
Effect of the Length of Oat Hay on Growth Performance, Health Status, Behavior Parameters and Rumen Fermentation of Holstein Female Calves

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
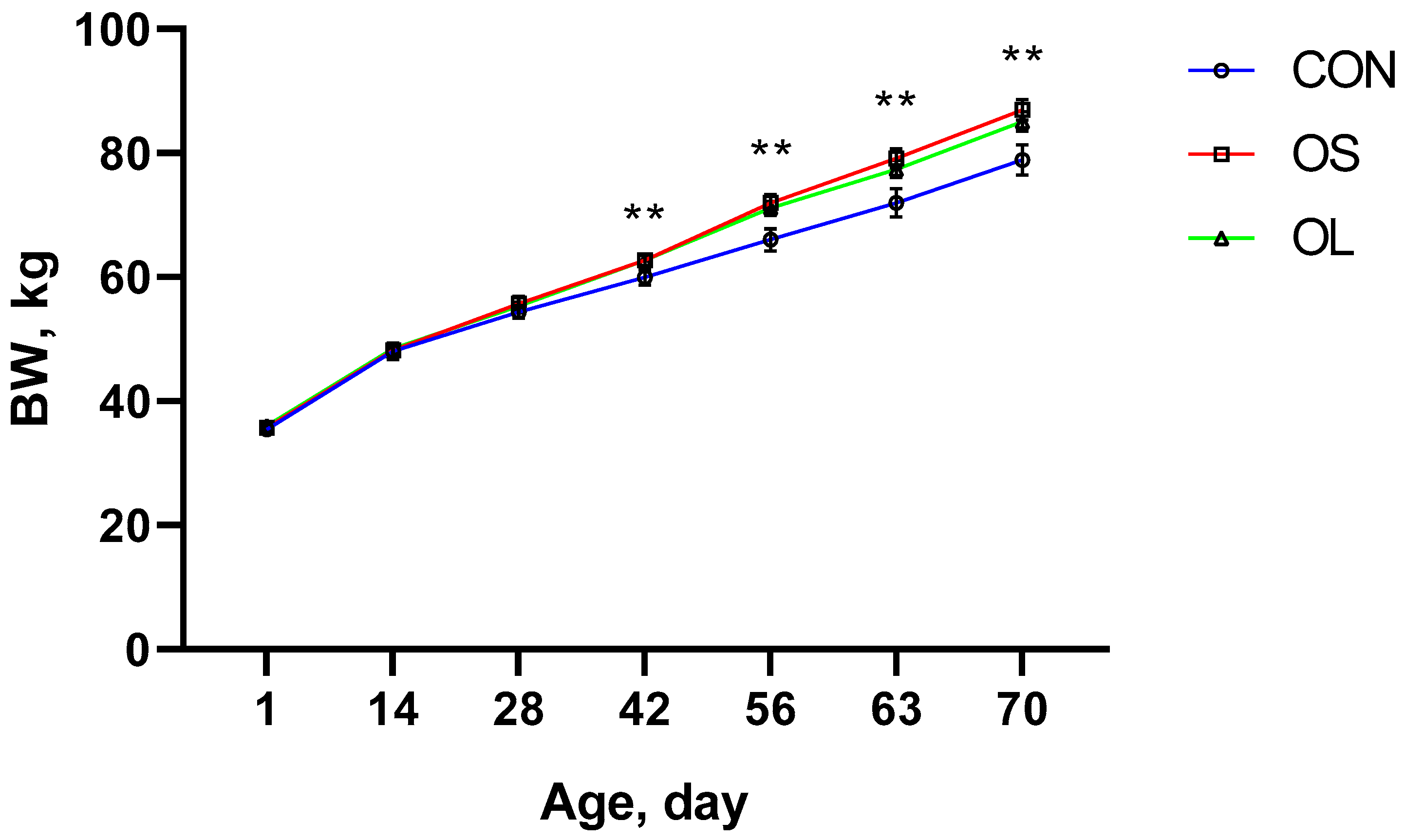
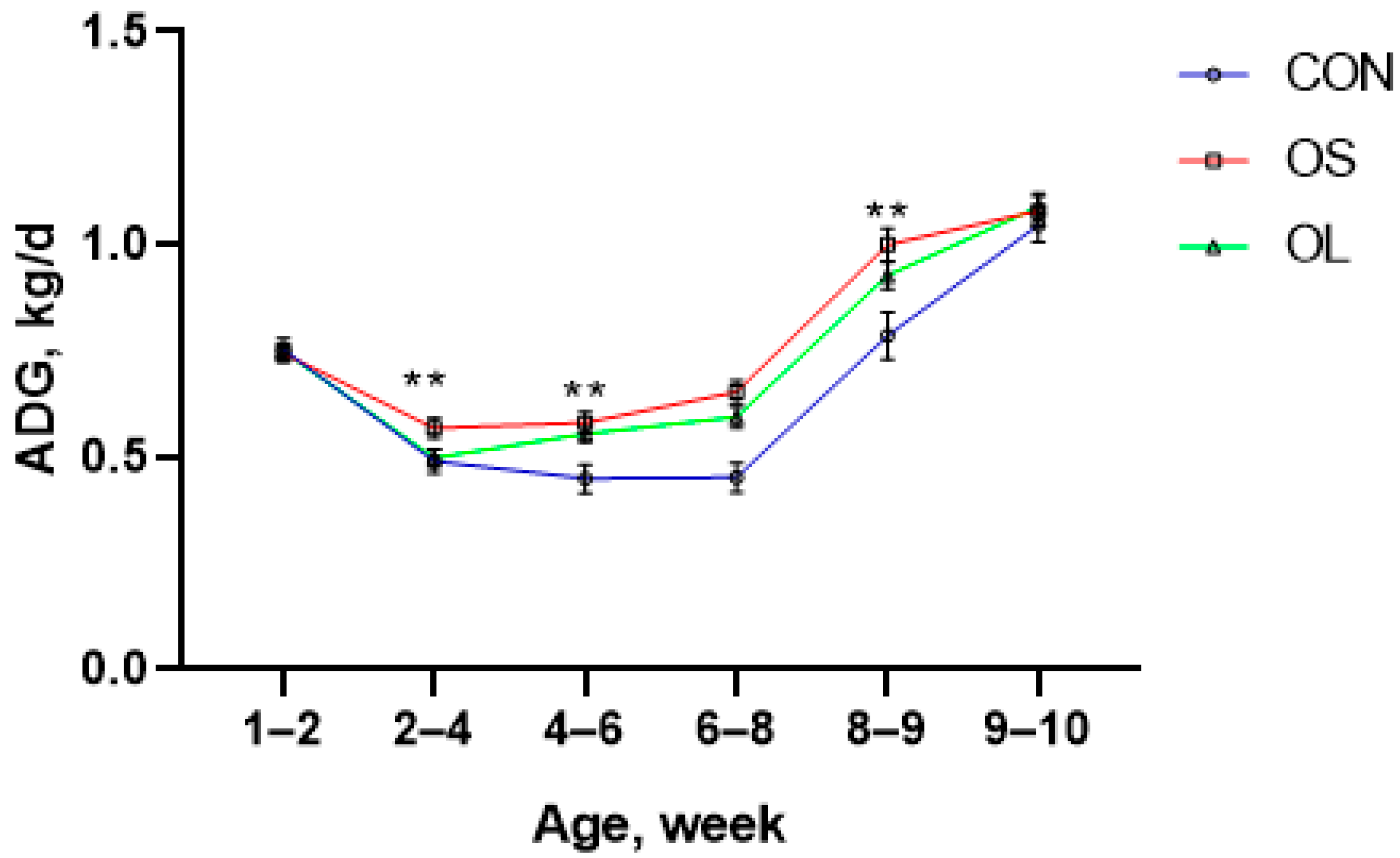
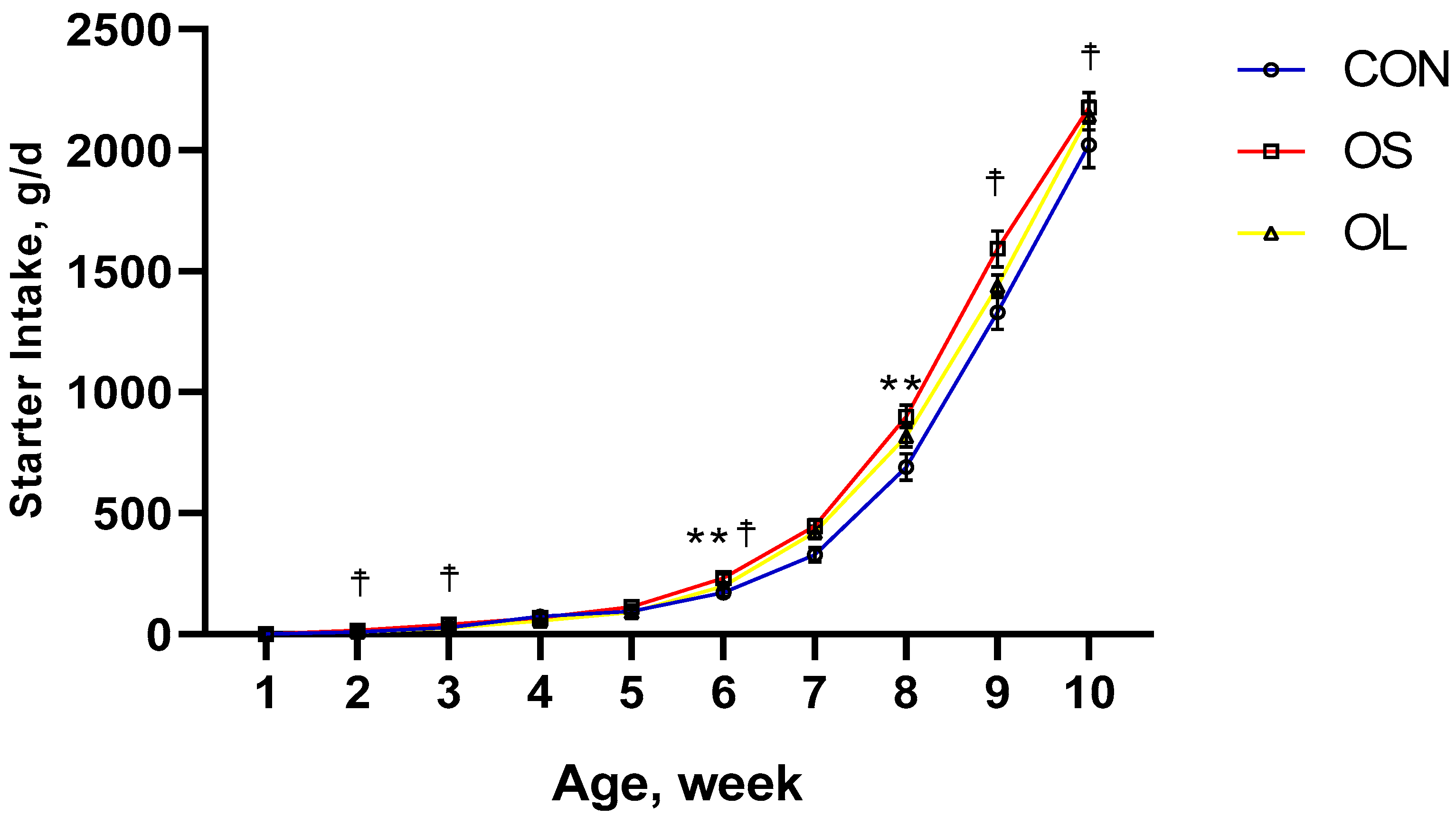
2.1. BW, ADG and Starter Intake
2.2. Body Growth Parameters
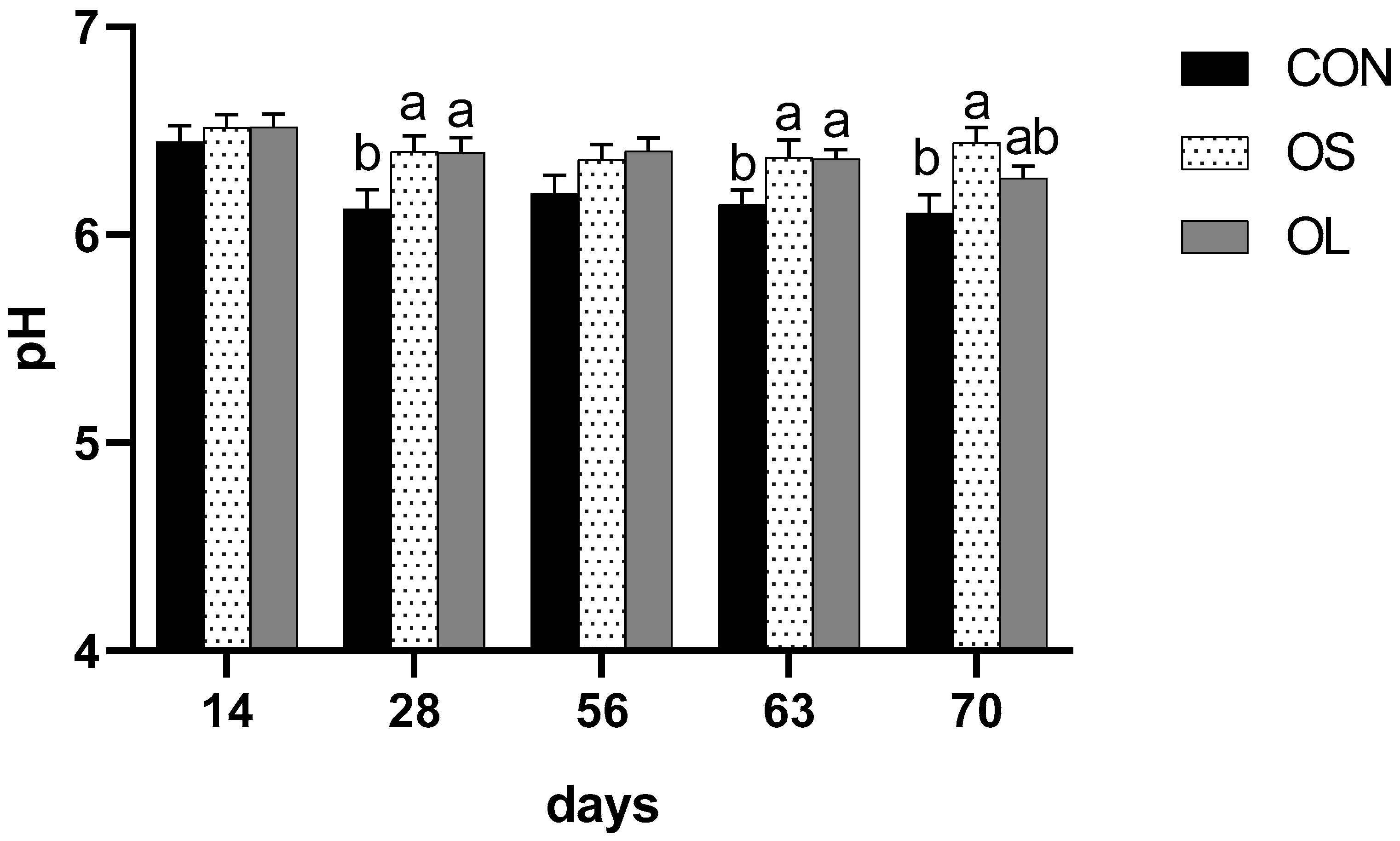
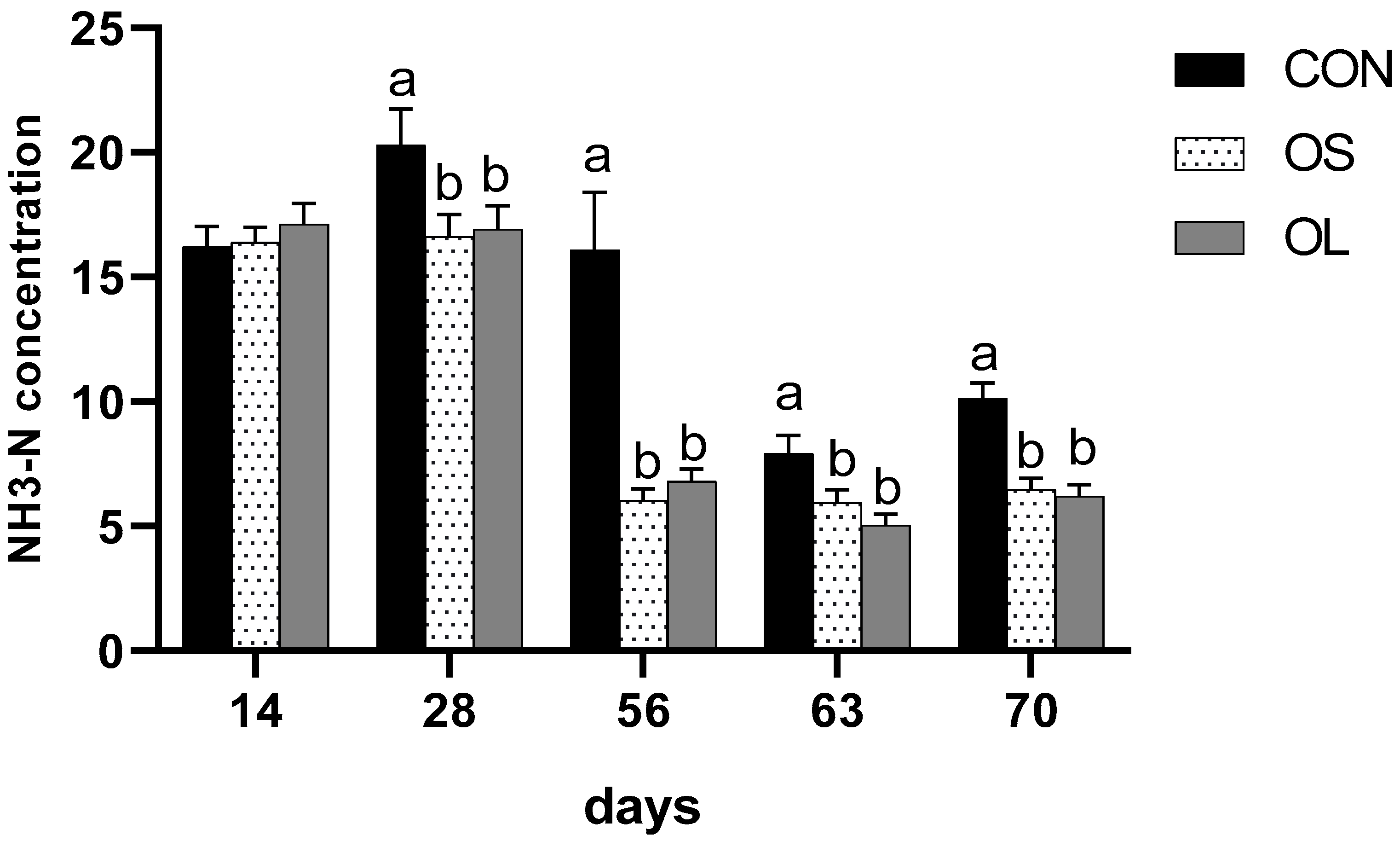
2.3. pH and NH3-N
2.4. Rumen Volatile Fatty Acids
2.5. Calf Health
2.6. Calf Behavior
3. Discussion
3.1. Calf Growth Performance
3.2. Rumen Fermentation
3.3. Calf Health
3.4. Calf Behavior
4. Materials and Methods
4.1. Ethical Statement
4.2. Samples Selection and Treatments
4.3. Feed Analysis and Body Measurements
4.4. Collection and Determination of the Rumen Fluid Samples
4.5. Evaluation of Calf Health Status
4.6. Calf Behavior
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Liu, Q.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L.; Guo, G.; Huo, W.J.; Yang, W.Z.; Wang, H. Effects of isobutyrate supplementation in pre- and post-weaned dairy calves diet on growth performance, rumen development, blood metabolites and hormone secretion. Animal 2017, 11, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Ørskov, E.R.; Benzie, D.; Kay, R.N.B. The effects of feeding procedure on closure of the oesophageal groove in young sheep. Br. J. Nutr. 1970, 24, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Schäff, C.; Gruse, J.; Maciej, J.; Pfuhl, R.; Zitnan, R.; Rajsky, M.; Hammon, H. Effects of feeding unlimited amounts of milk replacer for the first 5 weeks of age on rumen and small intestinal growth and development in dairy calves. J. Dairy Sci. 2018, 101, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, R.L., VI; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen development, intestinal growth and hepatic metabolism in the pre- and postweaning ruminant. J. Dairy Sci. 2004, 87, E55–E65. [Google Scholar] [CrossRef] [Green Version]
- Laarman, A.H.; Oba, M. Short communication: Effect of calf starter on rumen pH of Holstein dairy calves at weaning. J. Dairy Sci. 2011, 94, 5661–5664. [Google Scholar] [CrossRef]
- Van Ackeren, C.; Steingass, H.; Hartung, K.; Funk, R.; Drochner, W. Effect of roughage level in a total mixed ration on feed intake, ruminal fermentation patterns and chewing activity of early-weaned calves with ad libitum access to grass hay. Anim. Feed Sci. Technol. 2009, 153, 48–59. [Google Scholar] [CrossRef]
- Beharka, A.A.; Nagaraja, T.G.; Morrill, J.L.; Kennedy, G.A.; Klemm, R.D. Effects of Form of the Diet on Anatomical, Microbial, and Fermentative Development of the Rumen of Neonatal Calves. J. Dairy Sci. 1998, 81, 1946–1955. [Google Scholar] [CrossRef]
- Mirzaei, M.; Khorvash, M.; Ghorbani, G.R.; Kazemi-Bonchenari, M.; Riasi, A.; Nabipour, A.; van den Borne, J. Effects of supplementation level and particle size of alfalfa hay on growth characteristics and rumen development in dairy calves. J. Anim. Physiol. Anim. Nutr. 2015, 99, 553–564. [Google Scholar] [CrossRef]
- Norouzian, M.A.; Valizadeh, R. Effect of forage inclusion and particle size in diets of neonatal lambs on performance and rumen development. J. Anim. Physiol. Anim. Nutr. 2014, 98, 1095–1101. [Google Scholar] [CrossRef]
- Castells, L.; Bach, A.; Aris, A.; Terre, M. Effects of Forage Provision to Young Calves on Rumen Fermentation and Development of the Gastrointestinal Tract. J. Dairy Sci. 2013, 96, 5226–5236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoro, C.; Miller-Cushon, E.K.; DeVries, T.J.; Bach, A. Effect of physical form of forage on performance, feeding behavior, and digestibility of Holstein calves. J. Dairy Sci. 2013, 96, 1117–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemati, M.; Amanlou, H.; Khorvash, M.; Moshiri, B.; Mirzaei, M.; Khan, M.A.; Ghaffari, M.H. Rumen fermentation, blood metabolites, and growth performance of calves during transition from liquid to solid feed: Effects of dietary level and particle size of alfalfa hay. J. Dairy Sci. 2015, 98, 7131–7141. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K. Calf Nutrition from Birth to Breeding. Vet. Clin. N. Am. Food Anim. 2008, 24, 55–86. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle; NRC: Rockville, MD, USA, 2001. [Google Scholar]
- Allen, M.S. Relationship between fermentation acid production in the rumen and the requirement for physically effective fiber. J. Dairy Sci. 1997, 80, 1447–1462. [Google Scholar] [CrossRef]
- Hill, T.M.; Bateman, H.G.; Aldrich, J.M.; Schlotterbeck, R.L. Effects of the amount of chopped hay or cottonseed hulls in a textured calf starter on young calf performance. J. Dairy Sci. 2008, 91, 2684–2693. [Google Scholar] [CrossRef] [Green Version]
- Castells, L.I.; Bach, A.; Terre, M. Effect of Different Forage Sources on Performance and Feeding Behaviour of Holstein Calves. J. Dairy Sci. 2012, 95, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Kohari, D.; Namikawa, S.; Tanaka, M. How do visual and auditory cues influence calf stress reactions at weaning? J. Anim Sci. 2014, 85, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M.; Fraser, A.F. Domestic Animal Behaviour and Welfare; CABI: Wallingford, UK, 2015. [Google Scholar]
- Phillips, C.J. The effects of forage provision and group size on the behavior of calves. J. Dairy Sci. 2004, 87, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Suarez Hill, T.M.; Jones, C.M.; Heinrichs, A.J. Review: Effect of forage provision on feed intake in dairy calves. Prof. Anim. Sci. 2016, 32, 383–388. [Google Scholar] [CrossRef]
- Beiranvand, H.; Ghorbani, G.R.; Khorvash, M.; Nabipour, A.; Dehghan-Banadaky, M.; Homayouni, A.; Kargar, S. Interactions of Alfalfa Hay and Sodium Propionate on Dairy Calf Performance and Rumen Development. J. Dairy Sci. 2014, 97, 2270–2280. [Google Scholar] [CrossRef]
- Nemati, M.; Amanlou, H.; Khorvash, M.; Mirzaei, M.; Moshiri, B.; Ghaffari, M.H. Effect of different alfalfa hay levels on growth performance, rumen fermentation, and structural growth of Holstein dairy calves. J. Anim. Sci. 2016, 94, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Weary, D.M.; Von, M.A. Hay Intake Improves Performance and Rumen Development of Calves Fed Higher Quantities of Milk. J. Dairy Sci. 2011, 94, 3547–3553. [Google Scholar] [CrossRef] [Green Version]
- Tamate, H.; Mcgilliard, A.D.; Jacobson, N.L.; Getty, R. Effect of Various Dietaries on the Anatomical Development of the Stomach in the Calf1. J. Dairy Sci. 1962, 45, 408–420. [Google Scholar] [CrossRef]
- Coverdale, J.A.; Tyler, H.D.; Quigley, J.D.; Brumm, J.A. Effect of Various Levels of Forage and Form of Diet on Rumen Development and Growth in Calves. J. Dairy Sci. 2004, 87, 2554–2562. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, L.K.; Heinrichs, A.J. Increasing grass hay inclusion level on weaned dairy calf growth, intake, digestibility, and ruminal fermentation. J. Dairy Sci. 2020, 103, 9012–9023. [Google Scholar] [CrossRef]
- Zitnan, R.; Voigt, J.; Schönhusen, U.; Wegner, J.; Kokardová, M.; Hagemeister, H.; Levkut, M.; Kuhla, S.; Sommer, A. Influence of dietary concentrate to forage ratio on the development of rumen mucosa in calves. Archiv fur Tierernahrung 1998, 51, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.M.; Ghorbani, G.R.; Rezamand, P.; Khorvash, M. Determining Optimum Age of Holstein Dairy Calves when Adding Chopped Alfalfa Hay to Meal Starter Diets Based on Measures of Growth and Performance. J. Anim. Biosci. 2015, 10, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasiorek, M.; Stefanska, B.; Pruszynska-Oszmalek, E.; Taciak, M.; Komisarek, J.; Nowak, W. Effect of oat hay provision method on growth performance, rumen fermentation and blood metabolites of dairy calves during preweaning and postweaning periods. Animal 2020, 14, 2054–2062. [Google Scholar] [CrossRef] [PubMed]
- Suárez, B.J.; Van Reenen, C.G.; Stockhofe, N.; Dijkstra, J.; Gerrits, W.J.J. Effect of Roughage Source and Roughage to Concentrate Ratio on Animal Performance and Rumen Development in Veal Calves. J. Dairy Sci. 2007, 90, 2390–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Mena, F.X.; Heinrichs, A.J.; Jones, C.M.; Hill, T.M.; Quigley, J.D. Straw particle size in calf starters: Effects on digestive system development and rumen fermentation. J. Dairy Sci. 2016, 99, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Terre, M.; Pedrals, E.; Dalmau, A.; Bach, A. What do preweaned and weaned calves need in the diet: A high fiber content or a forage source? J. Dairy Sci. 2013, 96, 5217–5225. [Google Scholar] [CrossRef] [Green Version]
- Teimouri Yansari, A.; Valizadeh, R.; Naserian, A.; Christensen, D.A.; Yu, P.; Eftekhari Shahroodi, F. Effects of alfalfa particle size and specific gravity on chewing activity, digestibility, and performance of Holstein dairy cows. J. Dairy Sci. 2004, 87, 3912–3924. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.X.; Alugongo, G.M.; Li, J.H.; Wang, Y.J.; Li, S.L.; Cao, Z.J. Review: How Forage Feeding Early in Life Influences the Growth Rate, Ruminal Environment, and the Establishment of Feeding Behavior in Pre-Weaned Calves. Animals 2020, 10, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanile, G.; Di Palo, R.; Infascelli, F.; Gasparrini, B.; Neglia, G.; Zicarelli, F.; D’Occhio, M.J. Influence of rumen protein degradability on productive and reproductive performance in buffalo cows. Reprod. Nutr. Dev. 2003, 43, 557–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, G.R.; Morgavi, D.P.; Beauchemin, K.A.; Leedle, J.A.Z. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables, and the microbial populations of feedlot cattle. J. Anim. Sci. 2002, 80, 1977–1985. [Google Scholar] [CrossRef] [Green Version]
- Firkins, J.L.; Yu, Z.; Morrison, M. Ruminal nitrogen metabolism: Perspectives for integration of microbiology and nutrition for dairy. J. Dairy Sci. 2007, 90 (Suppl. 1), E1–E16. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the Bovine Rumen Bacterial Community from Birth to Adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257. [Google Scholar] [CrossRef]
- Belanche, A.; Doreau, M.; Edwards, J.E.; Moorby, J.M.; Pinloche, E.; Newbold, C.J. Shifts in the Rumen Microbiota Due to the Type of Carbohydrate and Level of Protein Ingested by Dairy Cattle Are Associated with Changes in Rumen Fermentation. J. Nutr. 2012, 142, 1684–1692. [Google Scholar] [CrossRef]
- Lin, X.Y.; Wang, J.; Hou, Q.L.; Wang, Y.; Hu, Z.Y.; Shi, K.R.; Yan, Z.G.; Wang, Z.H. Effect of hay supplementation timing on rumen microbiota in suckling calves. Microbiol. Open. 2018, 7, 8. [Google Scholar] [CrossRef]
- Karimizadeh, E.; Chaji, M.; Mohammadabadi, T. Effects of physical form of diet on nutrient digestibility, rumen fermentation, rumination, growth performance and protozoa population of finishing lambs. Anim. Nutr. 2017, 3, 139–144. [Google Scholar] [CrossRef]
- Kiran, D.; Mutsvangwa, T. Effects of barley grain processing and dietary ruminally degradable protein on urea nitrogen recycling and nitrogen metabolism in growing lambs. J. Anim Sci. 2007, 85, 3391–3399. [Google Scholar] [CrossRef]
- Blottiere, H.M.; Buecher, B.; Galmiche, J.P.; Cherbut, C. Molecular Analysis of the Effect of Short-chain Fatty Acids on Intestinal Cell Proliferation. Proc. Nutr. Soc. 2003, 62, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwińska, B.; Hanczakowska, E.; Arciszewski, M.B.; Klebaniuk, R. Review: Exogenous butyrate: Implications for the functional development of ruminal epithelium and calf performance. Animal 2017, 11, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calsamiglia, S.; Cardozo, P.W.; Ferret, A.; Bach, A. Changes in Rumen Microbial Fermentation Are Due to a Combined Effect of Type of Diet and pH. J. Anim. Sci. 2008, 86, 702–711. [Google Scholar] [CrossRef]
- Porter, J.C.; Warner, R.G.; Kertz, A.F. Effect of Fiber Level and Physical Form of Starter on Growth and Development of Dairy Calves Fed No Forage. J. Prof. Anim. Sci. 2007, 23, 395–400. [Google Scholar] [CrossRef]
- Ridpath, J.E.; Neill, J.D.; Endsley, J.; Roth, J.A. Effect of passive immunity on the development of a protective immune response against bovine viral diarrhea virus in calves. Am. J. Vet. Res. 2003, 64, 65–69. [Google Scholar] [CrossRef]
- Bringhenti, L.; Pallu, M.; Silva, J.; Tomazi, T.; Tomazi, A.C.; Rodrigues, M.X.; Duarte, L.M.; Bilby, T.R.; Bicalho, R.C. Effect of metaphylactic administration of tildipirosin on the incidence of pneumonia and otitis and on the upper respiratory tract and fecal microbiome of preweaning Holstein calves. J. Dairy Sci. 2021, 104, 6020–6038. [Google Scholar] [CrossRef]
- Callan, R.J.; Garry, F.B. Biosecurity and bovine respiratory disease. Vet. Clin. N. Am. Food Anim. Pract. 2002, 18, 57–77. [Google Scholar] [CrossRef]
- Maier, G.U.; Love, W.J.; Karle, B.M.; Dubrovsky, S.A.; Williams, D.R.; Champagne, J.D.; Anderson, R.J.; Rowe, J.D.; Lehenbauer, T.W.; Van Eenennaam, A.L.; et al. Management factors associated with bovine respiratory disease in preweaned calves on California dairies: The BRD 100 study. J. Dairy Sci. 2019, 102, 7288–7305. [Google Scholar] [CrossRef]
- Brown, A.; Bivort, B. Ethology As a Physical Science. Nat. Phys. 2018, 14, 653–657. [Google Scholar] [CrossRef]
- Herd, R.M.; Arthur, P.F. Physiological basis for residual feed intake. J. Anim. Sci. 2009, 87, E64–E71. [Google Scholar] [CrossRef]
- Richardson, E.C.; Kilgour, R.J.; Archer, J.A.; Herd, R.M. Pedometers measure differences in activity in bulls selected for high or low net feed efficiency. Proc. Aust. Soc. Study Anim. Behav. 1999, 26, 16. [Google Scholar]
- Allen, M.S. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci. 2000, 83, 1598–1624. [Google Scholar] [CrossRef]
- Horvath, K.C.; Miller-Cushon, E.K. Evaluating effects of providing hay on behavioral development and performance of group-housed dairy calves. J. Dairy Sci. 2019, 102, 10411–10422. [Google Scholar] [CrossRef] [Green Version]
- Flower, F.C.; Weary, D.M. Effects of early separation on the dairy cow and calf: 2. Separation at 1 day and 2 weeks after birth. Appl. Anim. Behav. Sci. 2001, 70, 275–284. [Google Scholar] [CrossRef]
- Randhawa, S.S.; Singh, J.; Misra, S.K. An experimental study of acid-base status of buffalo-calves in rumen acidosis. Zentralblatt fur Veterinarmedizin Reihe A 1980, 27, 255–258. [Google Scholar] [CrossRef]
- Chen, T.; Xiao, J.; Li, T.; Ma, J.; Alugongo, G.M.; Khan, M.Z.; Liu, S.; Wang, W.; Wang, Y.; Li, S. Effect of the Initial Time of Providing Oat Hay on Performance, Health, Behavior and Rumen Fermentation in Holstein Female Calves. Agriculture 2021, 11, 862. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis of AOAC International, 16th edition. Volume 2. Trends Food Sci. Technol. 1995, 6, 382. [Google Scholar] [CrossRef]
- Erwin, E.S.; Marco, G.J.; Emery, E.M. Volatile Fatty Acid Analyses of Blood and Rumem Fluid by Gas Chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Zhang, N.F.; Diao, Q.Y.; Hui, L.I. Effect of Different Contents of Plant Protein on the Incidence of Diarrhea and Blood Parameters in Calves Aged 6-11 Days. Sci. Agric. Sin. 2010, 43, 4094–4100. [Google Scholar]
- Love, W.J.; Lehenbauer, T.W.; Kass, P.H.; Van Eenennaam, A.L.; Aly, S.S. Development of a novel clinical scoring system for on-farm diagnosis of bovine respiratory disease in pre-weaned dairy calves. PeerJ 2014, 2, e238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnoldmeeks, C.; McGlone, J.J. Validating Techniques to Sample Behavior of Confined, Young-pigs. Appl. Anim. Behav. Sci. 1986, 16, 149–155. [Google Scholar] [CrossRef]
- Mitlohner, F.M.; Morrow-Tesch, J.L.; Wilson, S.C.; Dailey, J.W.; McGlone, J.J. Behavioral sampling techniques for feedlot cattle. J. Anim. Sci. 2001, 79, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, E.M.; Schutz, M.M.; Lay, D.C., Jr.; Marchant-Forde, J.N.; Eicher, S.D. Effect of group size on behavior, health, production, and welfare of veal calves. J. Anim. Sci. 2013, 91, 5455–5465. [Google Scholar] [CrossRef] [PubMed]
- Loberg, J.M.; Hernandez, C.E.; Thierfelder, T.; Jensen, M.B.; Berg, C.; Lidfors, L. Reaction of foster cows to prevention of suckling from and separation from four calves simultaneously or in two steps. J. Anim. Sci. 2007, 85, 1522–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kargar, S.; Kanani, M. Reconstituted Versus Dry Alfalfa Hay in Starter Feed Diets of Holstein Dairy Calves: Effects on Feed Intake, Feeding and Chewing Behavior, Feed Preference, and Health Criteria. J. Dairy Sci. 2019, 102, 4061–4071. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Treatment 2 | SEM | Contrast p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | OS | OL | T | t | T * t | p 4 | T * p | ||
| Initial BW (kg) | 35.43 | 35.70 | 36.01 | - | 0.46 | - | - | - | - |
| BW (kg) | |||||||||
| Pre-weaning | 52.76 b | 54.87 a | 54.69 a | 0.53 | <0.01 | < 0.01 | < 0.01 | - | - |
| Post-weaning | 75.41 b | 83.05 a | 81.24 a | 0.90 | <0.01 | < 0.01 | 0.42 | - | - |
| Entire trial | 64.09 b | 68.92 a | 67.96 a | 0.78 | <0.01 | - | - | <0.01 | <0.01 |
| ADG (kg/d) | |||||||||
| Pre-weaning | 0.54 c | 0.64 a | 0.60 b | 0.03 | < 0.01 | <0.01 | < 0.01 | - | - |
| Post-weaning | 0.92 b | 1.04 a | 1.01 a | 0.04 | < 0.01 | <0.01 | 0.17 | - | - |
| Entire trial | 0.72 c | 0.84 a | 0.80 b | 0.014 | < 0.01 | - | - | <0.01 | 0.87 |
| Calf starter intake 3 (g/d) | |||||||||
| Pre-weaning | 174.65 b | 226.83 a | 202.63 ab | 11.87 | <0.05 | <0.01 | <0.01 | - | - |
| Post-weaning | 1675.97 | 1883.45 | 1790.92 | 62.28 | 0.10 | <0.01 | 0.17 | - | - |
| Entire trial | 925.31 b | 1055.14 a | 996.78 ab | 35.71 | <0.05 | - | - | <0.01 | 0.18 |
| Treatment 2 | SEM | Contrast p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Items | CON | OS | OL | T | t | T * t | p 3 | T * p | |
| Body height (cm) | |||||||||
| Pre-weaning | 80.36 | 80.20 | 80.45 | 0.30 | 0.72 | <0.01 | 0.19 | - | - |
| Post-weaning | 88.08 | 88.49 | 88.42 | 0.31 | 0.66 | <0.01 | 0.53 | - | - |
| Entire trial | 84.22 | 84.34 | 84.43 | 0.30 | 0.84 | - | - | <0.01 | 0.23 |
| Body length (cm) | |||||||||
| Pre-weaning | 75.04 | 75.19 | 75.46 | 0.22 | 0.37 | <0.01 | 0.09 | - | - |
| Post-weaning | 84.24 b | 85.94 a | 86.00 a | 0.30 | <0.01 | <0.01 | 0.84 | - | -- |
| Entire trial | 79.64 b | 80.56 a | 80.73 a | 0.22 | <0.05 | - | - | <0.01 | <0.01 |
| Heart girth (cm) | |||||||||
| Pre-weaning | 86.49 | 86.97 | 87.05 | 0.32 | 0.35 | <0.01 | <0.05 | - | - |
| Post-weaning | 100.39 b | 101.92 a | 101.53 a | 0.39 | <0.05 | <0.01 | 0.97 | - | - |
| Entire trial | 93.44 b | 94.43 a | 94.29 a | 0.27 | 0.08 | - | - | <0.01 | <0.05 |
| Abdominal Girth (cm) | |||||||||
| Pre-weaning | 92.48 b | 94.21 a | 94.20 a | 0.38 | <0.01 | <0.01 | 0.07 | - | - |
| Post-weaning | 114.35 b | 118.50 a | 118.14 a | 0.60 | <0.01 | <0.01 | 0.53 | - | - |
| Entire trial | 103.42 b | 106.36 a | 106.17 a | 0.44 | <0.01 | - | - | <0.01 | <0.01 |
| Circumference of cannon bone (cm) | |||||||||
| Pre-weaning | 10.74 | 10.80 | 10.81 | 0.04 | 0.39 | <0.01 | 0.24 | - | - |
| Post-weaning | 11.47 | 11.60 | 11.59 | 0.05 | 0.20 | <0.01 | 0.90 | - | - |
| Entire trial | 11.10 | 11.20 | 11.20 | 0.04 | 0.23 | - | - | <0.01 | 0.40 |
| Treatment 2 | SEM | Contrast p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Items | CON | OS | OL | T | t | T * t | p 3 | T * p | |
| pH | |||||||||
| Pre-weaning | 6.26 b | 6.43 a | 6.44 a | 0.05 | <0.05 | <0.01 | 0.69 | - | - |
| Post-weaning | 6.13 b | 6.41 a | 6.32 a | 0.05 | <0.01 | 0.72 | 0.46 | - | - |
| Entire trial | 6.19 b | 6.42 a | 6.38 a | 0.03 | <0.01 | - | - | <0.05 | 0.44 |
| NH3-N (mmol/L) | |||||||||
| Pre-weaning | 17.54 a | 13.03 b | 13.63 b | 0.59 | <0.01 | <0.01 | <0.01 | - | - |
| Post-weaning | 9.02 a | 6.23 b | 5.64 b | 0.37 | <0.01 | <0.01 | 0.36 | - | - |
| Entire trial | 13.29 a | 9.65 b | 9.64 b | 0.52 | <0.01 | - | - | <0.01 | 0.49 |
| Treatment 2 | SEM | Contrast p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Items | CON | OS | OL | T | t | T * t | p 3 | T * p | |
| Acetate (%) | |||||||||
| Pre-weaning | 50.61 | 51.39 | 51.71 | 0.009 | 0.81 | <0.01 | 0.28 | - | - |
| Post-weaning | 44.44 b | 49.05 a | 49.11 a | 0.009 | <0.05 | <0.01 | 0.67 | - | - |
| Entire trial | 47.50 | 50.41 | 50.61 | 0.014 | 0.24 | - | - | <0.05 | 0.67 |
| Propionate (%) | |||||||||
| Pre-weaning | 26.39 | 26.23 | 28.40 | 0.007 | 0.34 | <0.01 | 0.07 | - | - |
| Post-weaning | 41.29 a | 34.67 b | 37.10 b | 0.009 | <0.01 | 0.61 | 0.20 | - | - |
| Entire trial | 33.86 a | 30.46 b | 32.71 a | 0.009 | <0.05 | - | - | <0.01 | <0.05 |
| Butyrate (%) | |||||||||
| Pre-weaning | 12.31 a | 13.30 a | 11.34 b | 0.020 | <0.05 | <0.01 | 0.06 | - | - |
| Post-weaning | 10.42 b | 13.39 a | 11.04 b | 0.007 | <0.05 | <0.05 | 0.23 | - | - |
| Entire trial | 12.30 | 13.57 | 11.55 | 0.007 | 0.12 | - | - | 0.34 | 0.24 |
| Valerate (%) | |||||||||
| Pre-weaning | 9.38 | 9.05 | 8.34 | 0.007 | 0.50 | <0.01 | 0.38 | - | - |
| Post-weaning | 3.85 | 2.89 | 2.75 | 0.004 | 0.16 | <0.01 | 0.77 | - | - |
| Entire trial | 6.68 | 5.95 | 5.53 | 0.005 | 0.29 | - | - | <0.01 | 0.91 |
| TVFA (mmol/L) | |||||||||
| Pre-weaning | 87.56 a | 82.69 ab | 73.92 b | 3.79 | <0.05 | <0.01 | 0.65 | - | - |
| Post-weaning | 189.03 | 172.66 | 170.19 | 8.58 | 0.33 | <0.05 | 0.87 | - | - |
| Entire trial | 133.62 a | 123.66 ab | 119.99 b | 4.86 | 0.12 | - | - | <0.01 | 0.85 |
| C2/C3 | |||||||||
| Pre-weaning | 2.55 | 2.43 | 2.19 | 0.23 | 0.30 | <0.01 | 0.15 | - | - |
| Post-weaning | 1.12 b | 1.49 a | 1.42 a | 0.09 | <0.05 | 0.32 | 0.61 | - | - |
| Entire trial | 1.82 | 1.96 | 1.81 | 0.13 | 0.67 | - | - | <0.01 | 0.24 |
| Treatment 2 | SEM | Contrast p-Value | |||||
|---|---|---|---|---|---|---|---|
| Items | CON | OS | OL | T | p 5 | T * p | |
| Diarrhea Frequency (%) 3 | |||||||
| Pre-weaning | 7.04% | 7.29% | 6.91% | 0.006 | 0.89 | - | - |
| Post-weaning | 2.86% | 2.15% | 2.64% | 0.005 | 0.76 | - | - |
| Entire trial | 4.95% | 4.72% | 4.78% | 0.006 | 0.95 | <0.01 | 0.70 |
| Diarrhea Duration (days) 4 | |||||||
| Pre-weaning | 3.94 | 4.08 | 3.87 | 0.33 | 0.89 | - | - |
| Post-weaning | 0.40 | 0.30 | 0.37 | 0.09 | 0.77 | - | - |
| Entire trial | 4.34 | 4.38 | 4.24 | 0.35 | 0.95 | - | - |
| Pneumonia Occurrence (%) | |||||||
| Pre-weaning | 47.92 | 43.36 | 45.13 | - | 0.49 | - | - |
| Treatment 1 | SEM | Contrast p-Value | |||||
|---|---|---|---|---|---|---|---|
| Items | CON | OS | OL | T | t | T * t | |
| Standing (min/d) | 461.60 a | 426.39 b | 430.33 b | 16.47 | <0.05 | 0.27 | 0.81 |
| Lying (min/d) | 877.09 b | 931.03 a | 910.06 b | 35.65 | <0.05 | 0.65 | 0.15 |
| Eating starter (min/d) | 123.38 a | 99.418 b | 101.86 b | 6.45 | <0.05 | 0.77 | 0.87 |
| Eating Hay (min/d) | - | 100.82 | 94.06 | 8.45 | - | 0.58 | 0.35 |
| Drinking (min/d) | 16.73 | 14.79 | 18.36 | 3.26 | 0.46 | 0.22 | 0.55 |
| Walking (min/d) | 20.65 | 15.05 | 15.63 | 2.02 | 0.16 | 0.82 | 0.22 |
| Chewing and Ruminating (min/d) | 133.20 b | 291.41 a | 263.05 a | 15.77 | <0.01 | 0.17 | 0.49 |
| Abnormal Behavior 2 (min/d) | 206.72 a | 78.87 b | 72.65 b | 9.36 | <0.01 | 0.60 | 0.09 |
| Self-Grooming 3 (min/d) | 12.92 | 19.43 | 15.88 | 2.25 | 0.16 | 0.23 | 0.35 |
| Head out of Pen (min/d) | 180.38 a | 127.91 b | 132.45 b | 8.10 | <0.01 | 0.66 | 0.89 |
| Component 1 | Stater 2 | Oat Hay 3 |
|---|---|---|
| DM (%) | 89.48 | 93.20 |
| CP (%, DM) | 29.86 | 6.18 |
| EE (%, DM) | 2.27 | 2.75 |
| Ash (%, DM) | 7.49 | 4.48 |
| NDF (%, DM) | 9.50 | 44.14 |
| ADF (%, DM) | 8.47 | 33.70 |
| ME (Mcal/kg) 4 | 3.23 | 2.87 |
| Calcium (%, DM) | 0.3–2.0 | - |
| Phosphorus (%, DM) | ≥0.2 | - |
| Sodium chloride (%, DM) | 0.2–2.4 | - |
| Lysine (%, DM) | ≥0.2 | - |
| Behavior | Definition of the Behavior |
|---|---|
| Standing 1 | Four hooves on the ground, whether moving or not |
| Lying 1 | Lying on the sternum with head held in a raised position or down |
| Eating starter 1 | Head in starter feed bucket accompanied by chewing movements |
| Eating Hay 1 | Head in hay feed bucket accompanied by chewing movements |
| Drinking 1 | Mouth around drinker |
| Walking 1 | Stepping and moving |
| Chewing and Ruminating 1 | Chewing irregularly and repeatedly without food in the mouth |
| Abnormal Behavior 2 | Calf licked any surface such as fences, floors, windshields |
| Self-Grooming 1 | Calf licked itself with its tongue |
| Head out of Pen 3 | Calf’s head out of the pen to look around and do not engage in any feeding activities |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, J.; Chen, T.; Alugongo, G.M.; Khan, M.Z.; Li, T.; Ma, J.; Liu, S.; Wang, W.; Wang, Y.; Li, S.; et al. Effect of the Length of Oat Hay on Growth Performance, Health Status, Behavior Parameters and Rumen Fermentation of Holstein Female Calves. Metabolites 2021, 11, 890. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120890
Xiao J, Chen T, Alugongo GM, Khan MZ, Li T, Ma J, Liu S, Wang W, Wang Y, Li S, et al. Effect of the Length of Oat Hay on Growth Performance, Health Status, Behavior Parameters and Rumen Fermentation of Holstein Female Calves. Metabolites. 2021; 11(12):890. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120890
Chicago/Turabian StyleXiao, Jianxin, Tianyu Chen, Gibson Maswayi Alugongo, Muhammad Zahoor Khan, Tingting Li, Jing Ma, Shuai Liu, Wei Wang, Yajing Wang, Shengli Li, and et al. 2021. "Effect of the Length of Oat Hay on Growth Performance, Health Status, Behavior Parameters and Rumen Fermentation of Holstein Female Calves" Metabolites 11, no. 12: 890. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120890