
The Linoleic Acid Content of the Stratum Corneum of Ichthyotic Golden Retriever Dogs Is Reduced as Compared to Healthy Dogs and a Significant Part Is Oxidized in Both Free and Esterified Forms

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
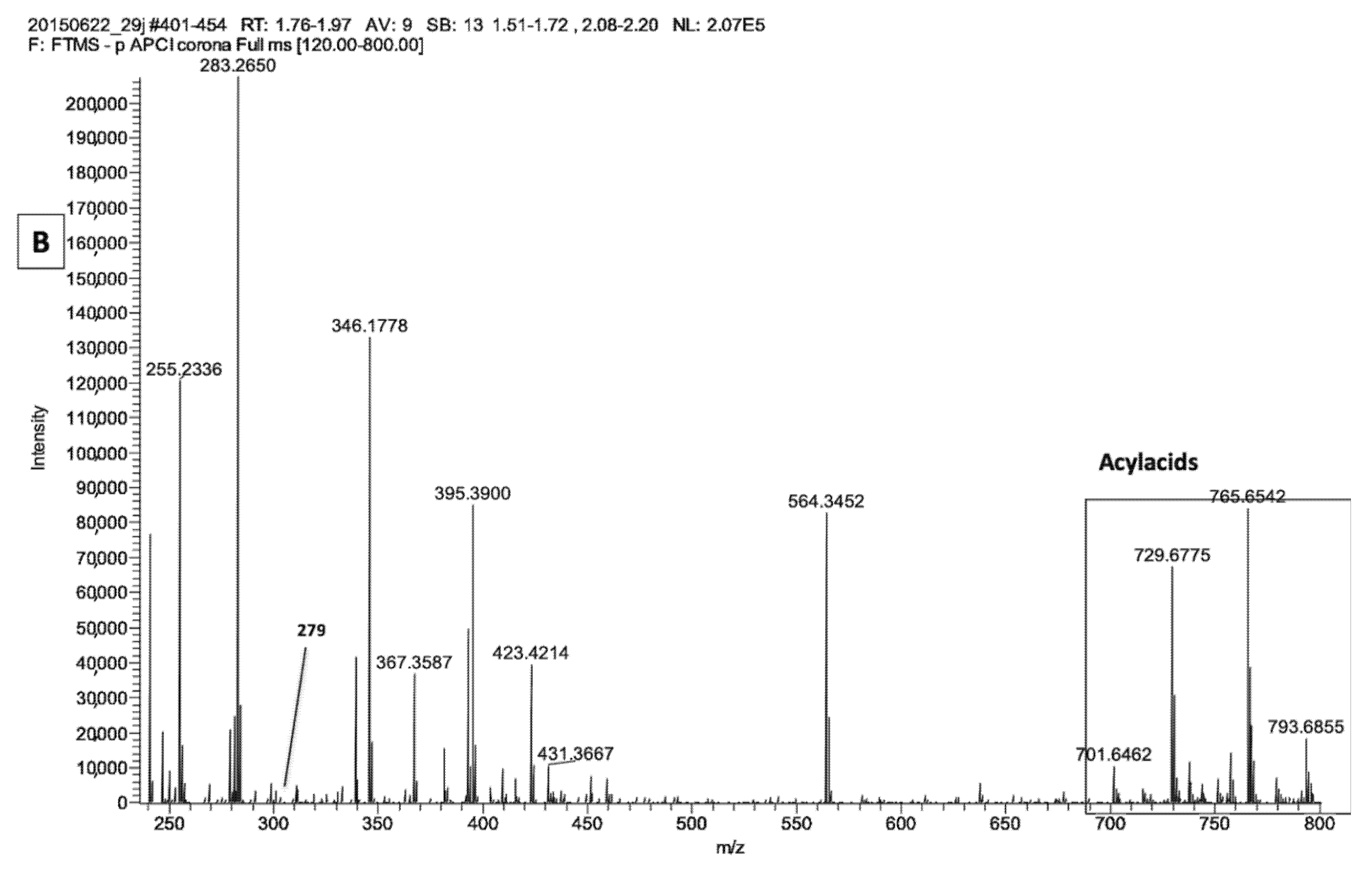
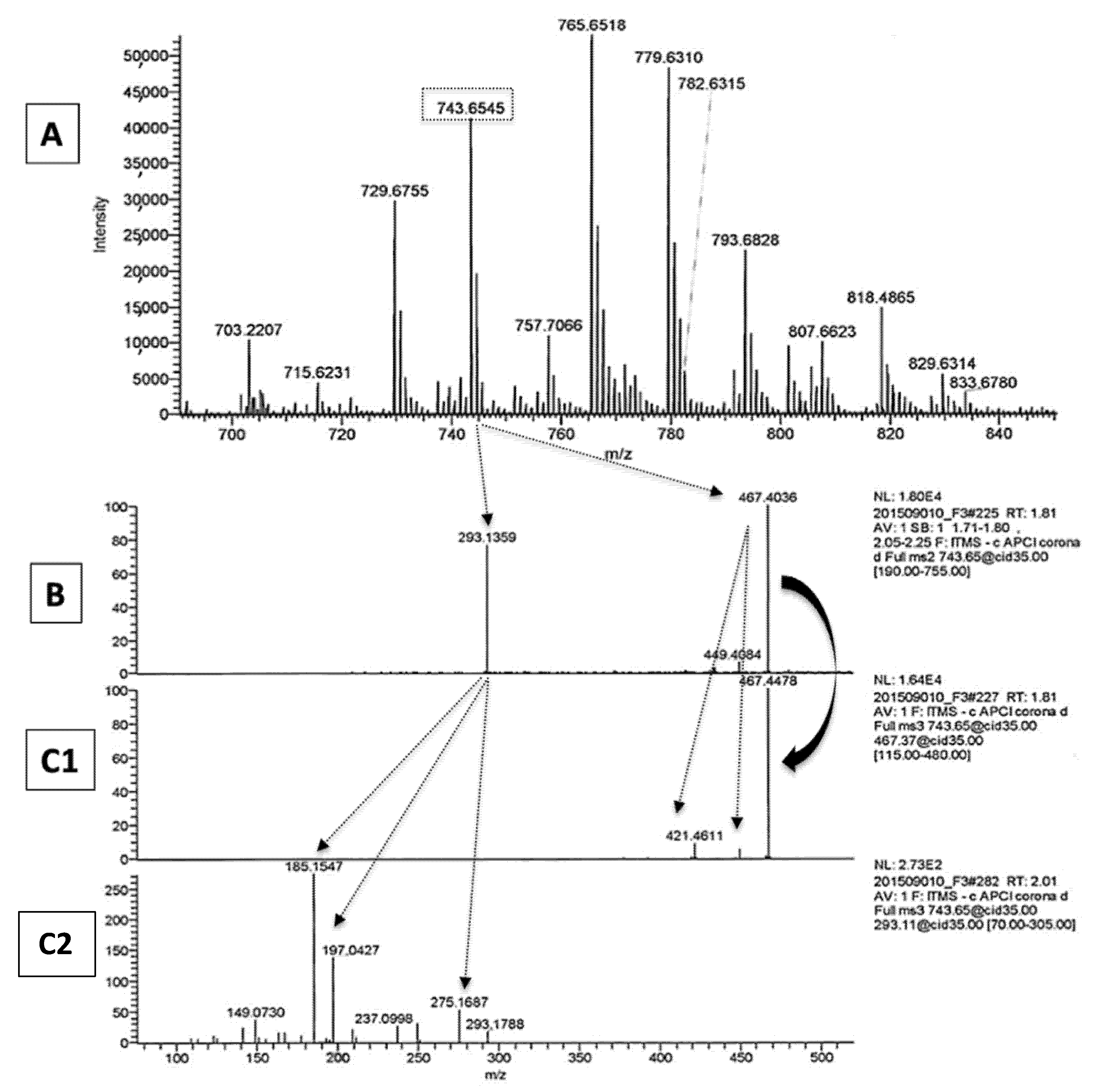
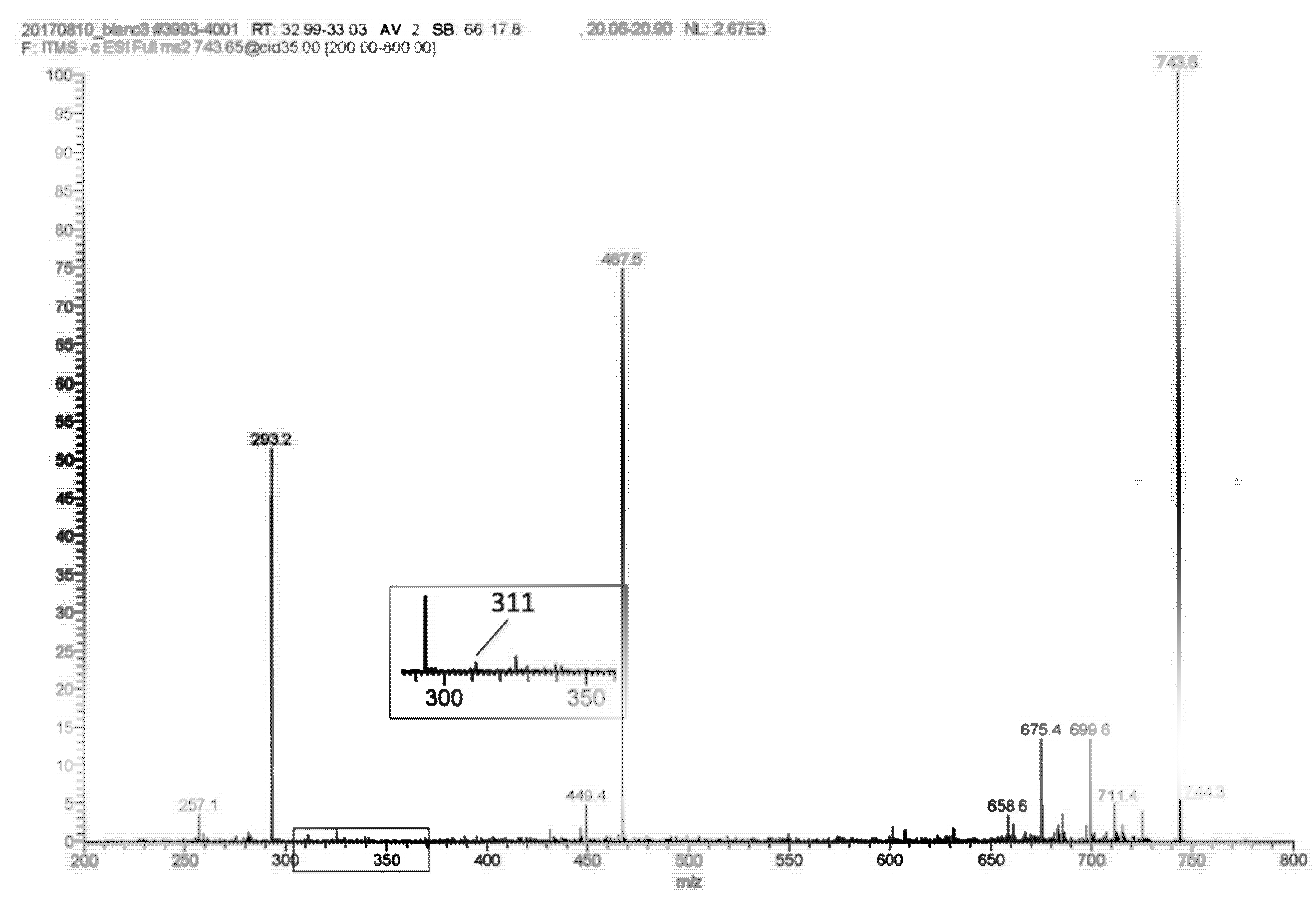
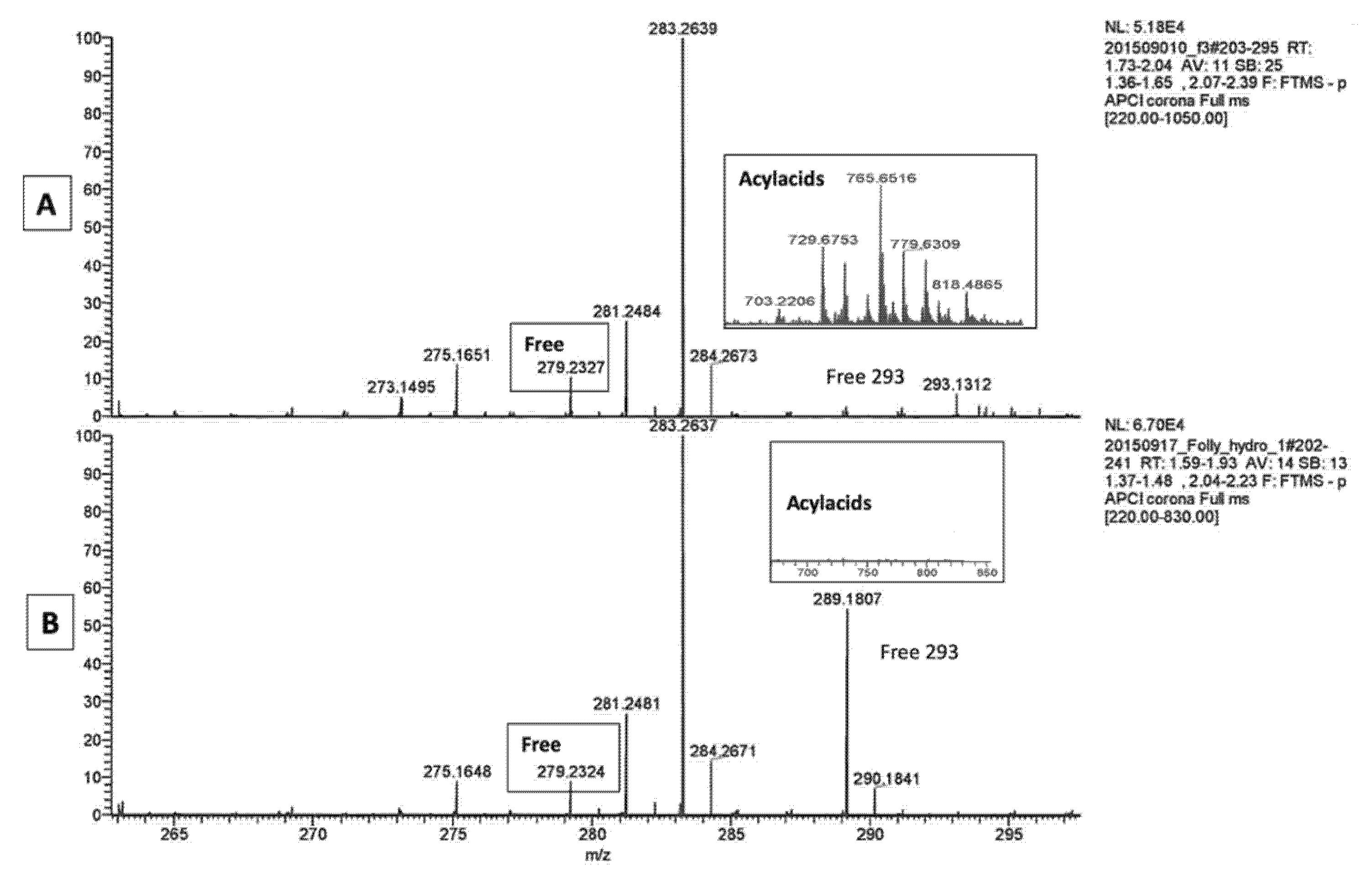
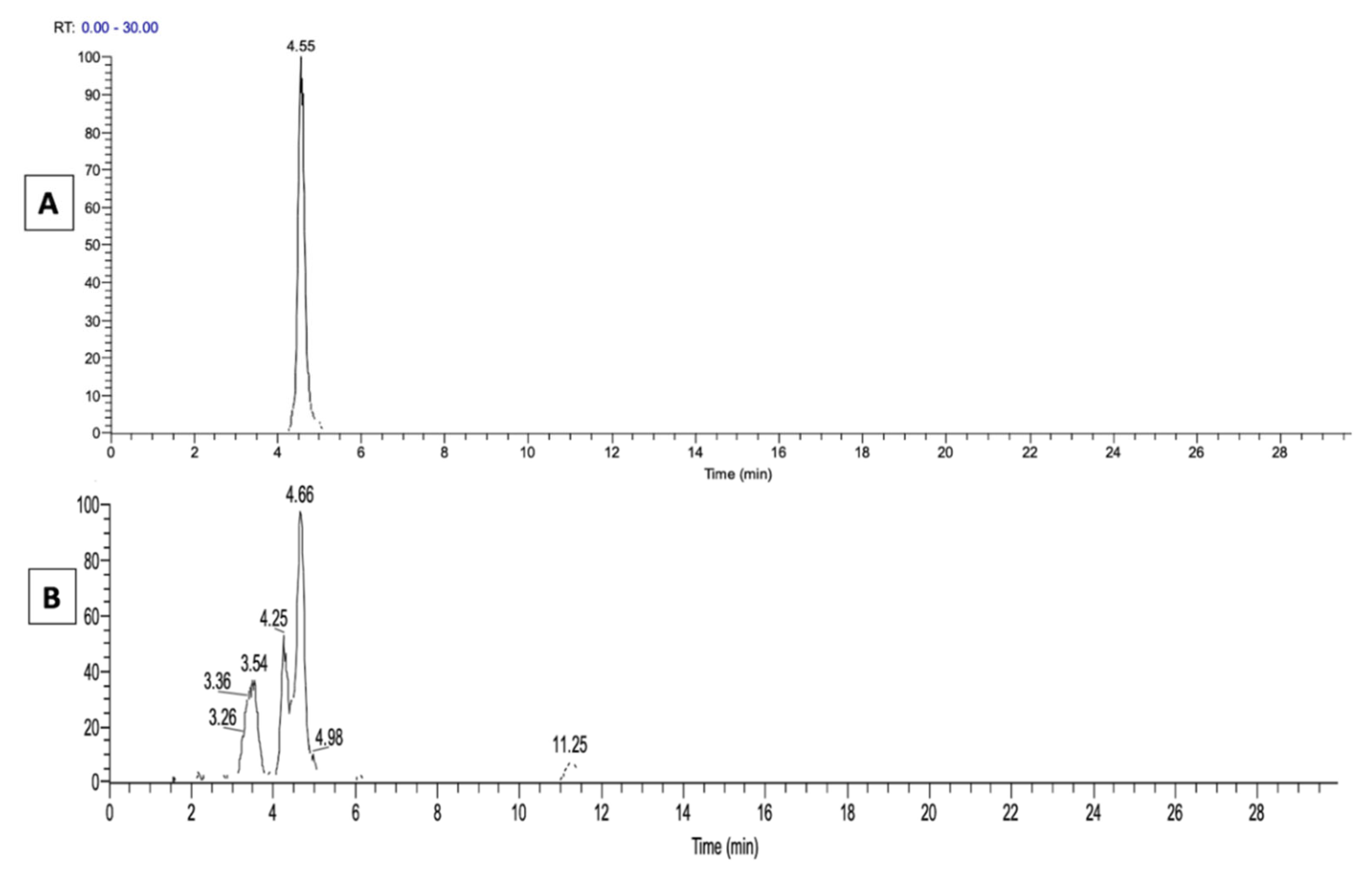
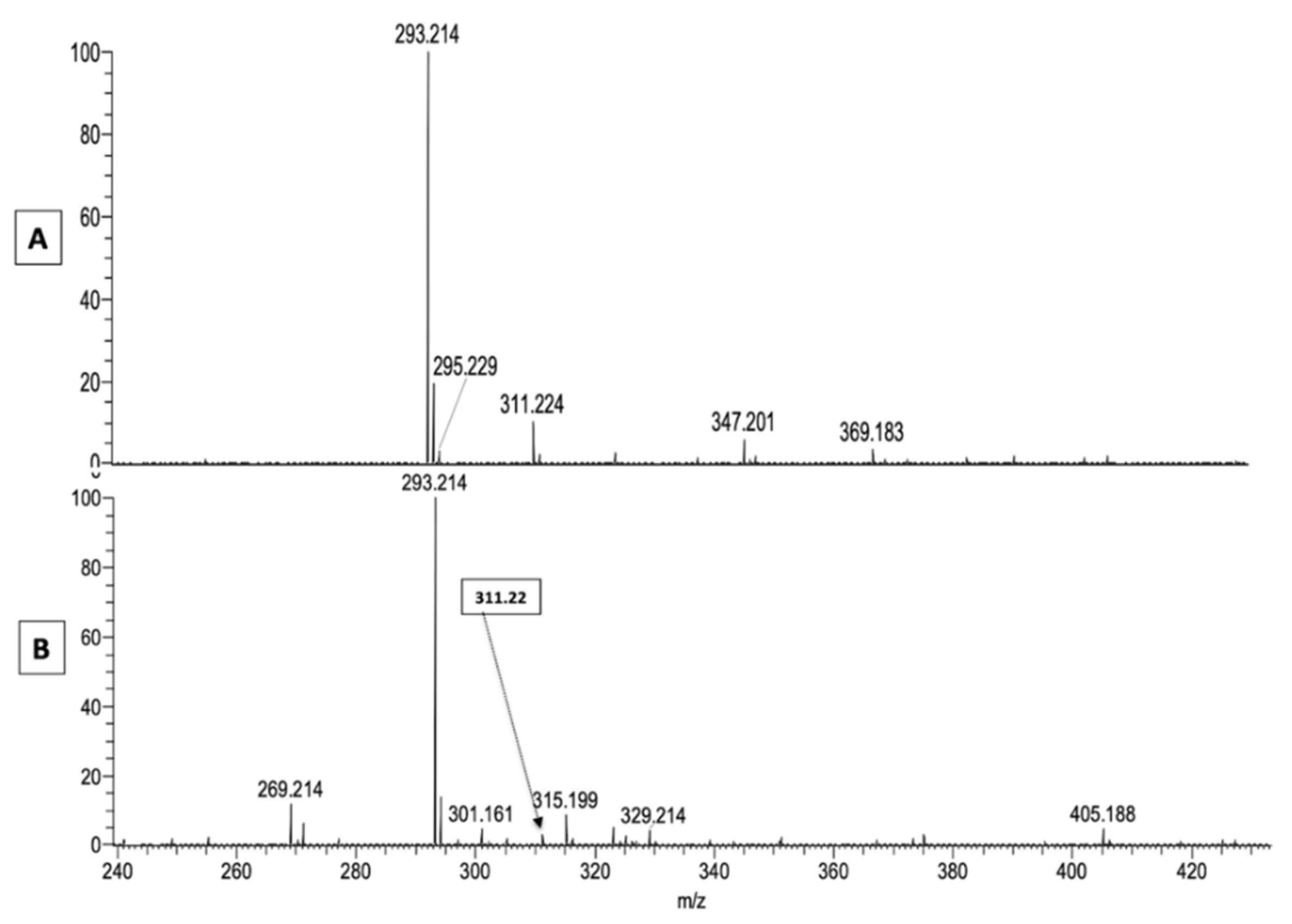
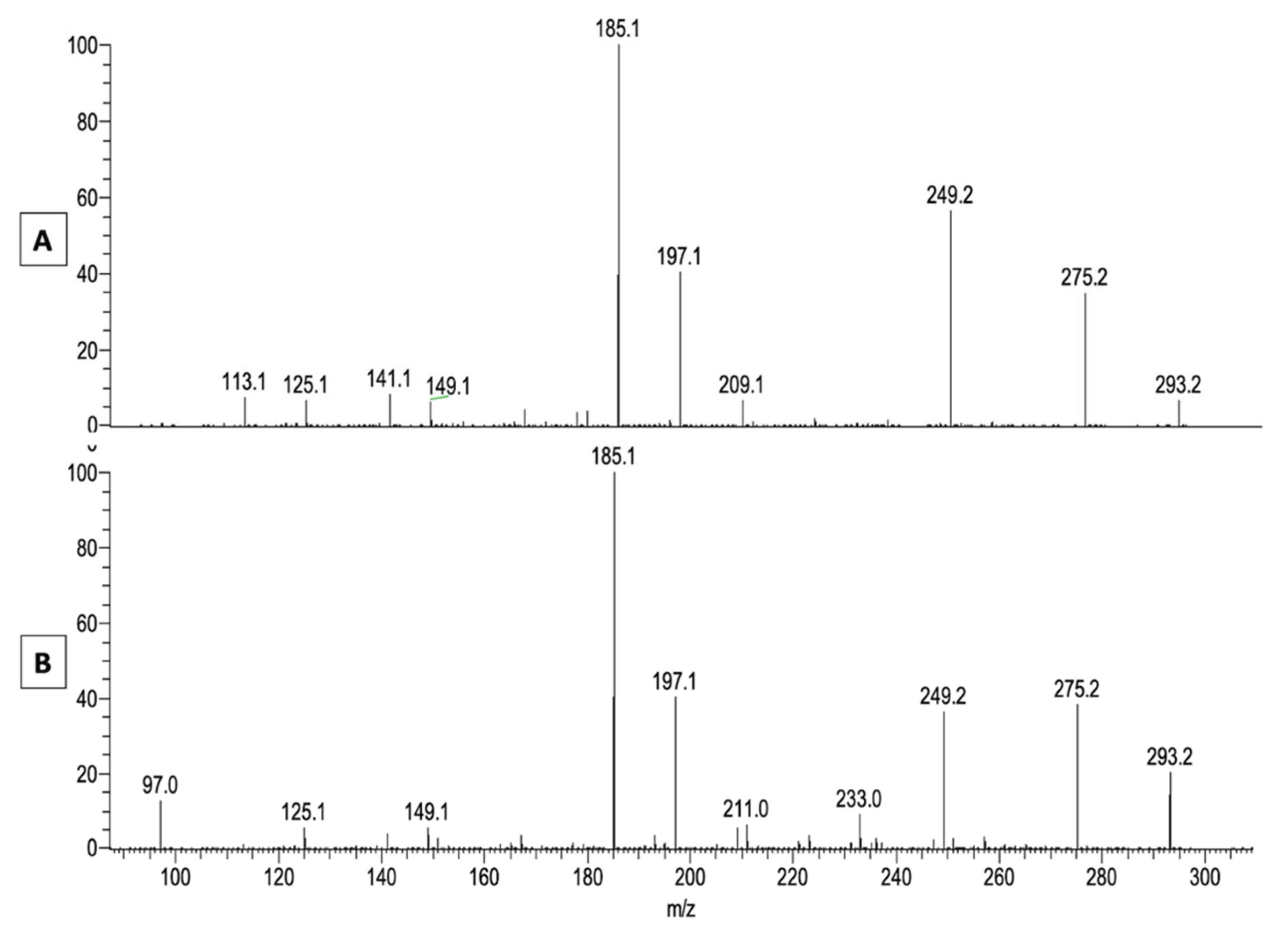
2.1. LA Acylacid LC-MS-APCI Identification and MS2 Fragmentation
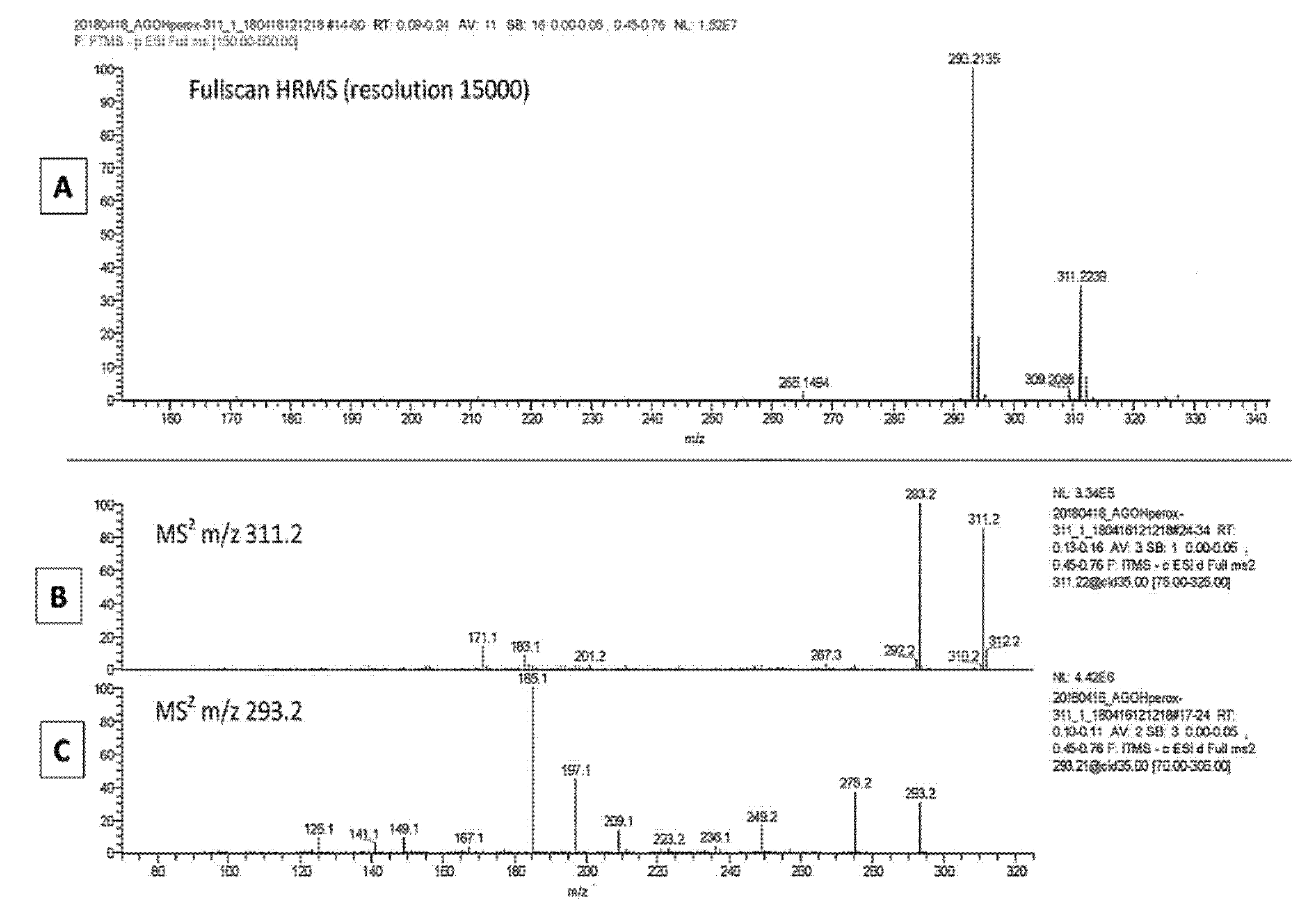
2.2. The LA Hydroperoxide Identification
3. Discussion
4. Materials and Methods
4.1. Tissues
4.2. Lipid Extraction and Analysis
4.3. Analysis of Free Fatty Acids by Liquid Chromatography-Mass Spectrometry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cunnane, S.; Anderson, M. Pure linoleate deficiency in the rat: Influence on growth, accumulation of n-6 polyunsaturates, and (14C) linoleate oxidation. J. Lipid Res. 1997, 38, 805–812. [Google Scholar] [CrossRef]
- Popa, I.; Watson, A.; Sogadi, A.; Butowski, C.; Allaway, D.; Portoukalian, J. Linoleate-enriched diet increases both linoleic acid esterified to omega hydroxy very long chain fatty acids and free ceramides of canine stratum corneum without effect on protein-bound ceramides and skin barrier function. Arch. Dermatol. Res. 2018, 310, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowser, P.A.; Nugteren, D.H.; White, R.J.; Houtsmuller, U.M.; Prottey, C. Identification, isolation and characterization of epidermal lipids containing linoleic acid. Biochim. Biophys. Acta 1985, 834, 419–428. [Google Scholar] [CrossRef]
- Wertz, P.W.; Downing, D.T. Acylglucosylceramides of pig epidermis: Structure determination. J. Lipid Res. 1983, 24, 753–758. [Google Scholar] [CrossRef]
- Melton, J.L.; Wertz, P.W.; Swartzendruber, D.C.; Downing, D.T. Effects of essential fatty acid deficiency on epidermal O-acylsphingolipids and transepidermal water loss in young pigs. Biochim. Biophys. Acta 1997, 921, 191–197. [Google Scholar] [CrossRef]
- Cadiergues, M.-A.; Patel, A.; Shearer, D.H.; Fermor, R.; Miah, S.; Hendricks, A. Cornification defect in the Golden retriever: Clinical, histopathological, ultrastructural and genetic characterisation. Vet. Dermatol. 2008, 19, 120–129. [Google Scholar] [CrossRef]
- Grall, A.; Guaguère, E.; Planchais, S.; Grond, S.; Bourrat, E.; Hausser, I.; Hitte, C.; LeGallo, M.; Derbois, C.; Kim, G.J.; et al. PNPLA1 mutations cause autosomal recessive congenital ichthyosis in Golden retriever dogs and humans. Nat. Genet. 2012, 44, 140–147. [Google Scholar] [CrossRef]
- Ohno, Y.; Kamiyama, N.; Nakamichi, S.; Kihara, A. PNPLA1 is a transacylase essential for the generation of the skin barrier lipid ω-O-acylceramide. Nat. Commun. 2017, 8, 14610. [Google Scholar] [CrossRef] [PubMed]
- Eckl, K.-M.; Tidhar, R.; Thiele, H.; Oji, V.; Hausser, I.; Brodesser, S.; Preil, M.-L.; Önal-Akan, A.; Stock, F.; Muller, D.; et al. Impaired epidermal ceramide synthesis causes autosomal recessive congenital ichthyosis and reveals the importance of ceramide acyl chain length. J. Investig. Dermatol. 2013, 133, 2202–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grond, S.; Eichmann, T.O.; Dubrac, S.; Kolb, D.; Schmuth, M.; Fischer, J.; Crumrine, D.; Elias, P.M.; Haemmerle, G.; Zechner, R.; et al. PNPLA1 deficiency in mice and humans leads to a defect in the synthesis of omega-O-acylceramides. J. Investig. Dermatol. 2017, 137, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Hirabayashi, T.; Anjo, T.; Kaneko, A.; Senoo, Y.; Shibata, A.; Takama, H.; Yokoyama, K.; Nishito, Y.; Ono, T.; Taya, C.; et al. PNPLA1 has a crucial role in skin barrier function by directing acylceramide biosynthesis. Nat. Commun. 2017, 8, 14609. [Google Scholar] [CrossRef] [PubMed]
- Oliw, E.H.; Garscha, U.; Nilsson, T.; Cristea, M. Payne rearrangement during analysis of epoxyalcohols of linoleic and ω-linolenic acids by normal phase liquid chromatography with tandem mass spectrometry. Anal. Biochem. 2006, 354, 111–126. [Google Scholar] [CrossRef]
- Yu, Z.; Schneider, C.; Boeglin, W.E.; Marnett, L.J.; Brash, A.R. The lipoxygenase gene ALOXE3 implicated in skin differentiation encodes a hydroperoxide isomerase. Proc. Natl. Acad. Sci. USA 2003, 100, 9162–9167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebert, M.; Krieg, P.; Lehmann, W.D.; Marks, F.; Fürstenberger, G. Enzymic characterization of epidermis-derived 12-lipoxygenase isoenzymes. Biochem. J. 2001, 355, 97–104. [Google Scholar] [CrossRef]
- MacMillan, D.K.; Murphy, R.C. Analysis of lipid hydroperoxides and long-chain conjugated keto acids by negative ion electrospray mass spectrometry. Am. Soc. Mass Spectrom. 1995, 6, 1190–1201. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Wojtowicz, J.C.; Molai, M. Human tear film and meibum, Very long chain wax esters and (O-acyl)-omega-hydroxy fatty acids of meibum. J. Lipid Res. 2009, 50, 2471–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yin, H.; Boeglin, W.E.; Elias, P.M.; Crumrine, D.; Beier, D.R.; Brash, A.R. Lipoxygenases mediate the effect of essential fatty acid in skin barrier formation. A proposed role in releasing omega-hydroxyceramide for construction of the corneocyte lipid envelope. J. Biol. Chem. 2011, 286, 24046–24056. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-S.; Sassaki, A.; Shimada, K.; Ide, K.; Iwasaki, T.; Nishifuji, K. Effects of age, sex, and breed on the composition of free extractable ceramides in the stratum corneum of healthy dogs. Vet. Res. Commun. 2021. (Online ahead of print). [Google Scholar] [CrossRef]
- Munoz-Garcia, A.; Thomas, C.P.; Keeney, D.S.; Zheng, Y.; Brash, A.R. The importance of the lipoxygenase-hepoxilin pathway in the mammalian epidermal barrier. Biochim. Biophys. Acta 2014, 184, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Hayakawa, M.; Niki, E. Total hydroxyloctadecanoic acid as a marker for lipid peroxidation in vivo. Biofactors 2005, 24, 7–15. [Google Scholar] [CrossRef]
- Patwardhan, A.M.; Akopian, A.N.; Ruparel, N.B.; Diogenes, A.; Weintraub, S.T.; Uhlson, C.; Murphy, R.C.; Hargreaves, K.M. Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents. J. Clin. Investig. 2009, 120, 1617–1627. [Google Scholar] [CrossRef]
- Denda, M.; Fuziwara, S.; Inoue, K.; Denda, S.; Akamatsu, H.; Tomitaka, A.; Matsunaga, K. Immunoreactivity of VR1 on epidermal keratinocyte of human skin. Biochem. Biophys. Res. Commun. 2001, 285, 1250–1252. [Google Scholar] [CrossRef]
- Denda, M.; Sokabe, T.; Fukumi-Tominaga, T.; Tominaga, M. Effects of skin surface temperature on epidermal permeability barrier homeostasis. J. Investig. Dermatol. 2007, 127, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marekow, L.N.; Steinert, P.M. Ceramides are bound to structural proteins of the human foreskin epidermal cornified cell envelope. J. Biol. Chem 1998, 273, 17763–17770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheleidt, O.; Kaiser, H.W.; Sandhoff, K. Deficiency of epidermal protein-bound ω-hydroxyceramides in atopic dermatitis. J. Investig. Dermatol. 2002, 119, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Popa, I.; Pin, D.; Remoué, N.; Osta, B.; Callejon, S.; Videmont, E.; Gatto, H.; Portoukalian, J.; Haftek, M. The lipid alterations in the stratum corneum of dogs with atopic dermatitis are alleviated by topical application of a sphingolipid-containing emulsion. Clin. Exp. Dermatol. 2012, 37, 665–671. [Google Scholar] [CrossRef] [PubMed]
- León-López, A.; Fuentes-Jiménez, L.; Hernández-Fuentes, A.D.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Hydrolysed Collagen from Sheepskins as a Source of Functional Peptides with Antioxidant Activity. Int. J. Mol. Sci. 2019, 20, 3931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre-Cruz, G.; Arely León-López, A.; Verónica Cruz-Gómez, V.; Jiménez-Alvarado, R.; Aguirre-Álvarez, G. Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants 2020, 9, 181. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dogs (n = 5) | C18:2 (µg/mg Proteins) | % as Free Fatty Acid | % as Acylacid |
|---|---|---|---|
| Control dogs | 26.7 ± 14.3 | 8.6 ± 6.4 | 91.3 ± 12.6 |
| Ichthyotic dogs | 8.1 ± 5.6 | 10.2 ± 7.5 | 90.1 ± 13.8 |
| M-H | M+Cl | Formula | Fragments | Structures |
|---|---|---|---|---|
| 701.643 | 737.620 | C46H85O4 | 439; 421; 279 | (O-C18:2)-ωOH-C28:0 |
| 715.659 | 751.659 | C47H87O4 | 439; 421; 293 | (O-ketoC18:2)-ωOH-C28:0 |
| 729.674 | 765.651 | C48H89O4 | 467; 449; 279 | (O-C18:2)-ωOH-C30:0 |
| 743.637 | 779.630 | C48H88O5Cl | 467; 293 | (O-ketoC18:2)-)-ωOH-C30:0 |
| 743.693 | 779.666 | C49H92O4Cl | 481; 463; 279 | (O-C18:2)-ωOH-C31:0 |
| 783.700 | 819.700 | C52H96O4Cl | 521; 503; 279 | (O-C18:2)-ωOH-C34:1 |
| Dogs (n = 5) | % LA as Keto-Derivatives |
|---|---|
| Control dogs | 3.7 ± 0.6 |
| Control dogs fed with LA-enriched diet [2] | 3.8 ± 0.7 |
| Ichthyotic Golden Retrievers dogs | 13.9 ± 1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popa, I.; Solgadi, A.; Pin, D.; Watson, A.L.; Haftek, M.; Portoukalian, J. The Linoleic Acid Content of the Stratum Corneum of Ichthyotic Golden Retriever Dogs Is Reduced as Compared to Healthy Dogs and a Significant Part Is Oxidized in Both Free and Esterified Forms. Metabolites 2021, 11, 803. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120803
Popa I, Solgadi A, Pin D, Watson AL, Haftek M, Portoukalian J. The Linoleic Acid Content of the Stratum Corneum of Ichthyotic Golden Retriever Dogs Is Reduced as Compared to Healthy Dogs and a Significant Part Is Oxidized in Both Free and Esterified Forms. Metabolites. 2021; 11(12):803. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120803
Chicago/Turabian StylePopa, Iuliana, Audrey Solgadi, Didier Pin, Adrian L. Watson, Marek Haftek, and Jacques Portoukalian. 2021. "The Linoleic Acid Content of the Stratum Corneum of Ichthyotic Golden Retriever Dogs Is Reduced as Compared to Healthy Dogs and a Significant Part Is Oxidized in Both Free and Esterified Forms" Metabolites 11, no. 12: 803. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120803