
The Effects of Consuming White Button Mushroom Agaricus bisporus on the Brain and Liver Metabolome Using a Targeted Metabolomic Analysis

Abstract
:1. Introduction
2. Results
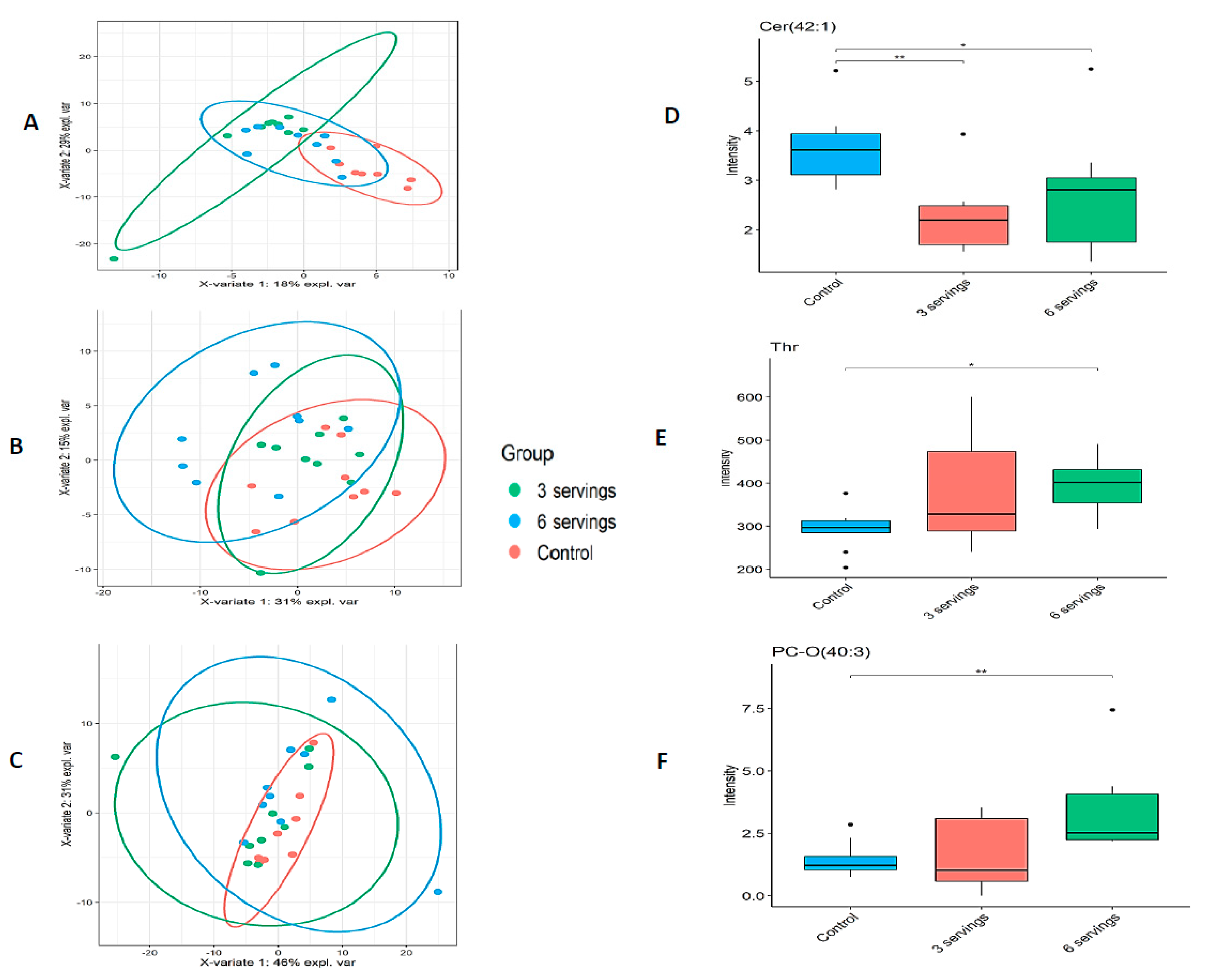
2.1. Metabolome Profile Analysis
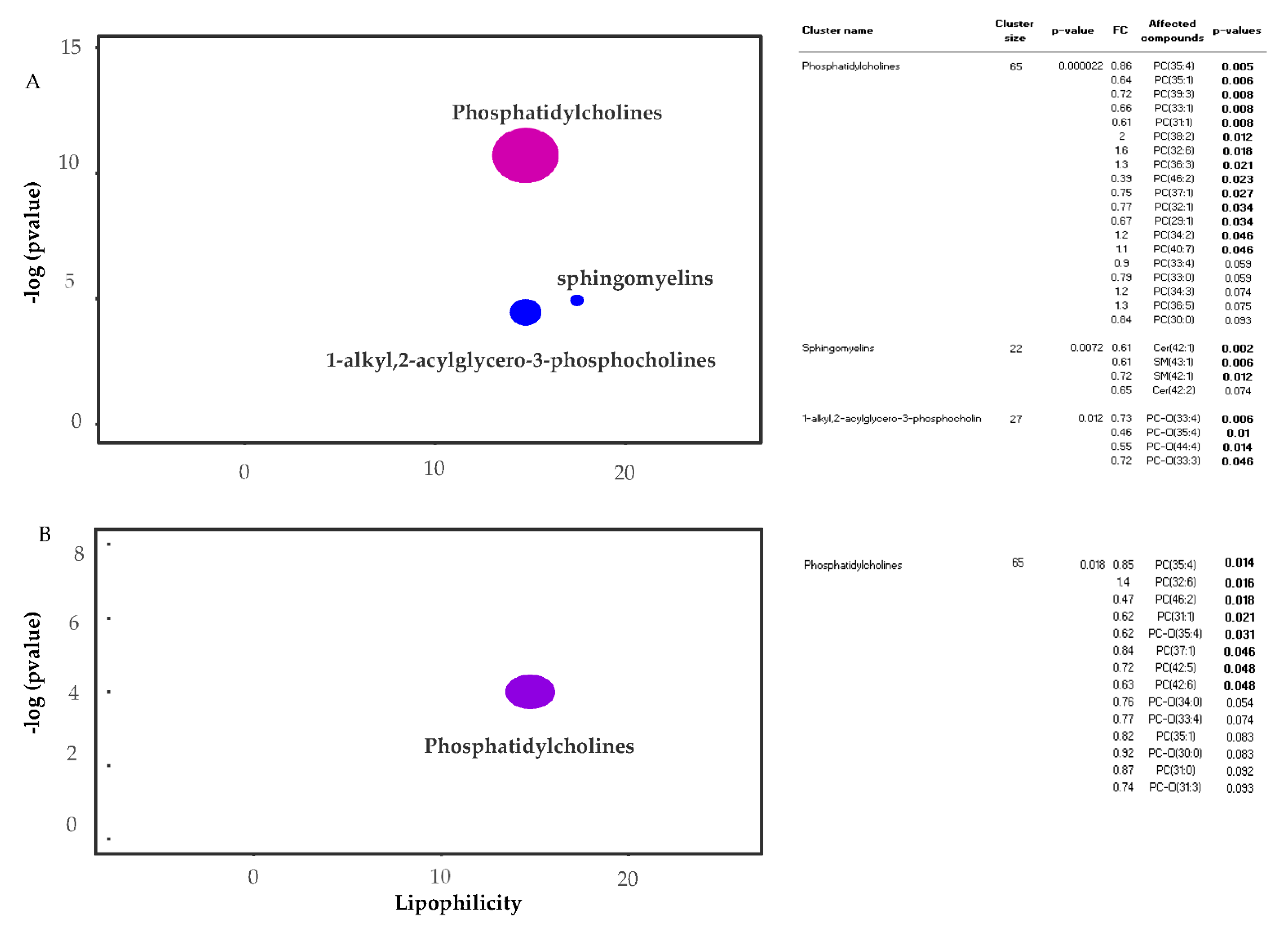
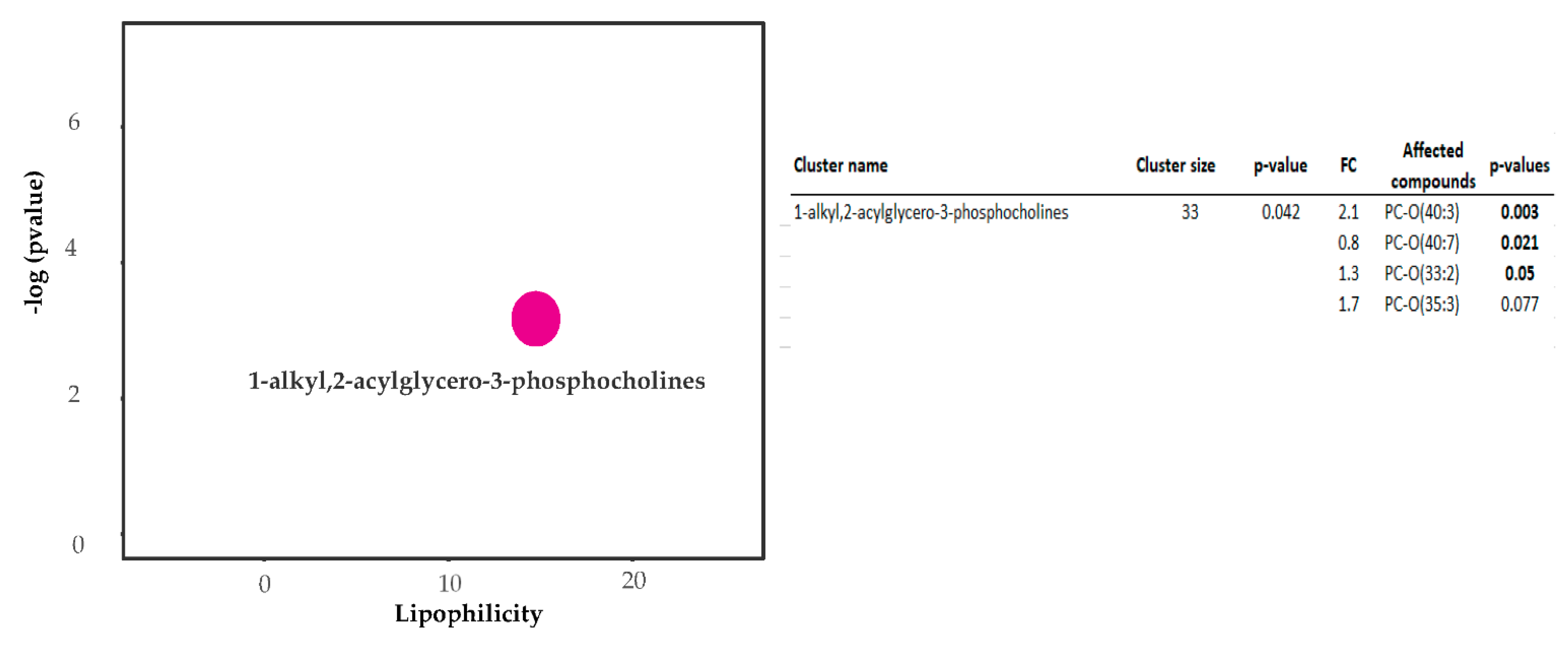
2.2. Metabolic Analysis by ChemRICH
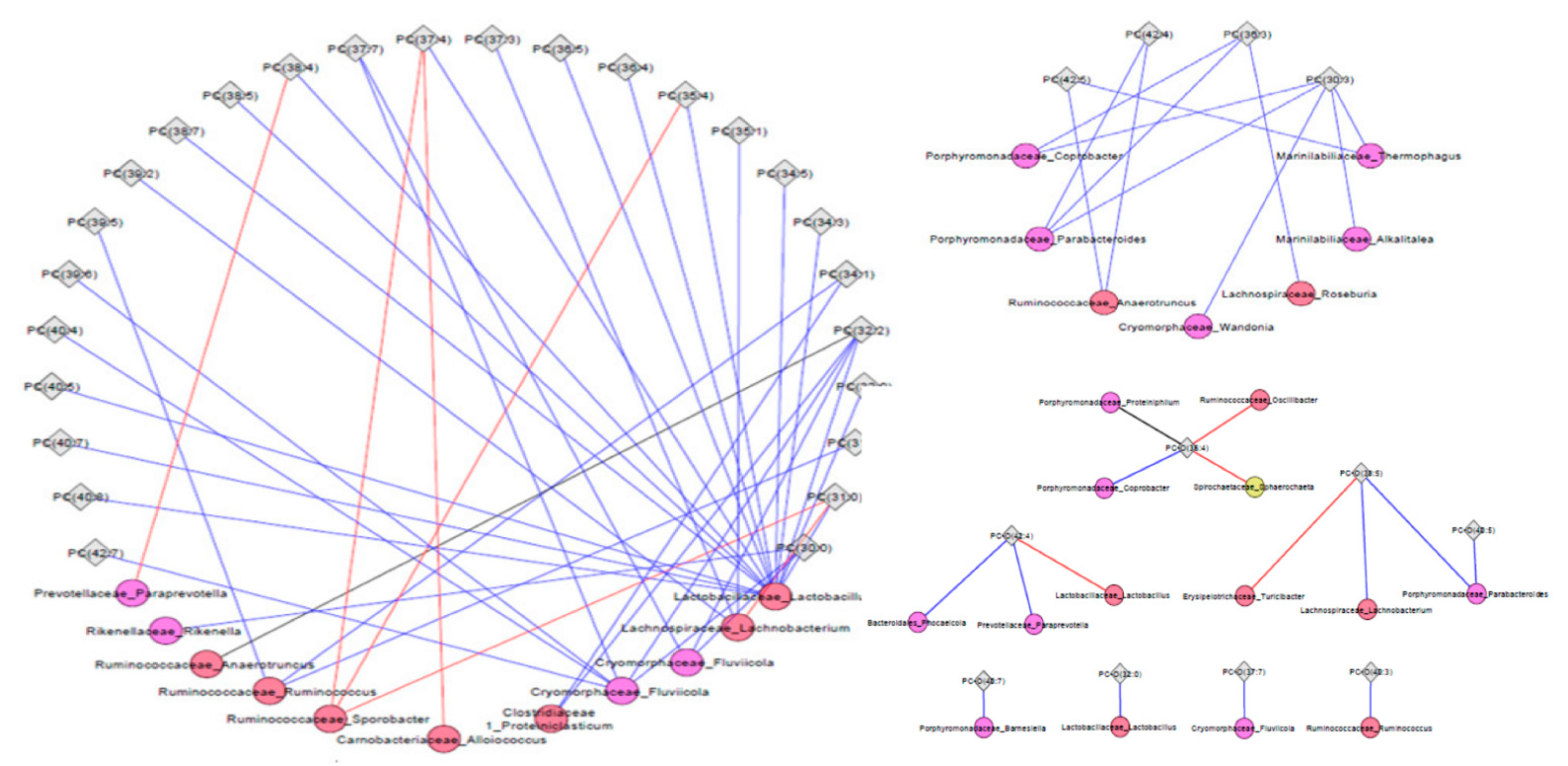
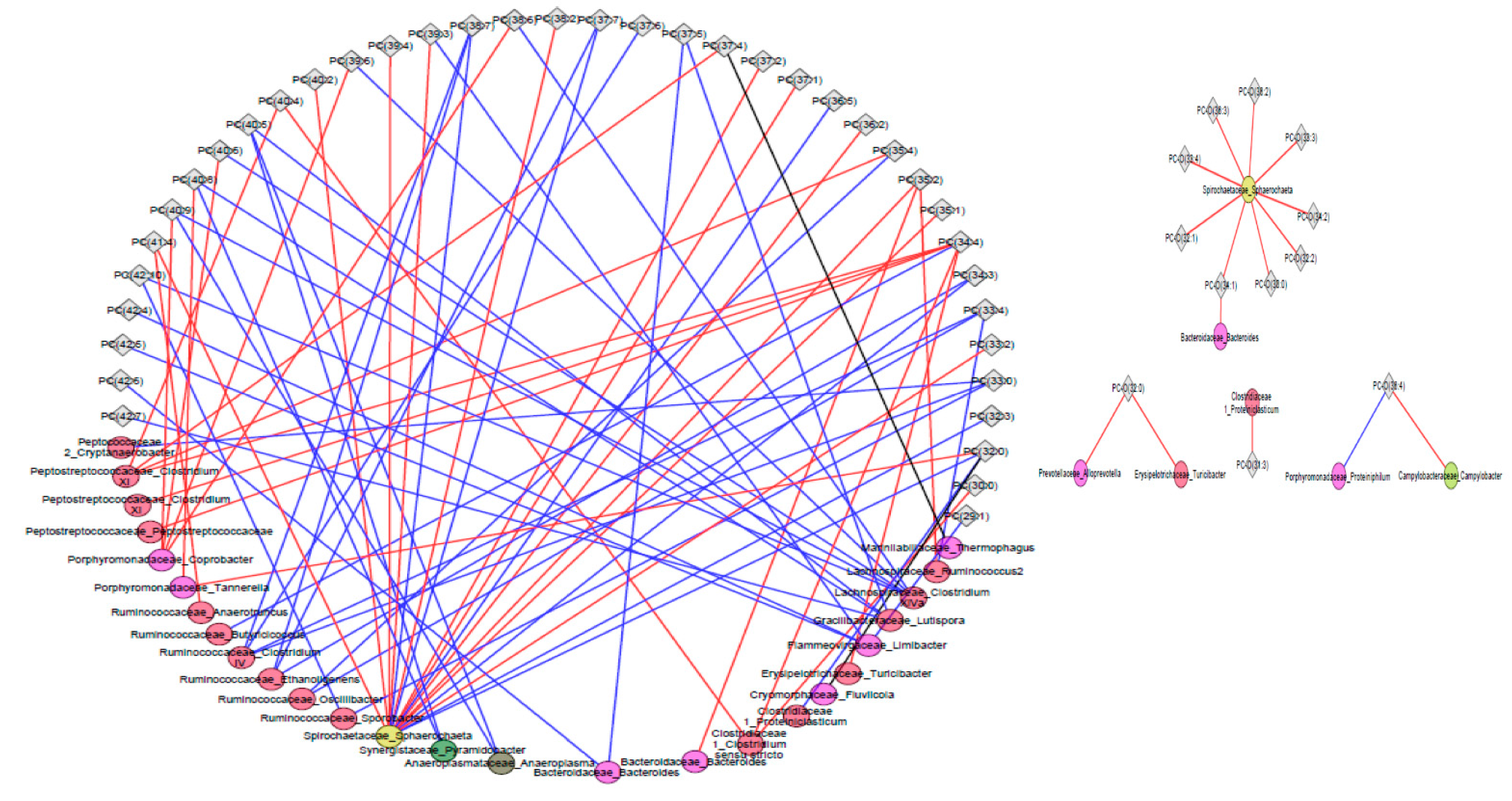
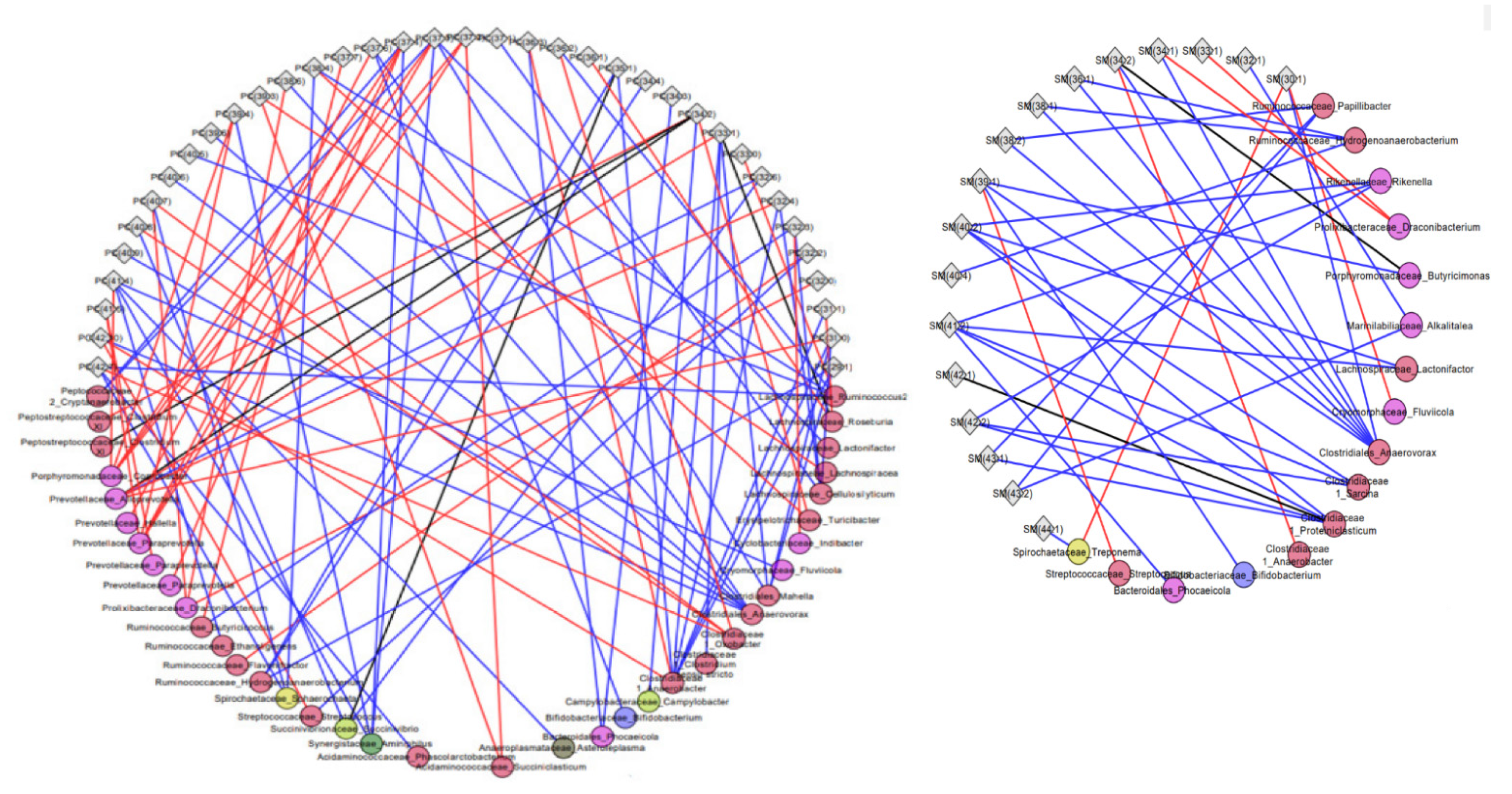
2.3. Associations of Metabolites with Fecal Microbiota
2.4. RNA Sequencing of Brain
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Diet Formulation and Tissue Sample Collection
4.3. Targeted Metabolomic Analysis
4.4. Statistical Analysis of Metabolomic Data
4.5. RNA Sequencing and Mapping
4.6. RNA-Seq Data Analysis
4.7. Differentially Expressed Genes-Real Time PCR Validation
4.8. 16 S rDNA Amplicon Multi-Tag Sequencing Data and Metabolite Correlations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fogarasi, M.; Socaci, S.A.; Dulf, F.V.; Diaconeasa, Z.M.; Fărcaș, A.C.; Tofană, M.; Semeniuc, C.A. Bioactive Compounds and Volatile Profiles of Five Transylvanian Wild Edible Mushrooms. Molecules 2018, 23, 3272. [Google Scholar] [CrossRef] [Green Version]
- Niego, A.; Rapior, S.; Thongklang, N.; Raspé, O.; Jaidee, W.; Lumyong, S.; Hyde, K. Macrofungi as a Nutraceutical Source: Promising Bioactive Compounds and Market Value. J. Fungi 2021, 7, 397. [Google Scholar] [CrossRef]
- Sande, D.; de Oliveira, G.P.; e Moura, M.A.F.; Martins, B.D.A.; Lima, M.T.N.S.; Takahashi, J.A. Edible mushrooms as a ubiquitous source of essential fatty acids. Food Res. Int. 2019, 125, 108524. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine—A diet-derived antioxidant with therapeutic potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumfield, M.; Abbott, K.; Duve, E.; Cassettari, T.; Marshall, S.; Fayet-Moore, F. Examining the health effects and bioactive components in Agaricus bisporus mushrooms: A scoping review. J. Nutr. Biochem. 2020, 84, 108453. [Google Scholar] [CrossRef]
- Ma, G.; Du, H.; Hu, Q.; Yang, W.; Pei, F.; Xiao, H. Health benefits of edible mushroom polysaccharides and associated gut microbiota regulation. Crit. Rev. Food Sci. Nutr. 2021, 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zheng, C.; Yang, J.; Li, J.; Su, J.; Xie, Y.; Lai, G. Immunomodulatory Activities of a Fungal Protein Extracted from Hericium erinaceus through Regulating the Gut Microbiota. Front. Immunol. 2017, 8, 666. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, M.; Zhang, C.; Tian, C.; Wang, X.; Song, X.; Jing, H.; Gao, Z.; Ren, Z.; Liu, W.; et al. Purification, in vitro antioxidant and in vivo anti-aging activities of soluble polysaccharides by enzyme-assisted extraction from Agaricus bisporus. Int. J. Biol. Macromol. 2018, 109, 457–466. [Google Scholar] [CrossRef]
- Venkatesh Gobi, V.; Rajasankar, S.; Ramkumar, M.; Dhanalakshmi, C.; Manivasagam, T.; Justin Thenmozhi, A.; Mohamed Essa, M.; Chidambaram, R.; Kalandar, A. Agaricus blazei extract abrogates rotenone-induced dopamine depletion and motor deficits by its anti-oxidative and anti-inflammatory properties in Parkinsonic mice. Nutr. Neurosci. 2018, 21, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Asad, F.; Anwar, H.; Yassine, H.M.; Ullah, M.I.; Rahman, A.-U.; Kamran, Z.; Sohail, M.U. White Button Mushroom, Agaricus bisporus (Agaricomycetes), and a Probiotics Mixture Supplementation Correct Dyslipidemia without Influencing the Colon Microbiome Profile in Hypercholesterolemic Rats. Int. J. Med. Mushrooms 2020, 22, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.C.; Jeong, Y.T.; Yang, B.K.; Islam, R.; Koyyalamudi, S.R.; Pang, G.; Cho, K.Y.; Song, C.H. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetic and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef]
- Su, A.; Yang, W.; Zhao, L.; Pei, F.; Yuan, B.; Zhong, L.; Ma, G.; Hu, Q. Flammulina velutipes polysaccharides improve scopolamine-induced learning and memory impairment in mice by modulating gut microbiota composition. Food Funct. 2018, 9, 1424–1432. [Google Scholar] [CrossRef]
- Thangthaeng, N.; Miller, M.G.; Gomes, S.M.; Shukitt-Hale, B. Daily supplementation with mushroom (Agaricus bisporus) improves balance and working memory in aged rats. Nutr. Res. 2015, 35, 1079–1084. [Google Scholar] [CrossRef]
- Rai, S.N.; Mishra, D.; Singh, P.; Vamanu, E.; Singh, M. Therapeutic applications of mushrooms and their biomolecules along with a glimpse of in silico approach in neurodegenerative diseases. Biomed. Pharmacother. 2021, 137, 111377. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-E.; Wang, H.-L.; Lu, K.-H.; Huang, Y.-J.; Panyod, S.; Liu, W.-T.; Yang, S.-H.; Chen, M.-H.; Lu, Y.-S.; Sheen, L.-Y. Water extract of Armillaria mellea (Vahl) P. Kumm. Alleviates the depression-like behaviors in acute- and chronic mild stress-induced rodent models via anti-inflammatory action. J. Ethnopharmacol. 2021, 265, 113395. [Google Scholar] [CrossRef]
- Yadav, D.; Negi, P.S. Bioactive components of mushrooms: Processing effects and health benefits. Food Res. Int. 2021, 148, 110599. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, N.; Nakao, S.; Nishiyama, M.; Takeda, Y.; Ishimoto, T.; Masuo, Y.; Matsumoto, S.; Suzuki, M.; Kato, Y. Oral Administration of the Food-Derived Hydrophilic Antioxidant Ergothioneine Enhances Object Recognition Memory in Mice. Curr. Mol. Pharmacol. 2021, 14, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Dominguez, R.; Castellano-Escuder, P.; Carmona, F.; Lefevre-Arbogast, S.; Low, D.Y.; Preez, A.D.; Ruigrok, S.R.; Manach, C.; Urpi-Sarda, M.; Korosi, A.; et al. Food and Microbiota Metabolites Associate with Cognitive Decline in Older Subjects: A twelve-Year Prospective Study. Mol Nutr Food Res. 2021, e2100606. [Google Scholar] [CrossRef] [PubMed]
- Cermenati, G.; Mitro, N.; Audano, M.; Melcangi, R.C.; Crestani, M.; De Fabiani, E.; Caruso, D. Lipids in the nervous system: From biochemistry and molecular biology to patho-physiology. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Morais, L.H.; Iv, H.L.S.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Genet. 2021, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.V.; Sherwin, E.; Schellekens, H.; Stanton, C.; Dinan, T.; Cryan, J.F. Feeding the microbiota-gut-brain axis: Diet, microbiome, and neuropsychiatry. Transl. Res. 2017, 179, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.; Hsu, T.M.; Kanoski, S.E. Gut to Brain Dysbiosis: Mechanisms Linking Western Diet Consumption, the Microbiome, and Cognitive Impairment. Front. Behav. Neurosci. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berding, K.; Vlckova, K.; Marx, W.; Schellekens, H.; Stanton, C.; Clarke, G.; Jacka, F.; Dinan, T.G.; Cryan, J.F. Diet and the Microbiota–Gut–Brain Axis: Sowing the Seeds of Good Mental Health. Adv. Nutr. 2021, 12, 1239–1285. [Google Scholar] [CrossRef] [PubMed]
- Giles, C.; Takechi, R.; Mellett, N.A.; Meikle, P.; Dhaliwal, S.; Mamo, J.C. The Effects of Long-Term Saturated Fat Enriched Diets on the Brain Lipidome. PLoS ONE 2016, 11, e0166964. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Liu, C.-W.; Yang, Y.; Hsiao, Y.-C.; Ru, H.; Lu, K. High-coverage metabolomics uncovers microbiota-driven biochemical landscape of interorgan transport and gut-brain communication in mice. Nat. Commun. 2021, 12, 6000. [Google Scholar] [CrossRef] [PubMed]
- Solano-Aguilar, G.I.; Jang, S.; Lakshman, S.; Gupta, R.; Beshah, E.; Sikaroodi, M.; Vinyard, B.; Molokin, A.; Gillevet, P.M.; Urban, J.J.F. The Effect of Dietary Mushroom Agaricus bisporus on Intestinal Microbiota Composition and Host Immunological Function. Nutrient 2018, 10, 1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warr, A.; Affara, N.; Aken, B.; Beiki, H.; Bickhart, D.M.; Billis, K.; Chow, W.; Eory, L.; A Finlayson, H.; Flicek, P.; et al. An improved pig reference genome sequence to enable pig genetics and genomics research. GigaScience 2020, 9, giaa051. [Google Scholar] [CrossRef]
- Dawson, H.D.; Chen, C.; Gaynor, B.; Shao, J.; Urbanjr, J.F. The porcine translational research database: A manually curated, genomics and proteomics-based research resource. BMC Genom. 2017, 18, 643. [Google Scholar] [CrossRef] [Green Version]
- Weiss, H.; Wester-Rosenloef, L.; Koch, C.; Koch, F.; Baltrusch, S.; Tiedge, M.; Ibrahim, S. The Mitochondrial Atp8 Mutation Induces Mitochondrial ROS Generation, Secretory Dysfunction, and β-Cell Mass Adaptation in Conplastic B6-mtFVB Mice. Endocrinology 2012, 153, 4666–4676. [Google Scholar] [CrossRef] [PubMed]
- Dalangin, R.; Kim, A.; Campbell, R.E. The Role of Amino Acids in Neurotransmission and Fluorescent Tools for Their Detection. Int. J. Mol. Sci. 2020, 21, 6197. [Google Scholar] [CrossRef] [PubMed]
- Rabiei, Z.; Bigdeli, M.R.; Rasoulian, B. Neuroprotection of dietary virgin olive oil on brain lipidomics during stroke. Curr. Neurovasc. Res. 2013, 10, 231–237. [Google Scholar] [CrossRef]
- Carrié, I.; Clément, M.; De Javel, D.; Francès, H.; Bourre, J.M. Specific phospholipid fatty acid composition of brain regions in mice. Effects of n-3 polyunsaturated fatty acid deficiency and phospholipid supplementation. J. Lipid Res. 2000, 41, 465–472. [Google Scholar] [CrossRef]
- Naoe, S.; Tsugawa, H.; Takahashi, M.; Ikeda, K.; Arita, M. Characterization of Lipid Profiles after Dietary Intake of Polyunsaturated Fatty Acids Using Integrated Untargeted and Targeted Lipidomics. Metababolites 2019, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulou, M.; Gordillo, R.; Koliaki, C.; Gancheva, S.; Jelenik, T.; De Filippo, E.; Herder, C.; Markgraf, D.; Jankowiak, F.; Esposito, I.; et al. Specific Hepatic Sphingolipids Relate to Insulin Resistance, Oxidative Stress, and Inflammation in Nonalcoholic Steatohepatitis. Diabetes Care 2018, 41, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonteh, A.N.; Chiang, A.J.; Arakaki, X.; Edminster, S.P.; Harrington, M.G. Accumulation of Cerebrospinal Fluid Glycerophospholipids and Sphingolipids in Cognitively Healthy Participants with Alzheimer’s Biomarkers Precedes Lipolysis in the Dementia Stage. Front. Neurosci. 2020, 14, 1320. [Google Scholar] [CrossRef] [PubMed]
- Blasko, I.; Defrancesco, M.; Oberacher, H.; Loacker, L.; Kemmler, G.; Marksteiner, J.; Humpel, C. Plasma phosphatidylcholines and vitamin B12/folate levels are possible prognostic biomarkers for progression of Alzheimer’s disease. Exp. Gerontol. 2021, 147, 111264. [Google Scholar] [CrossRef]
- Naudi, A.; Cabre, R.; Jove, M.; Ayala, V.; Gonzalo, H.; Portero-Otin, M.; Ferrer, L.; Pamplona, R. Lipidomics of human brain aging and Alzheimer’s disease pathology. Int. Rev. Neurobiol. 2015, 122, 133–189. [Google Scholar] [PubMed]
- Kim, M.; Nevado-Holgado, A.; Whiley, L.; Snowden, S.G.; Soininen, H.; Kloszewska, I.; Mecocci, P.; Tsolaki, M.; Vellas, B.; Thambisetty, M.; et al. Association between Plasma Ceramides and Phosphatidylcholines and Hippocampal Brain Volume in Late Onset Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 60, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Simpson, B.N.; Kim, M.; Chuang, Y.-F.; Beason-Held, L.; Kitner-Triolo, M.; Kraut, M.; Lirette, S.T.; Windham, B.G.; E Griswold, M.; Legido-Quigley, C.; et al. Blood metabolite markers of cognitive performance and brain function in aging. Br. J. Pharmacol. 2016, 36, 1212–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-Y.; Sun, Y.-P.; Luo, Y.-M.; Peng, D.-H.; Li, X.; Yang, B.-Y.; Wang, Q.-H.; Kuang, H.-X. Biomarkers for the Clinical Diagnosis of Alzheimer’s Disease: Metabolomics Analysis of Brain Tissue and Blood. Front. Pharmacol. 2021, 12, 700587. [Google Scholar] [CrossRef]
- Varma, V.R.; Oommen, A.; Varma, S.; Casanova, R.; An, Y.; Andrews, R.M.; O’Brien, R.; Pletnikova, O.; Troncoso, J.C.; Toledo, J.; et al. Brain and blood metabolite signatures of pathology and progression in Alzheimer disease: A targeted metabolomics study. PLoS Med. 2018, 15, e1002482. [Google Scholar] [CrossRef]
- Fernández-Irigoyen, J.; Cartas-Cejudo, P.; Iruarrizaga-Lejarreta, M.; Santamaría, E. Alteration in the Cerebrospinal Fluid Lipidome in Parkinson’s Disease: A Post-Mortem Pilot Study. Biomedicines 2021, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, I.; Rui, B.; Khan, J.; Datta, I.; Giri, S. An emerging potential of metabolomics in multiple sclerosis: A comprehensive overview. Cell. Mol. Life Sci. 2021, 78, 3181–3203. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Strom, A.; Straßburger, K.; Knebel, B.; Bönhof, G.J.; Kotzka, J.; Szendroedi, J.; Roden, M.; for the German Diabetes Study Group. Association of cardiac autonomic dysfunction with higher levels of plasma lipid metabolites in recent-onset type 2 diabetes. Diabetologia 2021, 64, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-H.; Zheng, K.I.; Wang, X.-D.; Qiao, J.; Li, Y.-Y.; Zhang, L.; Zheng, M.-H.; Wu, J. LC-MS-based lipidomic analysis in distinguishing patients with nonalcoholic steatohepatitis from nonalcoholic fatty liver. Hepatobiliary Pancreat. Dis. Int. 2021, 20, 452–459. [Google Scholar] [CrossRef]
- Vvedenskaya, O.; Rose, T.D.; Knittelfelder, O.; Palladini, A.; Wodke, J.A.H.; Schumann, K.; Ackerman, J.M.; Wang, Y.; Has, C.; Brosch, M.; et al. Non-alcoholic fatty liver disease Stratification by Liver Lipidomics. J. Lipid Res. 2021, 62, 100104. [Google Scholar] [CrossRef]
- Boone, S.; Mook-Kanamori, D.; Rosendaal, F.; Heijer, M.D.; Lamb, H.; De Roos, A.; Le Cessie, S.; Van Dijk, K.W.; De Mutsert, R. Metabolomics: A search for biomarkers of visceral fat and liver fat content. Metabolomics 2019, 15, 139. [Google Scholar] [CrossRef] [Green Version]
- Long-Smith, C.; O’Riordan, K.J.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota-Gut-Brain Axis: New Therapeutic Opportunities. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 477–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, C.; Guerri, C.; Ureña, J.; Pascual, M. Role of Microbiota-Derived Extracellular Vesicles in Gut-Brain Communication. Int. J. Mol. Sci. 2021, 22, 4235. [Google Scholar] [CrossRef] [PubMed]
- Haas-Neill, S.; Forsythe, P. A Budding Relationship: Bacterial Extracellular Vesicles in the Microbiota–Gut–Brain Axis. Int. J. Mol. Sci. 2020, 21, 8899. [Google Scholar] [CrossRef]
- Bruce-Keller, A.J.; Salbaum, J.M.; Luo, M.; Blanchard, E.; Taylor, C.; Welsh, D.A.; Berthoud, H.-R. Obese-type Gut Microbiota Induce Neurobehavioral Changes in the Absence of Obesity. Biol. Psychiatry 2015, 77, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, Q.; Wang, Y.; Sun, A.; Lin, Y.; Jin, Y.; Li, X. Fecal microbiota transplantation from chronic unpredictable mild stress mice donors affects anxiety-like and depression-like behavior in recipient mice via the gut microbiota-inflammation-brain axis. Stress 2019, 22, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xiao, W.; Li, H.; Pang, P.; Xue, F.; Wan, L.; Pei, L.; Yan, H. Metformin restores hippocampal neurogenesis and learning and memory via regulating gut microbiota in the obese mouse model. Brain Behav. Immun. 2021, 95, 68–83. [Google Scholar] [CrossRef]
- Lee, J.C.; Park, S.M.; Kim, I.Y.; Sung, H.; Seong, J.K.; Moon, M.H. High-fat diet-induced lipidome perturbations in the cortex, hippocampus, hypothalamus, and olfactory bulb of mice. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1863, 980–990. [Google Scholar] [CrossRef]
- Borg, M.L.; Omran, S.F.; Weir, J.; Meikle, P.J.; Watt, M.J. Consumption of a high-fat diet, but not regular endurance exercise training, regulates hypothalamic lipid accumulation in mice. J. Physiol. 2012, 590, 4377–4389. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T. The impact of indigenous microbes on Parkinson’s disease. Neurobiol. Dis. 2020, 135, 104426. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Hylemon, P.B.; Ridlon, J.M.; Heuman, D.M.; Daita, K.; White, M.B.; Monteith, P.; Noble, N.A.; Sikaroodi, M.; Gillevet, P.M. Colonic mucosal microbiome differs from stool microbiome in cirrhosis and hepatic encephalopathy and is linked to cognition and inflammation. Am. J. Physiol. Liver Physiol. 2012, 303, G675–G685. [Google Scholar] [CrossRef]
- Muller, E.; Algavi, Y.M.; Borenstein, E. A meta-analysis study of the robustness and universality of gut microbiome-metabolome associations. Microbiome 2021, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Olmo, M.B.; Butler, M.J.; Barrientos, R.M. Evolution of the Human Diet and Its Impact on Gut Microbiota, Immune Responses, and Brain Health. Nutrients 2021, 13, 196. [Google Scholar] [CrossRef]
- Ghosh, S.; Pramanik, S. Structural diversity, functional aspects and future therapeutic applications of human gut microbiome. Arch. Microbiol. 2021, 203, 5281–5308. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.; Fonseca, S.; Carding, S.R. Gut microbes and metabolites as modulators of blood-brain barrier integrity and brain health. Gut Microbes 2020, 11, 135–157. [Google Scholar] [CrossRef] [Green Version]
- Jurak, E.; Patyshakuliyeva, A.; Kapsokalyvas, D.; Xing, L.; van Zandvoort, M.A.; de Vries, R.P.; Gruppen, H.; Kabel, M.A. Accumulation of recalcitrant xylan in mushroom-compost is due to a lack of xylan substituent removing enzyme activities of Agaricus bisporus. Carbohydr. Polym. 2015, 132, 359–368. [Google Scholar] [CrossRef]
- Liao, Z.; Holtzapple, M.; Yan, Y.; Wang, H.; Li, J.; Zhao, B. Insights into Xylan Degradation and Haloalkaline Adaptation through Whole-Genome Analysis of Alkalitalea saponilacus, an Anaerobic Haloalkaliphilic Bacterium Capable of Secreting Novel Halostable Xylanase. Genes 2019, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Fernandez-Kim, S.-O.; Townsend, R.L.; Kruger, C.; Carmouche, R.; Newman, S.; Salbaum, J.M.; Berthoud, H.-R. Maternal obese-type gut microbiota differentially impact cognition, anxiety and compulsive behavior in male and female offspring in mice. PLoS ONE 2017, 12, e0175577. [Google Scholar] [CrossRef]
- Shi, H.; Yu, Y.; Lin, D.; Zheng, P.; Zhang, P.; Hu, M.; Wang, Q.; Pan, W.; Yang, X.; Hu, T.; et al. beta-glucan attenuates cognitive impairment via the gut-brain axis in diet-induced obese mice. Microbiome 2020, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zhang, Q.; Dong, Z.; Yan, Y.; Fu, Y.; Liu, X.; Zhao, B.; Duan, X. Phosphatidylcholine Ameliorates LPS-Induced Systemic Inflammation and Cognitive Impairments via Mediating the Gut–Brain Axis Balance. J. Agric. Food Chem. 2020, 68, 14884–14895. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, H.; Abujrais, S.; Herman, S.; Khoonsari, P.E.; Åkerfeldt, T.; Svenningsson, A.; Burman, J.; Kultima, K. Targeted metabolomics of CSF in healthy individuals and patients with secondary progressive multiple sclerosis using high-resolution mass spectrometry. Metabolomics 2020, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liang, Q.; Wu, Y.; Gao, Z.; Kobayashi, S.; Patel, J.; Li, C.; Cai, F.; Zhang, Y.; Liang, C.; et al. Comprehensive lipidomic profiling in serum and multiple tissues from a mouse model of diabetes. Metabolomics 2020, 16, 115. [Google Scholar] [CrossRef] [PubMed]
- Félix, B.; Léger, M.-E.; Albe-Fessard, D.; Marcilloux, J.-C.; Rampin, O.; Laplace, J.-P.; Duclos, A.; Fort, F.; Gougis, S.; Costa, M.; et al. Stereotaxic atlas of the pig brain. Brain Res. Bull. 1999, 49, 1–137. [Google Scholar] [CrossRef]
- Fleming, S.A.; Monaikul, S.; Mudd, A.T.; Jacob, R.; Dilger, R.N. Extraction and Dissection of the Domesticated Pig Brain. J. Vis. Exp. 2021, 2021, e62030. [Google Scholar] [CrossRef]
- Skill, N.J.; Elliott, C.M.; Ceballos, B.; Saxena, R.; Pepin, R.; Bettcher, L.; Ellensberg, M.; Raftery, D.; Maluccio, M.A.; Ekser, B.; et al. Metabolomic Characterization of Human Model of Liver Rejection Identifies Aberrancies Linked to Cyclooxygenase (COX) and Nitric Oxide Synthase (NOS). Ann. Transplant. 2019, 24, 341–349. [Google Scholar] [CrossRef]
- Barupal, D.; Fiehn, O. Chemical Similarity Enrichment Analysis (ChemRICH) as alternative to biochemical pathway mapping for metabolomic datasets. Sci. Rep. 2017, 7, 14567. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Class | Liver | Cortex | Hippocampus |
|---|---|---|---|
| Acyl-carnitines (55) * | 3 | 3 | 3 |
| Amino acids (21) | 17 | 17 | 17 |
| Biogenic Amines (21) | 11 | 7 | 7 |
| Cholesterol Esters (14) | 5 | 5 | 5 |
| Diglycerides (18) | 6 | 2 | 2 |
| Triglycerides (42) | 1 | 1 | 1 |
| Phosphatidylcholine (172) | 92 | 93 | 96 |
| Lyso-phosphatidylcholine (24) | 8 | 8 | 8 |
| Sphingomyelins (31) | 21 | 16 | 17 |
| Ceramides (9) | 2 | 1 | 1 |
| Monosaccharides (1) | 1 | 1 | 1 |
| total (408) | 167 | 154 | 158 |
| Mean Rank | 3 Serv. vs. Control | 6 Serv. vs. Control | 6 Serv. vs. 3 Serv. | Cortex Vip | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolite | Class | Control | 3 Servings | 6 Servings | FC | FDR | FC | FDR | FC | FDR | |
| Arg | Aminoacids | 13.11 * | 10.33 | 18.56 | 0.94 | 0.340 | 1.20 | 0.141 | 1.27 | 0.141 | 1.93 |
| Gln | Aminoacids | 10.56 | 13.33 | 18.11 | 1.12 | 0.730 | 1.16 | 0.056 | 1.03 | 0.581 | 1.54 |
| Met | Aminoacids | 11.11 | 12.33 | 18.56 | 0.99 | 1.000 | 1.23 | 0.073 | 1.24 | 0.285 | 1.47 |
| Thr | Aminoacids | 8.61 | 15.50 | 17.89 | 1.11 | 0.183 | 1.35 | 0.017 | 1.22 | 0.730 | 1.41 |
| Taurine | Biogenic Amines | 12.56 | 10.72 | 18.72 | 0.86 | 0.596 | 1.12 | 0.140 | 1.31 | 0.140 | 1.58 |
| DG (42:2) | Glycerides | 9.78 | 13.06 | 19.17 | 1.07 | 0.401 | 1.18 | 0.045 | 1.10 | 0.170 | 2.05 |
| PC (29:1) | Glycerophospholipids | 10.72 | 12.83 | 18.44 | 1.10 | 0.562 | 1.22 | 0.154 | 1.11 | 0.200 | 1.80 |
| LPC (20:4) | Glycerophospholipids | 10.17 | 13.28 | 18.56 | 1.04 | 0.401 | 1.13 | 0.102 | 1.09 | 0.242 | 1.74 |
| PC-O (40:4) | Glycerophospholipids | 10.89 | 13.17 | 17.94 | 1.61 | 0.436 | 1.86 | 0.217 | 1.15 | 0.217 | 1.74 |
| PC-O (34:3) | Glycerophospholipids | 11.11 | 12.78 | 18.11 | 1.17 | 0.507 | 1.38 | 0.183 | 1.18 | 0.183 | 1.64 |
| PC-O (38:5) | Glycerophospholipids | 13.22 | 11.44 | 17.33 | 0.92 | 0.605 | 1.15 | 0.387 | 1.25 | 0.387 | 1.57 |
| PC-O (40:5) | Glycerophospholipids | 12.00 | 12.11 | 17.89 | 1.07 | 1.000 | 1.13 | 0.200 | 1.06 | 0.200 | 1.49 |
| PC (33:5) | Glycerophospholipids | 10.11 | 13.94 | 17.94 | 1.07 | 0.331 | 1.14 | 0.125 | 1.06 | 0.331 | 1.46 |
| PC (34:2) | Glycerophospholipids | 11.17 | 13.06 | 17.78 | 0.99 | 0.627 | 1.08 | 0.278 | 1.09 | 0.324 | 1.46 |
| PC (31:1) | Glycerophospholipids | 12.72 | 11.33 | 17.94 | 0.99 | 0.626 | 1.04 | 0.200 | 1.05 | 0.200 | 1.45 |
| SM (38:2) | Sphingolipids | 11.11 | 12.22 | 18.67 | 1.02 | 0.796 | 1.18 | 0.141 | 1.15 | 0.141 | 2.14 |
| SM (32:1) | Sphingolipids | 10.11 | 12.89 | 19.00 | 1.03 | 0.387 | 1.49 | 0.094 | 1.44 | 0.115 | 2.12 |
| SM (34:2) | Sphingolipids | 11.89 | 11.56 | 18.56 | 1.08 | 0.931 | 1.28 | 0.115 | 1.19 | 0.115 | 1.54 |
| - | - | Mean Ranks | 3 Serv. vs. Control | 6 Serv. vs. Control | 6 Serv. vs. 3 Serv. | Hippocampus VIP | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolite | Class | Control | 3 Serv. | 6 Serv. | FC | FDR | FC | FDR | FC | FDR | |
| Taurine | Biogenic Amines | 12 * | 11.56 | 18.44 | 0.82 | 0.863 | 1.26 | 0.141 | 1.54 | 0.141 | 1.48 |
| CE (19:2) | Cholesterol Esters | 16.17 | 9.56 | 16.28 | 0.85 | 0.153 | 1.06 | 0.931 | 1.24 | 0.153 | 1.86 |
| DG (38:5) | Glycerides | 14.67 | 9.78 | 17.56 | 0.79 | 0.285 | 1.64 | 0.436 | 2.08 | 0.151 | 1.83 |
| PC-O (40:3) | Glycerophospholipids | 11.33 | 10.44 | 20.22 | 0.85 | 0.48 | 2.08 | 0.008 | 2.46 | 0.063 | 2.24 |
| PC (39:3) | Glycerophospholipids | 14.39 | 10.22 | 17.39 | 0.86 | 0.376 | 1.00 | 0.401 | 1.17 | 0.232 | 1.96 |
| PC (41:4) | Glycerophospholipids | 13.56 | 10.78 | 17.67 | 0.78 | 0.436 | 1.25 | 0.387 | 1.60 | 0.282 | 1.80 |
| PC (39:4) | Glycerophospholipids | 14.89 | 10.83 | 16.28 | 0.89 | 0.465 | 1.05 | 0.757 | 1.17 | 0.465 | 1.67 |
| PC (33:2) | Glycerophospholipids | 11.94 | 11.89 | 18.17 | 1.03 | 1.000 | 1.51 | 0.183 | 1.47 | 0.183 | 1.66 |
| PC (42:4) | Glycerophospholipids | 17.22 | 10.17 | 14.61 | 0.83 | 0.19 | 0.97 | 0.536 | 1.17 | 0.404 | 1.65 |
| PC (36:3) | Glycerophospholipids | 14.67 | 10.56 | 16.78 | 0.80 | 0.426 | 0.99 | 0.594 | 1.24 | 0.333 | 1.61 |
| PC (37:1) | Glycerophospholipids | 13.22 | 11.61 | 17.17 | 0.51 | 0.666 | 1.27 | 0.434 | 2.51 | 0.434 | 1.61 |
| PC (38:2) | Glycerophospholipids | 14.44 | 10.22 | 17.33 | 0.74 | 0.35 | 1.05 | 0.401 | 1.41 | 0.255 | 1.52 |
| PC (37:2) | Glycerophospholipids | 14.33 | 10.89 | 16.78 | 0.76 | 0.436 | 1.17 | 0.436 | 1.54 | 0.436 | 1.50 |
| PC (36:1) | Glycerophospholipids | 13.94 | 11.22 | 16.83 | 0.80 | 0.387 | 1.07 | 0.387 | 1.34 | 0.387 | 1.49 |
| PC (39:5) | Glycerophospholipids | 20.72 | 9.72 | 11.56 | 0.84 | 0.009 | 0.88 | 0.032 | 1.05 | 0.723 | 1.35 |
| PC (42:6) | Glycerophospholipids | 19.89 | 11.33 | 10.78 | 0.82 | 0.024 | 0.75 | 0.077 | 0.92 | 0.595 | 1.00 |
| PC-O (33:3) | Glycerophospholipids | 13.22 | 11.00 | 17.78 | 0.87 | 0.436 | 1.58 | 0.242 | 1.83 | 0.242 | 1.48 |
| PC (31:1) | Glycerophospholipids | 12.61 | 12.67 | 16.72 | 0.99 | 1.000 | 1.06 | 0.510 | 1.07 | 0.510 | 1.46 |
| PC-O (33:4) | Glycerophospholipids | 13.22 | 11.44 | 17.33 | 0.85 | 0.596 | 1.13 | 0.387 | 1.34 | 0.387 | 1.45 |
| SM (38:2) | Sphingolipids | 13.11 | 11.00 | 17.89 | 0.99 | 0.536 | 1.33 | 0.277 | 1.34 | 0.277 | 1.87 |
| SM (41:2) | Sphingolipids | 12.44 | 11.28 | 18.28 | 0.74 | 0.791 | 1.38 | 0.200 | 1.87 | 0.200 | 1.81 |
| SM (40:2) | Sphingolipids | 12.11 | 11.39 | 18.50 | 0.92 | 0.73 | 1.58 | 0.153 | 1.71 | 0.153 | 1.74 |
| SM (43:2) | Sphingolipids | 13.00 | 11.22 | 17.78 | 0.95 | 0.605 | 1.44 | 0.285 | 1.51 | 0.285 | 1.69 |
| SM (42:2) | Sphingolipids | 13.00 | 10.89 | 18.11 | 0.84 | 0.536 | 1.48 | 0.236 | 1.77 | 0.231 | 1.68 |
| - | - | Mean Rank | 3 Serv. vs. Control | 6 Serv. vs. Control | 6 Serv. vs. 3 Serv. | Liver VIP- | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolite | Class | Control | 3 Serv. | 6 Serv. | FC | FDR | FC | FDR | FC | FDR | |
| AC (4:0-OH) | Acylcarnitines | 10.25 * | 16.17 | 13.72 | 1.12 | 0.275 | 1.13 | 0.627 | 1.01 | 0.627 | 1.54 |
| Serotonin | Biogenic Amines | 9.19 | 20.89 | 9.94 | 2.40 | 0.007 | 1.09 | 0.700 | 0.45 | 0.001 | 2.07 |
| CE (22:5) | Cholesterol Esters | 17.63 | 11.44 | 11.89 | 0.86 | 0.204 | 0.86 | 0.204 | 1.00 | 0.930 | 1.49 |
| DG (36:2) | Glycerides | 18.00 | 9.61 | 13.39 | 0.60 | 0.162 | 0.88 | 0.222 | 1.48 | 0.222 | 1.67 |
| PC (31:1) | Glycerophospholipids | 20.00 | 9.33 | 11.89 | 0.61 | 0.024 | 0.62 | 0.031 | 1.03 | 0.427 | 2.31 |
| PC (35:1) | Glycerophospholipids | 19.31 | 9.39 | 12.44 | 0.64 | 0.017 | 0.82 | 0.125 | 1.28 | 0.453 | 2.26 |
| PC-O (33:4) | Glycerophospholipids | 19.38 | 8.56 | 13.22 | 0.73 | 0.017 | 0.77 | 0.112 | 1.05 | 0.161 | 2.23 |
| PC-O (35:4) | Glycerophospholipids | 19.69 | 9.78 | 11.72 | 0.46 | 0.029 | 0.62 | 0.047 | 1.33 | 0.574 | 2.18 |
| PC (46:2) | Glycerophospholipids | 19.63 | 9.78 | 11.78 | 0.39 | 0.034 | 0.47 | 0.034 | 1.19 | 0.418 | 2.16 |
| LPC (15:0) | Glycerophospholipids | 19.25 | 10.67 | 11.22 | 0.67 | 0.045 | 0.83 | 0.045 | 1.23 | 0.860 | 2.14 |
| PC (39:3) | Glycerophospholipids | 18.63 | 8.00 | 14.44 | 0.72 | 0.024 | 0.85 | 0.228 | 1.18 | 0.095 | 2.12 |
| PC (33:1) | Glycerophospholipids | 18.88 | 9.44 | 12.78 | 0.66 | 0.024 | 0.74 | 0.209 | 1.12 | 0.436 | 2.07 |
| PC (35:4) | Glycerophospholipids | 20.50 | 10.00 | 10.78 | 0.86 | 0.014 | 0.85 | 0.021 | 0.99 | 0.930 | 1.98 |
| PC-O (33:3) | Glycerophospholipids | 17.88 | 10.11 | 13.00 | 0.72 | 0.139 | 0.75 | 0.300 | 1.04 | 0.436 | 1.91 |
| PC (37:1) | Glycerophospholipids | 19.00 | 10.67 | 11.44 | 0.75 | 0.070 | 0.84 | 0.070 | 1.11 | 0.863 | 1.86 |
| PC (32:6) | Glycerophospholipids | 7.19 | 15.89 | 16.72 | 1.61 | 0.027 | 1.44 | 0.027 | 0.89 | 0.791 | 1.84 |
| PC (38:2) | Glycerophospholipids | 8.00 | 17.72 | 14.17 | 2.00 | 0.037 | 1.86 | 0.152 | 0.93 | 0.331 | 1.81 |
| PC (31:0) | Glycerophospholipids | 17.50 | 11.78 | 11.67 | 0.97 | 0.290 | 0.87 | 0.275 | 0.89 | 0.931 | 1.65 |
| PC (33:0) | Glycerophospholipids | 17.63 | 10.28 | 13.06 | 0.79 | 0.178 | 0.92 | 0.354 | 1.17 | 0.453 | 1.64 |
| PC (30:0) | Glycerophospholipids | 17.88 | 11.33 | 11.78 | 0.84 | 0.171 | 0.76 | 0.171 | 0.90 | 0.931 | 1.60 |
| PC (32:1) | Glycerophospholipids | 17.81 | 10.39 | 12.78 | 0.77 | 0.103 | 0.89 | 0.402 | 1.16 | 0.659 | 1.48 |
| SM (43:1) | Sphingolipids | 20.50 | 9.67 | 11.11 | 0.61 | 0.008 | 0.64 | 0.008 | 1.06 | 0.605 | 2.43 |
| SM (42:1) | Sphingolipids | 20.38 | 11.17 | 9.72 | 0.72 | 0.018 | 0.63 | 0.018 | 0.88 | 0.666 | 2.16 |
| Cer (42:1) | Sphingolipids | 20.13 | 8.78 | 12.33 | 0.61 | 0.007 | 0.78 | 0.041 | 1.28 | 0.297 | 2.16 |
| SM (41:1) | Sphingolipids | 17.88 | 11.78 | 11.33 | 0.92 | 0.171 | 0.86 | 0.171 | 0.93 | 0.931 | 1.81 |
| Cer (42:2) | Sphingolipids | 17.63 | 9.89 | 13.44 | 0.65 | 0.223 | 0.75 | 0.258 | 1.17 | 0.258 | 1.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solano-Aguilar, G.I.; Lakshman, S.; Jang, S.; Gupta, R.; Molokin, A.; Schroeder, S.G.; Gillevet, P.M.; Urban, J.F., Jr. The Effects of Consuming White Button Mushroom Agaricus bisporus on the Brain and Liver Metabolome Using a Targeted Metabolomic Analysis. Metabolites 2021, 11, 779. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110779
Solano-Aguilar GI, Lakshman S, Jang S, Gupta R, Molokin A, Schroeder SG, Gillevet PM, Urban JF Jr. The Effects of Consuming White Button Mushroom Agaricus bisporus on the Brain and Liver Metabolome Using a Targeted Metabolomic Analysis. Metabolites. 2021; 11(11):779. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110779
Chicago/Turabian StyleSolano-Aguilar, Gloria I., Sukla Lakshman, Saebyeol Jang, Richi Gupta, Aleksey Molokin, Steven G. Schroeder, Patrick M. Gillevet, and Joseph F. Urban, Jr. 2021. "The Effects of Consuming White Button Mushroom Agaricus bisporus on the Brain and Liver Metabolome Using a Targeted Metabolomic Analysis" Metabolites 11, no. 11: 779. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11110779