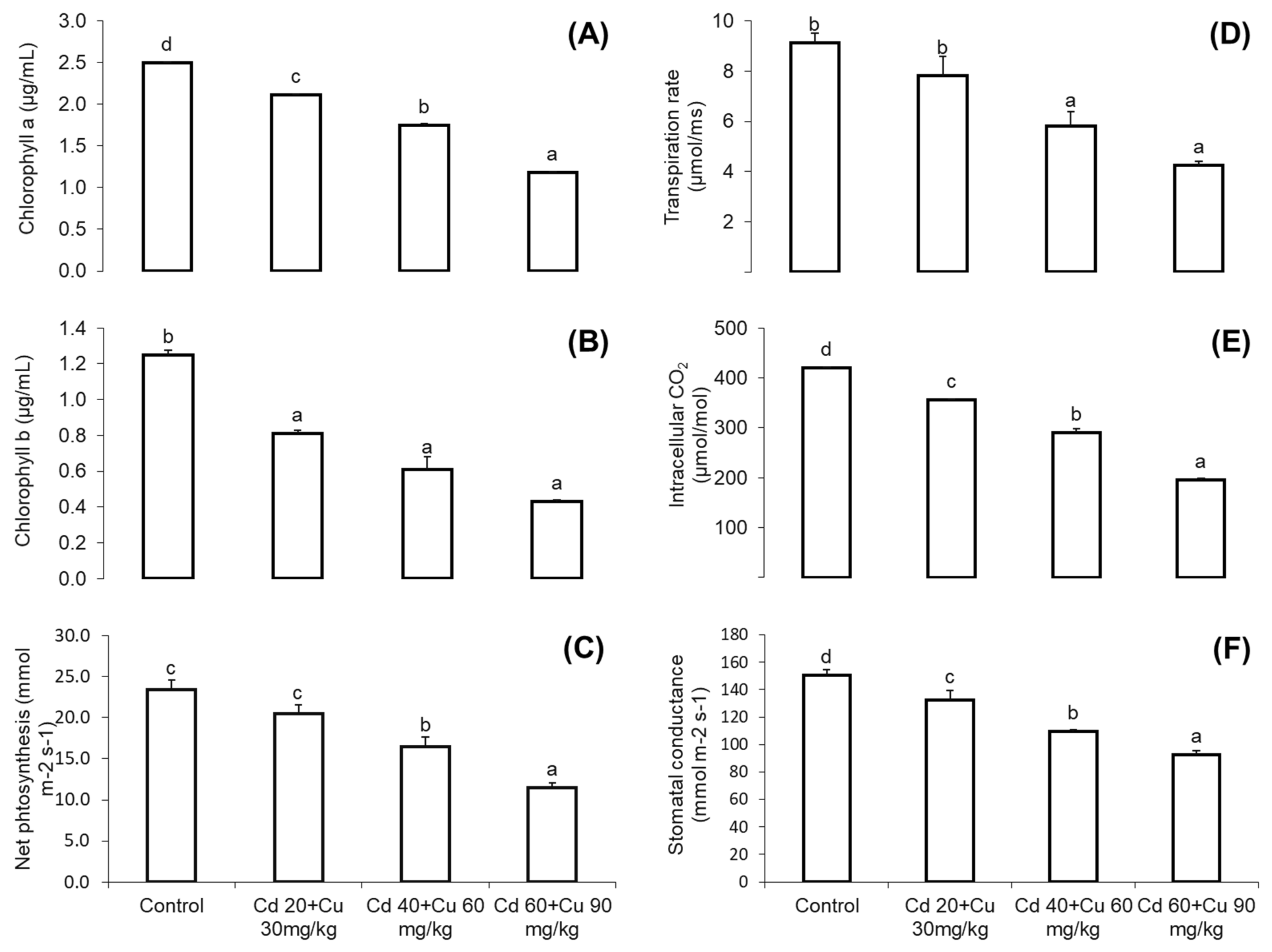
2.2. Chlorophyll Content and Photosynthetic Parameters
2.3. Flavonoid, Phenolic, and Proline Contents
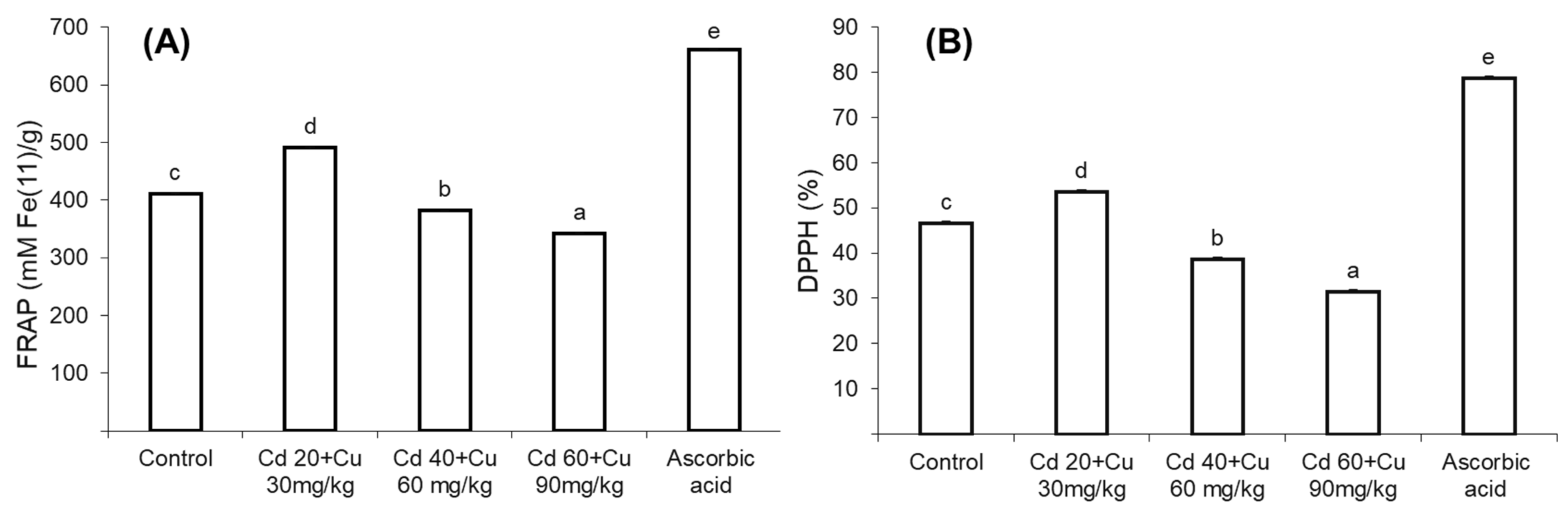
2.4. Medicinal Properties (Anti-Bacterial, DPPH, and FRAP Activity)
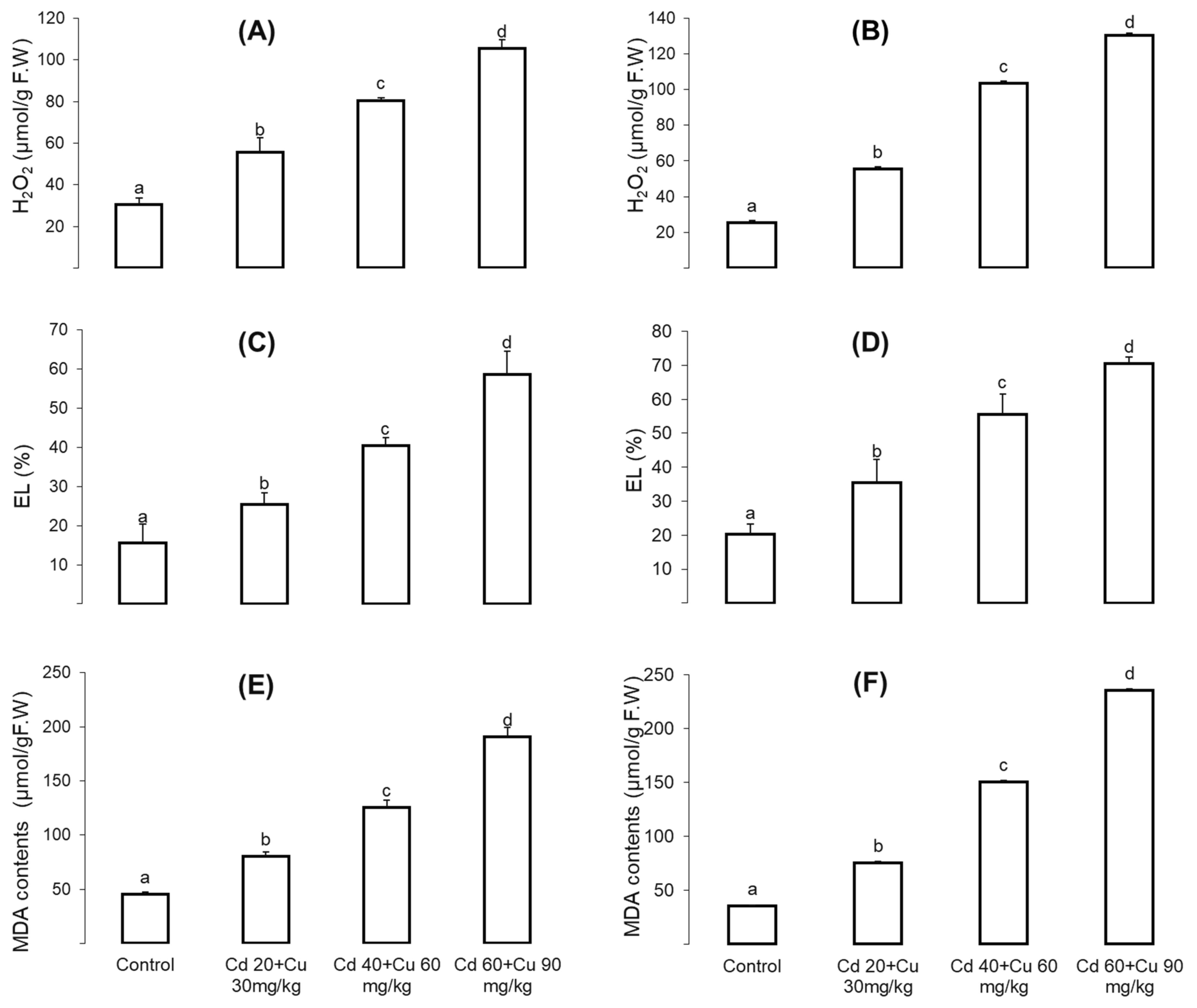
2.5. Oxidative Stresses
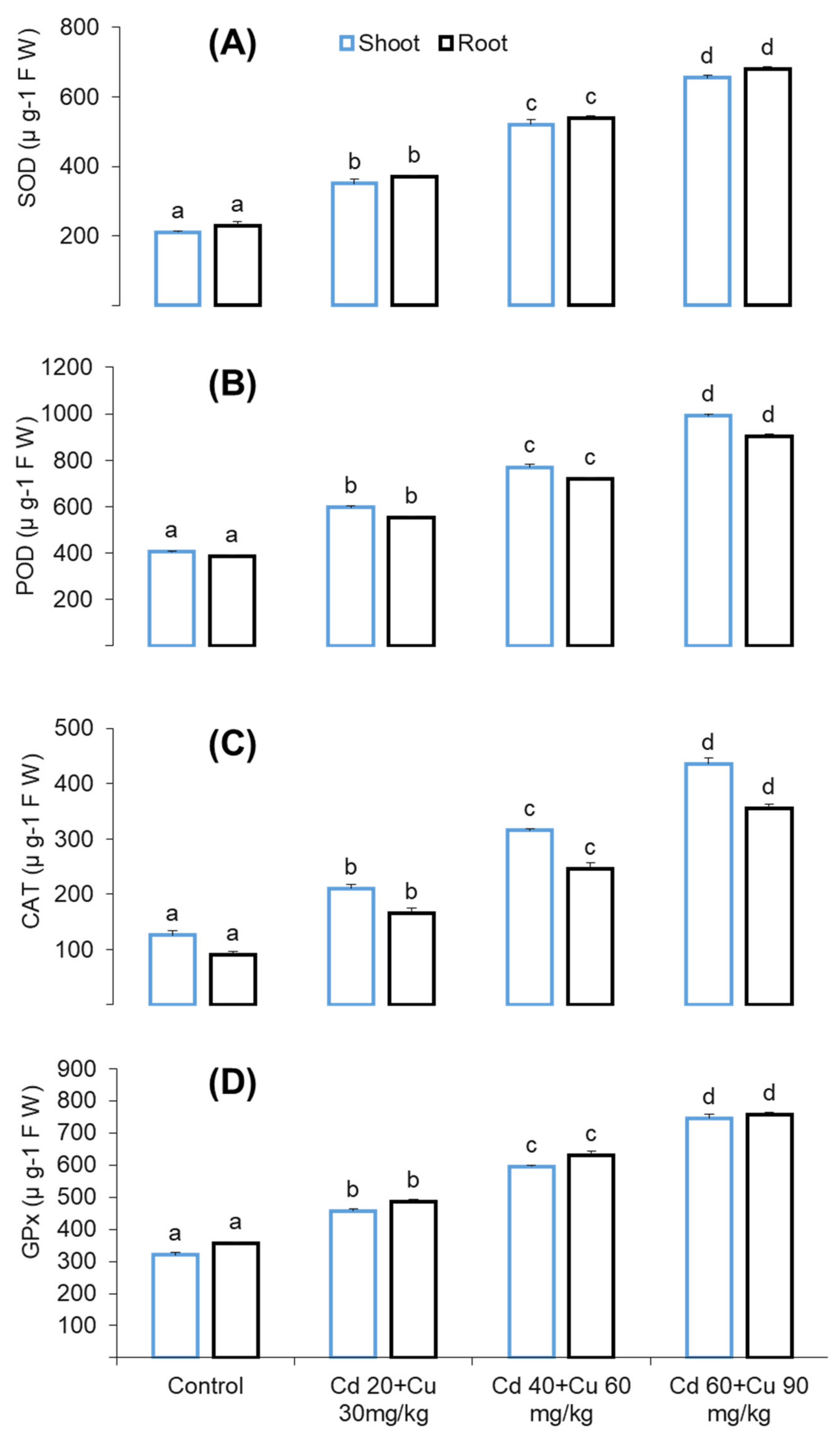
2.6. Antioxidant Enzyme Response
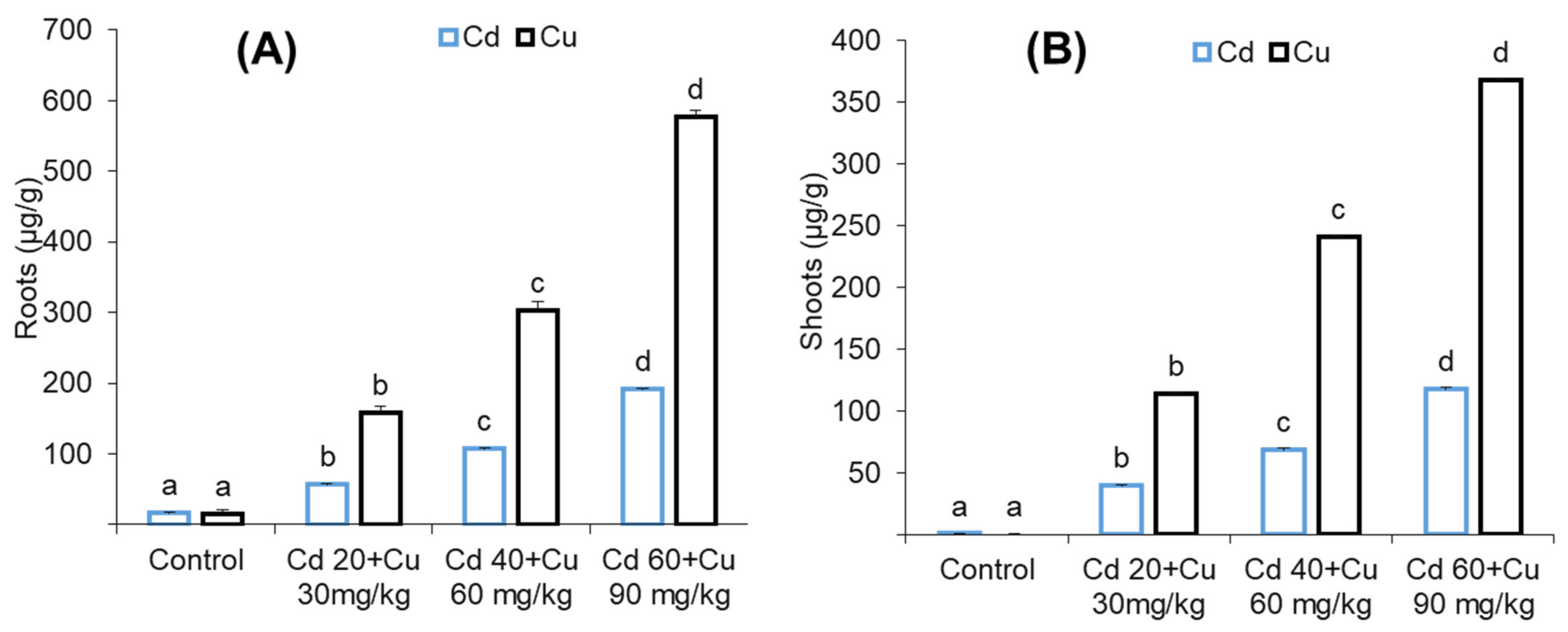
2.7. Metal (Cd and Cu) Accumulation
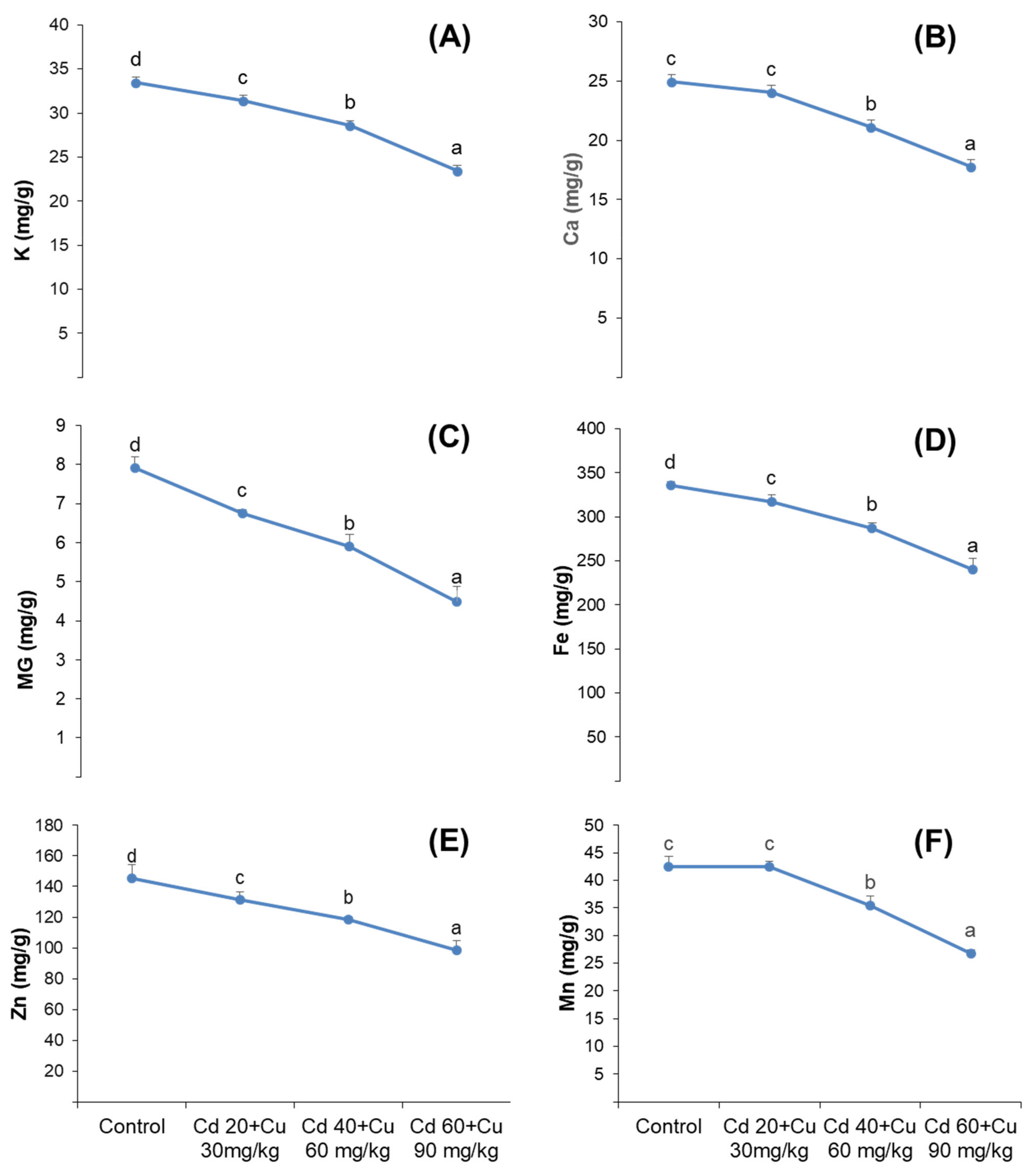
2.8. Nutrient Uptake
3. Discussion
4. Materials and Methods
4.1. Material and Growth Conditions
4.2. Soil Preparation and Experimental Designs
4.3. Determination of Gaseous Exchange
4.4. Chlorophyll a and b Determination
4.5. Determination of Proline Contents
4.6. Determination of Total Phenolics and Flavonoids
4.7. DPPH Free Radical Scavenging Activity
4.8. FRAP (Ferric-Reducing Antioxidant Power) Assay
4.9. Anti-Bacterial Assay
4.10. MDA Contents
4.11. Hydrogen Peroxide (H2O2)
4.12. Electrolyte Leakage
4.13. Antioxidant Enzyme Extraction
4.14. Enzyme Determinations
4.15. Elemental Analysis
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations and Acronyms
References
- Masindi, V.; Muedi, K.L. Environmental Contamination by Heavy Metals. Heavy Met. 2018, 10, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Zwolak, A.; Sarzyńska, M.; Szpyrka, E.; Stawarczyk, K. Sources of Soil Pollution by Heavy Metals and Their Accumulation in Vegetables: A Review. Water Air Soil Pollut. 2019, 230, 164. [Google Scholar] [CrossRef] [Green Version]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Jovanovic, M.; Boricic, N.; Wallace, D. Toxic effect of acute cadmium and lead exposure in rat blood, liver and kidney. Int. J. Environ. Res. Public Health 2019, 16, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuana, R.A.; Okieimen, F.E. Heavy metals in contaminated soils: A review of sources, chemistry, risks and best available strategies for remediation. Int. Sch. Res. Not. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Nriagu, J.O. Global Metal Pollution: Poisoning the Biosphere? Environ. Sci. Policy Sustain. Dev. 1990, 32, 7–33. [Google Scholar] [CrossRef]
- Nriagu, J.O.; Pacyna, J.M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 1988, 333, 134–139. [Google Scholar] [CrossRef]
- Violante, A.; Cozzolino, V.; Perelomov, L.; Caporale, A.; Pigna, M. Mobility and bioavailability of heavy metals and metalloids in soil environments. J. Soil Sci. Plant Nutr. 2010, 10, 268–292. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Shao, S.; Ni, H.; Fu, Z.; Hu, L.; Zhou, Y.; Min, X.; She, S.; Chen, S.; Huang, M. Current status, spatial features, health risks, and potential driving factors of soil heavy metal pollution in China at province level. Environ. Pollut. 2020, 266, 114961. [Google Scholar] [CrossRef]
- Yan, C.; Zhuang, T.; Bai, J.; Wen, X.; Lu, Q.; Zhang, L. Assessment of As, Cd, Zn, Cu and Pb Pollution and Toxicity in River Wetland Sediments and Artificial Wetland Soils Affected by Urbanization in a Chinese Delta. Wetlands 2020, 40, 2799–2809. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, L.; Dong, J.; Wu, J.; Ye, Z.; Zhao, W.; Ding, L.; Fu, W. Risk assessment, spatial patterns and source apportionment of soil heavy metals in a typical Chinese hickory plantation region of southeastern China. Geoderma 2020, 360, 114011. [Google Scholar] [CrossRef]
- Khan, A.; Ali, S.; Murad, W.; Hayat, K.; Siraj, S.; Jawad, M.; Khan, R.A.; Uddin, J.; Al-Harrasi, A.; Khan, A. Phytochemical and pharmacological uses of medicinal plants to treat cancer: A case study from Khyber Pakhtunkhwa, North Pakistan. J. Ethnopharmacol. 2021, 281, 114437. [Google Scholar] [CrossRef]
- Hayat, K.; Khan, J.; Khan, A.; Ullah, S.; Ali, S.; Fu, Y. Ameliorative Effects of Exogenous Proline on Photosynthetic Attributes, Nutrients Uptake, and Oxidative Stresses under Cadmium in Pigeon Pea (Cajanus cajan L.). Plants 2021, 10, 796. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Khan, J.; Hayat, K.; Abdelfattah Elateeq, A.; Salam, U.; Yu, B.; Tang, Z.H. Comparative Study of Growth, Cadmium Accumulation and Tolerance of Three Chickpea (Cicer arietinum L.) Cultivars. Plants 2020, 9, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aprile, A.; Sabella, E.; Francia, E.; Milc, J.; Ronga, D.; Pecchioni, N.; Ferrari, E.; Luvisi, A.; Vergine, M.; De Bellis, L. Combined Effect of Cadmium and Lead on Durum Wheat. Int. J. Mol. Sci. 2019, 20, 5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, M.C. Heavy Metal Concentrations in Soils and Factors Affecting Metal Uptake by Plants in the Vicinity of a Korean Cu-W Mine. Sensors 2008, 8, 2413–2423. [Google Scholar] [CrossRef] [Green Version]
- Vögeli-Lange, R.; Wagner, G.J. Relationship between cadmium, glutathione and cadmium-binding peptides (phytochelatins) in leaves of intact tobacco seedlings. Plant Sci. 1996, 114, 11–18. [Google Scholar] [CrossRef]
- Romanova, T.E.; Shuvaeva, O.V. Identification of the binding forms of cadmium during accumulation by water hyacinth. Chem. Speciat. Bioavailab. 2015, 27, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Llases, M.E.; Morgada, M.N.; Vila, A.J. Biochemistry of copper site assembly in heme-copper oxidases: A theme with variation. Int. J. Mol. Sci. 2019, 20, 3830. [Google Scholar] [CrossRef] [Green Version]
- Lidon, F.C.; Henriques, F.S. Limiting step on photosynthesis of rice plants treated with varying copper levels. J. Plant Physiol. 1991, 138, 115–118. [Google Scholar] [CrossRef]
- Shao, X.P.; Liu, W.B.; Xu, W.N.; Xia, W.; Jiang, Y.Y. Effects of dietary copper sources and levels on performance, copper status, plasma antioxidant activities and relative copper bioavailability in Carassius auratus gibelio. Aquaculture 2010, 308, 60–65. [Google Scholar] [CrossRef]
- Prasad, M.N.V.; Strzałka, K.; Hagemeyer, J. Impact of Heavy Metals on Photosynthesis. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 1999; pp. 117–138. [Google Scholar]
- Bechaieb, R.; Ben Akacha, A.; Gérard, H. Quantum chemistry insight into Mg-substitution in chlorophyll by toxic heavy metals: Cd, Hg and Pb. Chem. Phys. Lett. 2016, 663, 27–32. [Google Scholar] [CrossRef]
- Küpper, H.; Kiipper, F.; Spiller, M. Environmental relevance of heavy metal-substituted chlorophylls using the example of water plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Carrió-Seguí, À.; Romero, P.; Sanz, A.; Penarrubia, L. Interaction between ABA signaling and copper homeostasis in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 1568–1582. [Google Scholar]
- Ivakhnenko, E.P.; Knyazev, P.A.; Vitkovskaya, Y.G.; Popov, L.D.; Lyssenko, K.A.; Demidov, O.P.; Minkin, V.I. Synthesis, Staructure and Redox Properties of Cu (II) Chelate Complexes on the Basis of 2-(Hydroxyphenyl)-1H-benzo [d] imidazol-1-yl Phenol Ligands. Eur. J. Inorg. Chem. 2021, 2021, 2055–2062. [Google Scholar] [CrossRef]
- Pu, W.; Sun, J.; Zhang, F.; Wen, X.; Liu, W.; Huang, C. Effects of copper mining on heavy metal contamination in a rice agrosystem in the Xiaojiang River Basin, southwest China. Acta Geochim. 2019, 38, 753–773. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Kamran, M.; Iqbal, N.; Azeem, M.; Javed, M.T.; Ali, Q.; Haider, M.Z.; Irshad, S.; Rizwan, M.; et al. Ethylenediaminetetraacetic Acid (EDTA) Mitigates the Toxic Effect of Excessive Copper Concentrations on Growth, Gaseous Exchange and Chloroplast Ultrastructure of Corchorus capsularis L. and Improves Copper Accumulation Capabilities. Plants 2020, 9, 756. [Google Scholar] [CrossRef]
- Wu, L.I.N.; Thurman, D.A.; Bradshaw, A.D. The uptake of copper and its effect upon respiratory processes of roots of copper-tolerant and non-tolerant clones of Agrostis stolonifera. New Phytol. 1975, 75, 225–229. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Zhang, X.; Chen, C.-J.; Zhou, M.-G.; Wang, H.-C. Effects of fungicides JS399-19, azoxystrobin, tebuconazloe, and carbendazim on the physiological and biochemical indices and grain yield of winter wheat. Pestic. Biochem. Physiol. 2010, 98, 151–157. [Google Scholar] [CrossRef]
- Sghaier, D.B.; Duarte, B.; Bankaji, I.; Caçador, I.; Sleimi, N. Growth, chlorophyll fluorescence and mineral nutrition in the halophyte Tamarix gallica cultivated in combined stress conditions: Arsenic and NaCl. J. Photochem. Photobiol. B Biol. 2015, 149, 204–214. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-Induced Up-Regulation of the Antioxidant Defense and Methylglyoxal Detoxification System Reduces Salinity-Induced Damage in Rapeseed Seedlings. Biol. Trace Elem. Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Henriques, F.S. Biochemical, physiological, and structural effects of excess copper in plants. Bot. Rev. 1991, 57, 246–273. [Google Scholar] [CrossRef]
- Douchiche, O.; Rihouey, C.; Schaumann, A.; Driouich, A.; Morvan, C. Cadmium-induced alterations of the structural features of pectins in flax hypocotyl. Planta 2007, 225, 1301–1312. [Google Scholar] [CrossRef]
- Apori, O.S.; Hanyabui, E.; Asiamah, Y.J. Remediation Technology for Copper Contaminated Soil: A Review. Asian Soil Res. J. 2018, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.; Handley, H.; Taylor, M.P. Widespread copper and lead contamination of household drinking water, New South Wales, Australia. Environ. Res. 2016, 151, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.H.; Kong, Y.C.; Zain, N.A.M. Effect of Cadmium and Copper Exposure on Growth, Secondary Metabolites and Antioxidant Activity in the Medicinal Plant Sambung Nyawa (Gynura procumbens (Lour.) Merr). Molecules 2017, 22, 1623. [Google Scholar] [CrossRef] [Green Version]
- Hayat, K.; Ali, S.; Ullah, S.; Fu, Y.; Hussain, M. Green synthesized silver and copper nanoparticles induced changes in biomass parameters, secondary metabolites production, and antioxidant activity in callus cultures of Artemisia absinthium L. Green Process. Synth. 2021, 10, 61–72. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Arasimowicz-Jelonek, M.; Izbiańska-Jankowska, K.; Frontasyeva, M.; Zinicovscaia, I.; Guiance-Varela, C.; Deckert, J. NADPH oxidase is involved in regulation of gene expression and ROS overproduction in soybean (Glycine max L.) seedlings exposed to cadmium. Acta Soc. Bot. Pol. 2017, 86, 1–17. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Springer: Singapore, 2017. [Google Scholar]
- Sarangthem, J.; Jain, M.; Gadre, R. Inhibition of δ-aminolevulinic acid dehydratase activity by cadmium in excised etiolated maize leaf segments during greening. Plant Soil Environ. 2011, 57, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Wojcik, M.; Vangronsveld, J.; Tukiendorf, A. Cadmium tolerance in Thlaspi caerulescensI. Growth parameters, metal accumulation and phytochelatin synthesis in response to cadmium. Environ. Exp. Bot. 2005, 53, 151–161. [Google Scholar] [CrossRef]
- Printz, B.; Lutts, S.; Hausman, J.F.; Sergeant, K. Copper trafficking in plants and its implication on cell wall dynamics. Front. Plant Sci. 2016, 7, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutase: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Mwamba, T.M.; Ali, S.; Ali, B.; Lwalaba, J.L.; Liu, H.; Farooq, M.A.; Shou, J.; Zhou, W. Interactive effects of cadmium and copper on metal accumulation, oxidative stress, and mineral composition in Brassica napus. Int. J. Environ. Sci. Technol. 2016, 13, 2163–2174. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Gautam, A.; Dubey, R.S. Metal toxicity in plants: Induction of oxidative stress, antioxidative defense system, metabolic alterations and phytoremediation. In Molecular Physiology of Abiotic Stresses in Plant Productivity; Hemantaranjan, A., Ed.; Scientific Publisher: Jodhpur, India, 2018; pp. 256–290. [Google Scholar]
- Alscher, R.G.; Erturk, N.; Heath, L. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Pinto, D.C.G.A. Biological Potential and Medical Use of Secondary Metabolites. Medicines 2019, 6, 66. [Google Scholar] [CrossRef] [Green Version]
- Nasim, S.A.; Dhir, B. Heavy metals alter the potency of medicinal plants. Rev. Environ. Contam. Toxicol. 2009, 203, 139–149. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Sumner, M.E. Methods of Soil Analysis, Part 3. Chemical Methods; SSSA Inc./ASA Inc.: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Okigbo, R.N.; Omodamiro, O.D. Antimicrobial effect of leaf extracts of pigeon pea (Cajanus cajan (L.) Millsp.) on some human pathogens. J. Herbs Spices. Med Plants 2007, 12, 117–127. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Mensor, L.L.; Menezes, F.S.; Leitão, G.G.; Reis, A.S.; Santos, T.C.D.; Coube, C.S.; Leitão, S.G. Screening of Brazilian plant extracts for antioxidant activity by the use of DPPH free radical method. Phytother. Res. 2001, 15, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Nwodo, U.U.; Ngene, A.A.; Iroegbu, C.U.; Onyedikachi, O.A.L.; Chigor, V.; Okoh, A. In vivo evaluation of the antiviral activity of Cajanus cajan on measles virus. Arch. Virol. 2011, 15, 1551–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. N.Y. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized Assay for Hydrogen Peroxide Determination in Plant Tissue Using Potassium Iodide. Am. J. Anal. Chem. 2014, 5, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.M.; Matloub, A.A.; Aboutabl, M.E.; Ibrahim, N.A.; Mohamed, S.M. Assessment of anti-inflammatory, antinociceptive, immunomodulatory, and antioxidant activities of Cajanus cajan L. seeds cultivated in Egypt and its phytochemical composition. Pharm. Biol. 2015, 54, 1380–1391. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Fu, K.; Fu, Y.J.; Zu, Y.G.; Chang, F.R.; Chen, Y.H.; Gu, C.B. Antioxidant activities of extracts and main components of pigeon pea [Cajanus cajan (L.) Millsp.] leaves. Molecules 2009, 14, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Roots | Shoots | ||||
|---|---|---|---|---|---|---|
| Fresh Weight (g) | Dry Weight (g) | Length (cm) | Fresh Weight (g) | Dry Weight (g) | Length (cm) | |
| Control | 5.55 ± 1.0 d | 2.61 ± 0.23 d | 23.40 ± 1.1 d | 9.45 ± 0.55 b | 5.11 ± 0.43 d | 91.40 ± 1.15 d |
| Cd 20 + Cu 30 mg/kg | 5.10 ± 0.99 c | 2.42 ± 0.60 c | 21.40 ± 0.85 c | 8.75 ± 0.77 b | 4.91 ± 0.89 c | 86.10 ± 0.85 c |
| Cd 40 + Cu 60 mg/kg | 4.72 ± 0.7 b | 2.11 ± 0.32 b | 18.45 ± 0.77 b | 7.11 ± 0.01 a | 4.01 ± 1.02 b | 80.50 ± 0.5 b |
| Cd 60 + Cu 90 mg/kg | 3.92 ± 1.23 a | 1.82 ± 0.94 a | 13.45 ± 0.9 a | 6.21 ± 0.5 a | 3.21 ± 0.93 a | 73.40 ± 0.23 a |
| Treatment | Flavonoids (mg/g DM GAE) | Phenols (mg/g DM Rutin) | Proline (µg/g FW) |
|---|---|---|---|
| Control | 32.35 ± 0.34 c | 55.51 ± 0.91 c | 20.43 ± 0.15 a |
| Cd 20 + Cu 30 mg/kg | 42.40 ± 0.12 d | 68.53 ± 0.21 d | 45.41 ± 0.18 b |
| Cd 40 + Cu 60 mg/kg | 26.45 ± 0.39 b | 45.42 ± 0.45 b | 70.40 ± 0.71 c |
| Cd 60 + Cu 90 mg/kg | 17.41 ± 0.78 a | 32.46 ± 0.81 a | 90.47 ± 0.12 d |
| Antibacterial Activities | Inhibition Zone (mm) | ||
|---|---|---|---|
| Gram-Positive Bacteria | Gram-Negative Bacteria | ||
| S. aureus | E. coli | S. thyphi | |
| Control | 11.60 ± 0.86 c | 10.52 ± 0.57 c | 12.36 ± 0.97 c |
| Cd 20 + Cu 30mg/kg | 12.93 ± 0.34 d | 11.22 ± 0.77 d | 13.75 ± 0.58 d |
| Cd 40 + Cu 60 mg/kg | 10.10 ± 0.57 b | 9.92 ± 0.33 b | 10.91 ± 0.11 b |
| Cd 60 + Cu 90mg/kg | 9.65 ± 0.83 a | 8.44 ± 0.45 a | 8.21 ± 0.63 a |
| Ciprofloxacin | 17.61 ± 0.88 e | 19.59 ± 0.71 e | 21.29 ± 0.77 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayat, K.; Khan, A.; Bibi, F.; Salahuddin; Murad, W.; Fu, Y.; Batiha, G.E.-S.; Alqarni, M.; Khan, A.; Al-Harrasi, A. Effect of Cadmium and Copper Exposure on Growth, Physio-Chemicals and Medicinal Properties of Cajanus cajan L. (Pigeon Pea). Metabolites 2021, 11, 769. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110769
Hayat K, Khan A, Bibi F, Salahuddin, Murad W, Fu Y, Batiha GE-S, Alqarni M, Khan A, Al-Harrasi A. Effect of Cadmium and Copper Exposure on Growth, Physio-Chemicals and Medicinal Properties of Cajanus cajan L. (Pigeon Pea). Metabolites. 2021; 11(11):769. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110769
Chicago/Turabian StyleHayat, Khizar, Asif Khan, Farkhanda Bibi, Salahuddin, Waheed Murad, Yujie Fu, Gaber El-Saber Batiha, Mohammed Alqarni, Ajmal Khan, and Ahmed Al-Harrasi. 2021. "Effect of Cadmium and Copper Exposure on Growth, Physio-Chemicals and Medicinal Properties of Cajanus cajan L. (Pigeon Pea)" Metabolites 11, no. 11: 769. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110769