
Metabolic Fluctuations in the Human Stool Obtained from Blastocystis Carriers and Non-Carriers

 , , , , ,
, , , , ,
Abstract
:1. Introduction
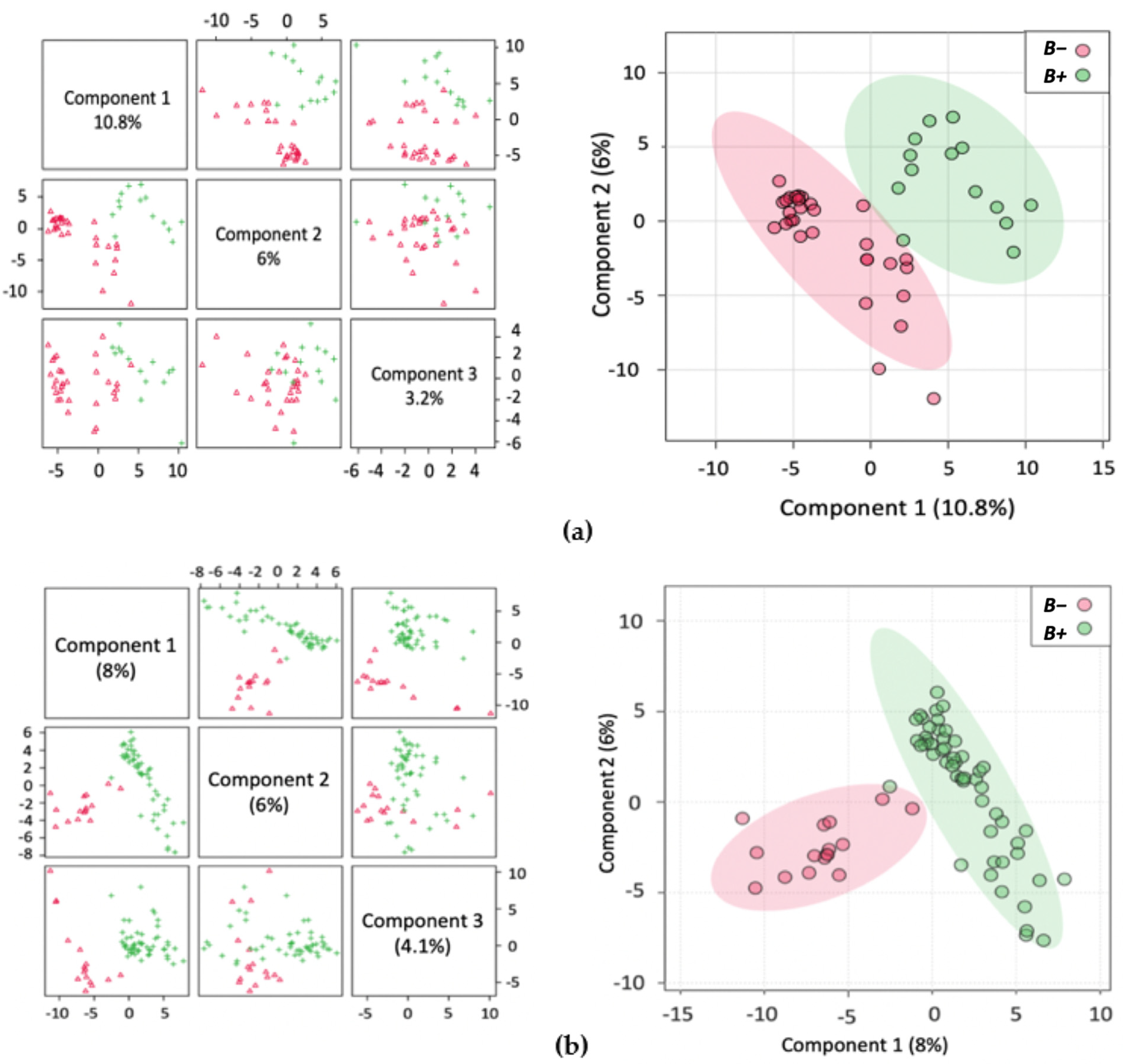
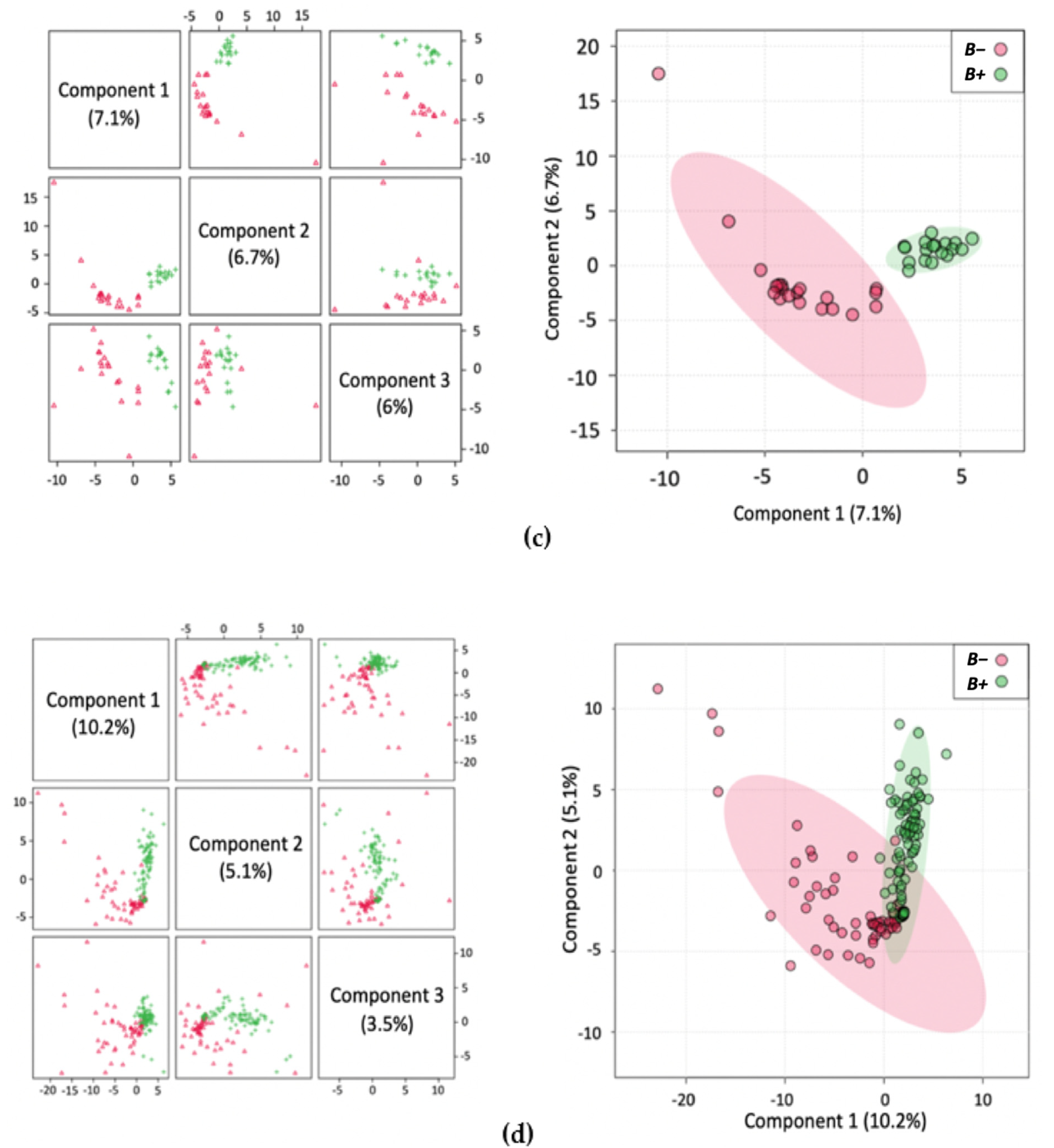
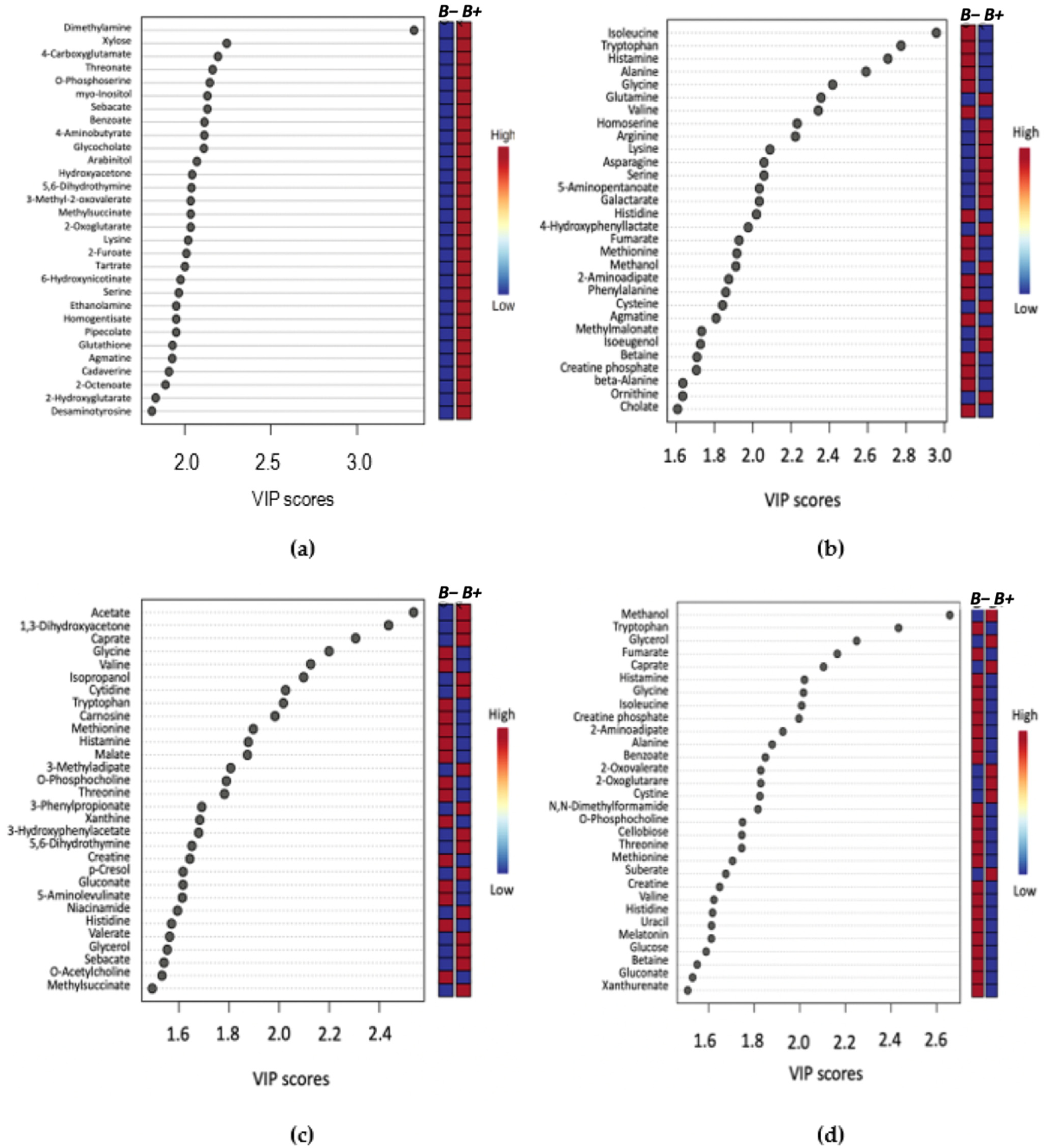
2. Results
2.1. Metabolite Profiles
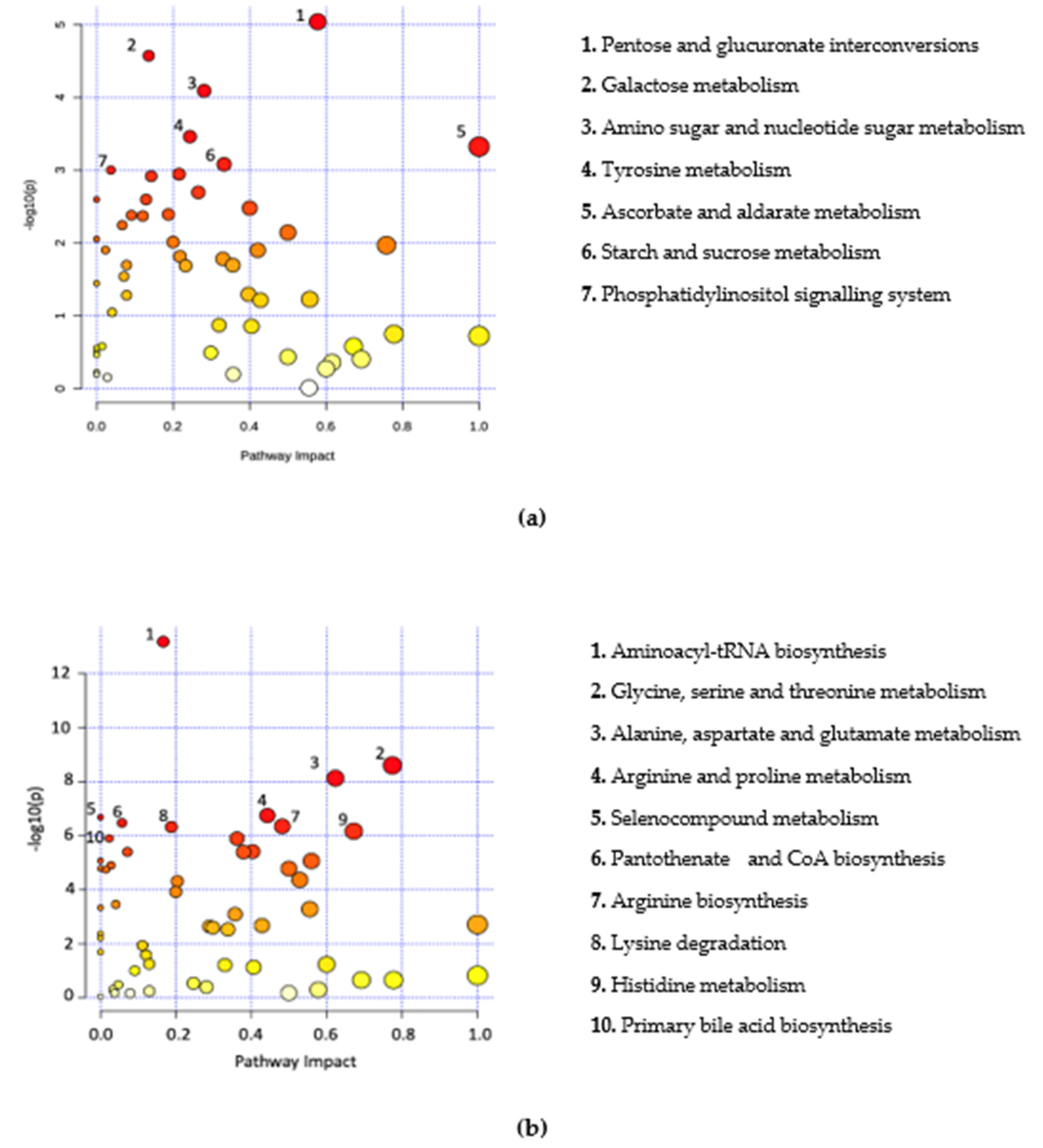
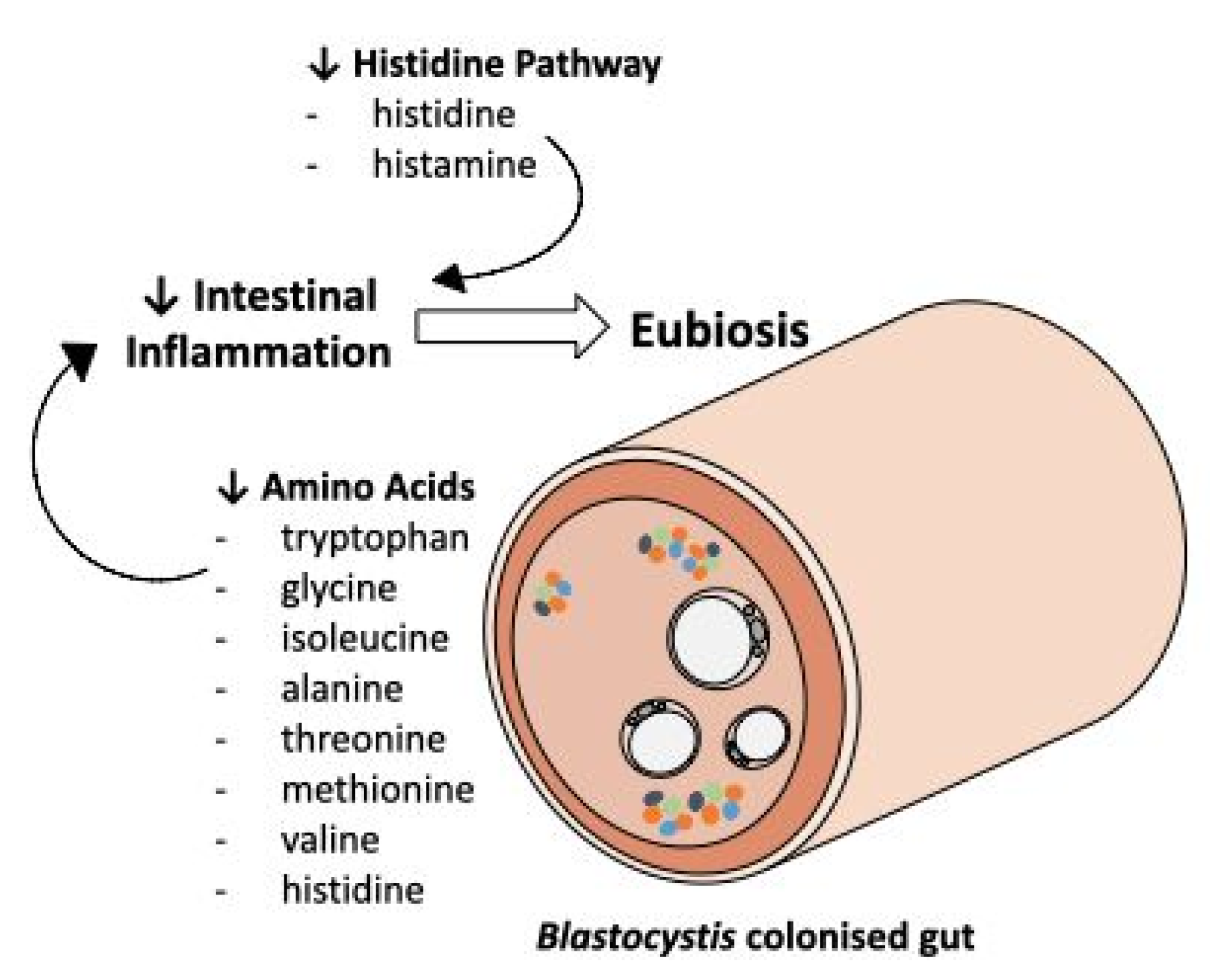
2.2. Metabolic Profile and Pathway Analysis of Blastocystis Infected Individuals
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Sample Collection
4.3. Blastocystis Detection
4.4. Sample Preparation
4.5. NMR Spectroscopy
4.6. Data Processing and Statistical Analysis
4.6.1. Analysis of Spectra
4.6.2. Grouping and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basolo, A.; Hohenadel, M.; Ang, Q.Y.; Piaggi, P.; Heinitz, S.; Walter, M.; Walter, P.; Parrington, S.; Trinidad, D.D.; von Schwartzenberg, R.J.; et al. Effects of Underfeeding and Oral Vancomycin on Gut Microbiome and Nutrient Absorption in Humans. Nat. Med. 2020, 26, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome—A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [Green Version]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut Microbiota Functions: Metabolism of Nutrients and Other Food Components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A New Genomic Blueprint of the Human Gut Microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Pasolli, E.; Asnicar, F.; Manara, S.; Zolfo, M.; Karcher, N.; Armanini, F.; Beghini, F.; Manghi, P.; Tett, A.; Ghensi, P.; et al. Extensive Unexplored Human Microbiome Diversity Revealed by Over 150,000 Genomes from Metagenomes Spanning Age, Geography, and Lifestyle. Cell 2019, 176, 649–662.e20. [Google Scholar] [CrossRef] [Green Version]
- Chabé, M.; Lokmer, A.; Ségurel, L. Gut Protozoa: Friends or Foes of the Human Gut Microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.L.; Gilchrist, C.A.; Lynn, T.C.; Petri, W.A. Parasitic Protozoa and Interactions with the Host Intestinal Microbiota. Infect Immun. 2017, 85, e00101-17. [Google Scholar] [CrossRef] [Green Version]
- Hicks, S.J.; Theodoropoulos, G.; Carrington, S.D.; Corfield, A.P. The Role of Mucins in Host–Parasite Interactions. Part I—Protozoan Parasites. Parasitol. Today 2000, 16, 476–481. [Google Scholar] [CrossRef]
- Jenkins, T.P.; Formenti, F.; Castro, C.; Piubelli, C.; Perandin, F.; Buonfrate, D.; Otranto, D.; Griffin, J.L.; Krause, L.; Bisoffi, Z.; et al. A Comprehensive Analysis of the Faecal Microbiome and Metabolome of Strongyloides Stercoralis Infected Volunteers from a Non-Endemic Area. Sci. Rep. 2018, 8, 15651. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.M.; Graham, A.L.; Knowles, S.C.L. Parasite-Microbiota Interactions With the Vertebrate Gut: Synthesis Through an Ecological Lens. Front. Microbiol. 2018, 9, 843. [Google Scholar] [CrossRef] [Green Version]
- Boorom, K.F.; Smith, H.; Nimri, L.; Viscogliosi, E.; Spanakos, G.; Parkar, U.; Li, L.H.; Zhou, X.N.; Ok, L.Z.; Leelayoova, S.; et al. Oh My Aching Gut: Irritable Bowel Syndrome, Blastocystis, and Asymptomatic Infection. Parasites Vectors 2008, 1, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, R.T.; El-Bali, M.A.; Mohamed, A.A.; Abdel-Fatah, M.A.; EL-Malky, M.A.; Mowafy, N.M.; Zaghlool, D.A.; Bakri, R.A.; Al-Harthi, S.A. Subtyping of Blastocystis Sp. Isolated from Symptomatic and Asymptomatic Individuals in Makkah, Saudi Arabia. Parasites Vectors 2017, 10, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, D.; van Hal, S.; Marriott, D.; Ellis, J.; Harkness, J. Irritable Bowel Syndrome: A Review on the Role of Intestinal Protozoa and the Importance of Their Detection and Diagnosis. Int. J. Parasitol. 2007, 37, 11–20. [Google Scholar] [CrossRef]
- Taşova, Y.; Sahin, B.; Koltaş, S.; Paydaş, S. Clinical Significance and Frequency of Blastocystis Hominis in Turkish Patients with Hematological Malignancy. Acta Med. Okayama 2000, 54, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Yakoob, J.; Jafri, W.; Jafri, N.; Islam, M.; Asim Beg, M. In Vitro Susceptibility of Blastocystis Hominis Isolated from Patients with Irritable Bowel Syndrome. Br. J. Biomed. Sci. 2004, 61, 75–77. [Google Scholar] [CrossRef]
- Mirza, H.; Wu, Z.; Kidwai, F.; Tan, K.S.W. A Metronidazole-Resistant Isolate of Blastocystis Spp. Is Susceptible to Nitric Oxide and Downregulates Intestinal Epithelial Inducible Nitric Oxide Synthase by a Novel Parasite Survival Mechanism. Infect. Immun. 2011, 79, 5019–5026. [Google Scholar] [CrossRef] [Green Version]
- Mirza, H.; Wu, Z.; Teo, J.D.W.; Tan, K.S.W. Statin Pleiotropy Prevents Rho Kinase-Mediated Intestinal Epithelial Barrier Compromise Induced by Blastocystis Cysteine Proteases. Cell Microbiol. 2012, 14, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Puthia, M.K.; Sio, S.W.S.; Lu, J.; Tan, K.S.W. Blastocystis Ratti Induces Contact-Independent Apoptosis, F-Actin Rearrangement, and Barrier Function Disruption in IEC-6 Cells. Infect. Immun. 2006, 74, 4114–4123. [Google Scholar] [CrossRef] [Green Version]
- Puthia, M.K.; Vaithilingam, A.; Lu, J.; Tan, K.S.W. Degradation of Human Secretory Immunoglobulin A by Blastocystis. Parasitol. Res. 2005, 97, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Gentekaki, E.; Curtis, B.; Archibald, J.M.; Roger, A.J. Lateral Gene Transfer in the Adaptation of the Anaerobic Parasite Blastocystis to the Gut. Curr. Biol. 2017, 27, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Gentekaki, E.; Curtis, B.A.; Stairs, C.W.; Klimeš, V.; Eliáš, M.; Salas-Leiva, D.E.; Herman, E.K.; Eme, L.; Arias, M.C.; Henrissat, B.; et al. Extreme Genome Diversity in the Hyper-Prevalent Parasitic Eukaryote Blastocystis. PLOS Biol. 2017, 15, e2003769. [Google Scholar] [CrossRef] [PubMed]
- Yason, J.A.; Liang, Y.R.; Png, C.W.; Zhang, Y.; Tan, K.S.W. Interactions between a Pathogenic Blastocystis Subtype and Gut Microbiota: In Vitro and in Vivo Studies. Microbiome 2019, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audebert, C.; Even, G.; Cian, A.; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M. Colonization with the Enteric Protozoa Blastocystis Is Associated with Increased Diversity of Human Gut Bacterial Microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Nieves-Ramírez, M.E.; Partida-Rodríguez, O.; Laforest-Lapointe, I.; Reynolds, L.A.; Brown, E.M.; Valdez-Salazar, A.; Morán-Silva, P.; Rojas-Velázquez, L.; Morien, E.; Parfrey, L.W.; et al. Asymptomatic Intestinal Colonization with Protist Blastocystis Is Strongly Associated with Distinct Microbiome Ecological Patterns. mSystems 2018, 3, e00007-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yowang, A.; Tsaousis, A.D.; Chumphonsuk, T.; Thongsin, N.; Kullawong, N.; Popluechai, S.; Gentekaki, E. High Diversity of Blastocystis Subtypes Isolated from Asymptomatic Adults Living in Chiang Rai, Thailand. Infect. Genet. Evol. 2018, 65, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Betts, E.L.; Gentekaki, E.; Tsaousis, A.D. Exploring Micro-Eukaryotic Diversity in the Gut: Co-Occurrence of Blastocystis Subtypes and Other Protists in Zoo Animals. Front. Microbiol. 2020, 11, 288. [Google Scholar] [CrossRef]
- Betts, E.L.; Hoque, S.; Torbe, L.; Bailey, J.R.; Ryan, H.; Toller, K.; Breakell, V.; Carpenter, A.I.; Diana, A.; Matechou, E.; et al. Parasites, Drugs and Captivity: Blastocystis-Microbiome Associations in Captive Water Voles. Biology 2021, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.B.; Bonde, I.; Nielsen, H.B.H.B.; Stensvold, C.R. A Retrospective Metagenomics Approach to Studying Blastocystis. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Clark, C.G. Current Status of Blastocystis: A Personal View. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef]
- Billy, V.; Lhotská, Z.; Jirků, M.; Kadlecová, O.; Frgelecová, L.; Parfrey, L.W.; Pomajbíková, K.J. Blastocystis Colonization Alters the Gut Microbiome and, in Some Cases, Promotes Faster Recovery From Induced Colitis. Front Microbiol 2021, 12, 641483. [Google Scholar] [CrossRef]
- Li, W.; Lee, R.E.B.; Lee, R.E.; Li, J. Methods for Acquisition and Assignment of Multidimensional High-Resolution Magic Angle Spinning NMR of Whole Cell Bacteria. Anal. Chem. 2005, 77, 5785–5792. [Google Scholar] [CrossRef] [PubMed]
- Bruno, C.; Patin, F.; Bocca, C.; Nadal-Desbarats, L.; Bonnier, F.; Reynier, P.; Emond, P.; Vourc’h, P.; Joseph-Delafont, K.; Corcia, P.; et al. The Combination of Four Analytical Methods to Explore Skeletal Muscle Metabolomics: Better Coverage of Metabolic Pathways or a Marketing Argument? J. Pharm. Biomed. Anal. 2018, 148, 273–279. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Turbitt, J.R.; Colson, K.L.; Killday, K.B.; Milstead, A.; Neto, C.C. Application of 1H-NMR-Based Metabolomics to the Analysis of Cranberry (Vaccinium Macrocarpon) Supplements. Phytochem. Anal. 2019, 31, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Viana, T.; Petersen, M.A.; Larsen, F.H.; Arneborg, N. Metabolic Footprint Analysis of Metabolites That Discriminate Single and Mixed Yeast Cultures at Two Key Time-Points during Mixed Culture Alcoholic Fermentations. Metabolomics 2018, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.N.; Panagos, C.G.; Mosedale, W.R.T.; Kváč, M.; Howard, M.J.; Tsaousis, A.D. NMR Metabolomics Reveals Effects of Cryptosporidium Infections on Host Cell Metabolome. Gut Pathog. 2019, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermathen, M.; Müller, J.; Furrer, J.; Müller, N.; Vermathen, P. 1H HR-MAS NMR Spectroscopy to Study the Metabolome of the Protozoan Parasite Giardia Lamblia. Talanta 2018, 188, 429–441. [Google Scholar] [CrossRef]
- Newton, J.M.; Betts, E.L.; Yiangou, L.; Ortega Roldan, J.; Tsaousis, A.D.; Thompson, G.S. Establishing a Metabolite Extraction Method to Study the Metabolome of Blastocystis Using NMR. Molecules 2021, 26, 3285. [Google Scholar] [CrossRef]
- Bond, A.; Vernon, A.; Reade, S.; Mayor, A.; Wastling, J.; Minetti, C.; Lamden, K.; Probert, C. PWE-173 Investigation of Volatile Organic Compounds Emitted from Faeces for the Diagnosis of Giardiasis. Gut 2015, 64, A288. [Google Scholar] [CrossRef]
- Hublin, J.S.Y.; Ryan, U.; Trengove, R.D.; Maker, G.L. Development of an Untargeted Metabolomics Method for the Analysis of Human Faecal Samples Using Cryptosporidium-Infected Samples. Mol. Biochem. Parasitol. 2012, 185, 145–150. [Google Scholar] [CrossRef]
- Hublin, J.S.Y.N.; Ryan, U.; Trengove, R.; Maker, G. Metabolomic Profiling of Faecal Extracts from Cryptosporidium parvum Infection in Experimental Mouse Models. PLoS ONE 2013, 8, e77803. [Google Scholar] [CrossRef]
- Allman, E.L.; Painter, H.J.; Samra, J.; Carrasquilla, M.; Llinás, M. Metabolomic Profiling of the Malaria Box Reveals Antimalarial Target Pathways. Antimicrob Agents Chemother 2016, 60, 6635–6649. [Google Scholar] [CrossRef] [Green Version]
- Jeelani, G.; Sato, D.; Husain, A.; Cadiz, A.E.; Sugimoto, M.; Soga, T.; Suematsu, M.; Nozaki, T. Metabolic Profiling of the Protozoan Parasite Entamoeba Invadens Revealed Activation of Unpredicted Pathway during Encystation. PLoS ONE 2012, 7, e37740. [Google Scholar] [CrossRef] [Green Version]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient Metabolism by the Human Gut Microbiome: Major Fermentation by-Products and Their Impact on Host Health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Yang, Z.-H.; Miyahara, H.; Takeo, J.; Katayama, M. Diet High in Fat and Sucrose Induces Rapid Onset of Obesity-Related Metabolic Syndrome Partly through Rapid Response of Genes Involved in Lipogenesis, Insulin Signalling and Inflammation in Mice. Diabetol. Metab. Syndr. 2012, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Poirier, P.; Wawrzyniak, I.; Vivarès, C.P.; Delbac, F.; El Alaoui, H. New Insights into Blastocystis Spp.: A Potential Link with Irritable Bowel Syndrome. PLoS Pathog. 2012, 8, e1002545. [Google Scholar] [CrossRef] [Green Version]
- Lennernäs, H. Intestinal Permeability and Its Relevance for Absorption and Elimination. Xenobiotica 2007, 37, 1015–1051. [Google Scholar] [CrossRef]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Brantley Hall, A.; Mallick, H.; McIver, L.J.; et al. Gut Microbiome Structure and Metabolic Activity in Inflammatory Bowel Disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut Microbiota-Derived Metabolites as Key Actors in Inflammatory Bowel Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Holmes, E.; Khan, F.; Kochhar, S.; Scanlan, P.; Shanahan, F.; Wilson, I.D.; Wang, Y. Rapid and Noninvasive Metabonomic Characterization of Inflammatory Bowel Disease. J. Proteome Res. 2007, 6, 546–551. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Roediger, W.E. Utilization of Nutrients by Isolated Epithelial Cells of the Rat Colon. Gastroenterology 1982, 83, 424–429. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2019, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Smolinska, S.; Jutel, M.; Crameri, R.; O’Mahony, L. Histamine and Gut Mucosal Immune Regulation. Allergy 2014, 69, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.; Villaret-Cazadamont, J.; Claus, S.P.; Canlet, C.; Guillou, H.; Cabaton, N.J.; Ellero-Simatos, S. Important Considerations for Sample Collection in Metabolomics Studies with a Special Focus on Applications to Liver Functions. Metabolites 2020, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Bosch, S.; Struys, E.A.; van Gaal, N.; Bakkali, A.; Jansen, E.W.; Diederen, K.; Benninga, M.A.; Mulder, C.J.; de Boer, N.K.H.; de Meij, T.G.J. Fecal Amino Acid Analysis Can Discriminate De Novo Treatment-Naïve Pediatric Inflammatory Bowel Disease From Controls. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 773–778. [Google Scholar] [CrossRef]
- Barcik, W.; Wawrzyniak, M.; Akdis, C.A.; O’Mahony, L. Immune Regulation by Histamine and Histamine-Secreting Bacteria. Curr. Opin. Immunol. 2017, 48, 108–113. [Google Scholar] [CrossRef]
- Fabisiak, A.; Włodarczyk, J.; Fabisiak, N.; Storr, M.; Fichna, J. Targeting Histamine Receptors in Irritable Bowel Syndrome: A Critical Appraisal. J. Neurogastroenterol. Motil. 2017, 23, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.X.; Png, C.W.; Tay, C.Y.B.; Teo, J.D.W.; Jiao, H.; Lehming, N.; Tan, K.S.g.W.; Zhang, Y. Differential regulation of proinflammatory cytokine expression by mitogen-activated protein kinases in macrophages in response to intestinal parasite infection. Infect. Immun. 2014, 82, 4789–4801. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Yoshikawa, H.; Yamada, M.; Kimata, I.; Arizono, N. Expression of interferon gamma and proinflammatory cytokines in the cecal mucosa of rats experimentally infected with Blastocystis sp. strain RN94-9. Parasitol. Res. 2005, 105, 135–140. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Won, E.J.; Kim, S.H.; Shin, J.H.; Chai, J.-Y. Molecular Detection and Subtyping of Human Blastocystis and the Clinical Implications: Comparisons between Diarrheal and Non-Diarrheal Groups in Korean Populations. Korean J. Parasitol. 2020, 58, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and Evaluation of a Real-Time PCR Assay for Detection and Quantification of Blastocystis Parasites in Human Stool Samples: Prospective Study of Patients with Hematological Malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Ahmed, U.N.; Andersen, L.O.; Nielsen, H.V. Development and evaluation of a genus-specific, probe-based, internal-process-controlled real-time PCR assay for sensitive and specific detection of Blastocystis spp. J. Clin. Microbiol. 2012, 50, 1847–1851. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Giskeødegård, G.F.; Bathen, T.F.; Euceda, L.R. Preprocessing of NMR Metabolomics Data. Scand. J. Clin. Lab. Investig. 2015, 75, 193–203. [Google Scholar]
- Xia, J.; Wishart, D.S. MetPA: A WeB−Based Metabolomics Tool for Pathway Analysis and Visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [Green Version]
- Goeman, J.J.; van de Geer, S.A.; de Kort, F.; van Houwelingen, H.C. A Global Test for Groups of Genes: Testing Association with a Clinical Outcome. Bioinformatics 2004, 20, 93–99. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Notation | No. Groups | No. Samples |
|---|---|---|---|
| Thailand | B+/B− | 2 | 44 |
| Turkey | B+/B− | 2 | 69 |
| South Korea | B+/B− | 2 | 38 |
| Positive vs. Negative | B+/B− | 2 | 151 |
| Asymptomatic Blastocystis positive vs. Symptomatic, Blastocystis negative | D−B+/D+B− | 2 | 98 |
| Asymptomatic, Blastocystis positive vs. Asymptomatic, Blastocystis negative | D−B+/D−B− | 2 | 62 |
| Blastocystis positive vs. Blastocystis negative (excluding Blastocystis negative from Korea) | B+/B− | 2 | 131 |
| Country and Blastocystis status | Country B+/B− | 6 | 151 |
| Country | Turkey/Thailand/S. Korea | 3 | 151 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betts, E.L.; Newton, J.M.; Thompson, G.S.; Sarzhanov, F.; Jinatham, V.; Kim, M.-J.; Popluechai, S.; Dogruman-Al, F.; Won, E.-J.; Gentekaki, E.; et al. Metabolic Fluctuations in the Human Stool Obtained from Blastocystis Carriers and Non-Carriers. Metabolites 2021, 11, 883. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120883
Betts EL, Newton JM, Thompson GS, Sarzhanov F, Jinatham V, Kim M-J, Popluechai S, Dogruman-Al F, Won E-J, Gentekaki E, et al. Metabolic Fluctuations in the Human Stool Obtained from Blastocystis Carriers and Non-Carriers. Metabolites. 2021; 11(12):883. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120883
Chicago/Turabian StyleBetts, Emma L., Jamie M. Newton, Gary S. Thompson, Fakhriddin Sarzhanov, Vasana Jinatham, Moon-Ju Kim, Siam Popluechai, Funda Dogruman-Al, Eun-Jeong Won, Eleni Gentekaki, and et al. 2021. "Metabolic Fluctuations in the Human Stool Obtained from Blastocystis Carriers and Non-Carriers" Metabolites 11, no. 12: 883. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120883