
Behavioral and Metabolome Differences between C57BL/6 and DBA/2 Mouse Strains: Implications for Their Use as Models for Depression- and Anxiety-Like Phenotypes

Abstract
:1. Introduction
2. Results
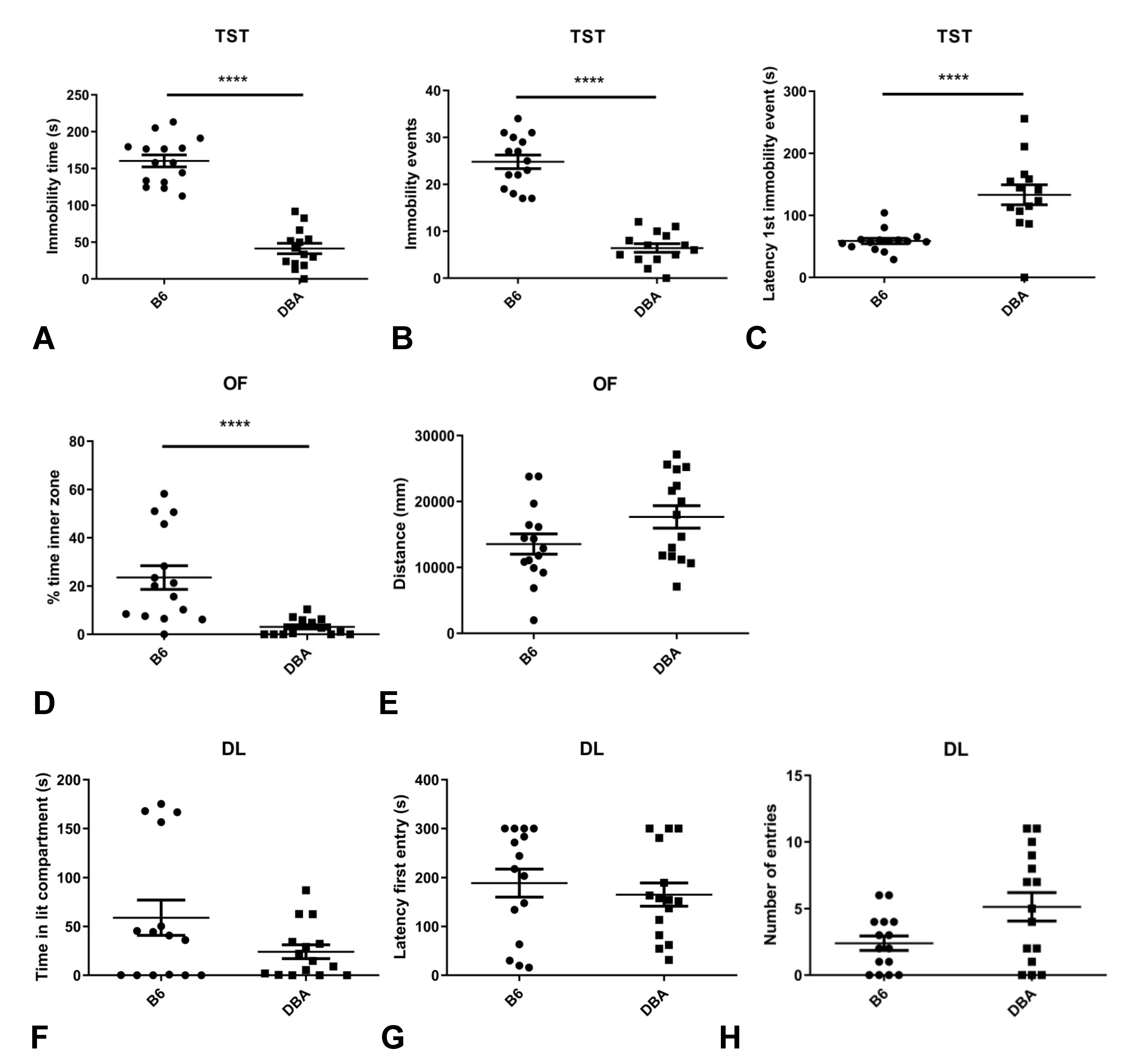
2.1. Different Behavioral Profiles in B6 and DBA Mice
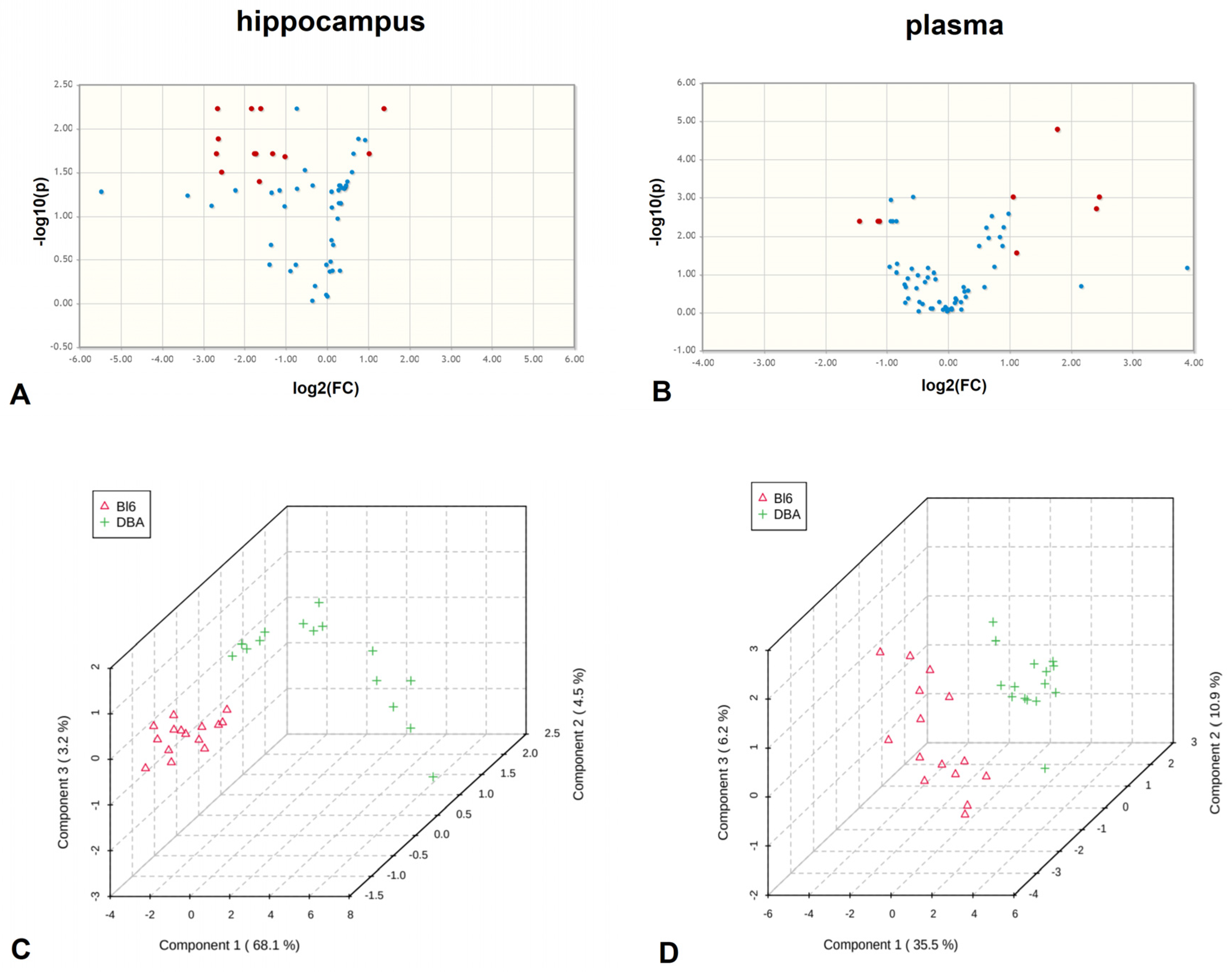
2.2. Distinct Metabolomic Profiles in B6 and DBA Mice Hippocampi
2.3. Distinct Metabolite Profiles in Plasma of B6 and DBA Mice
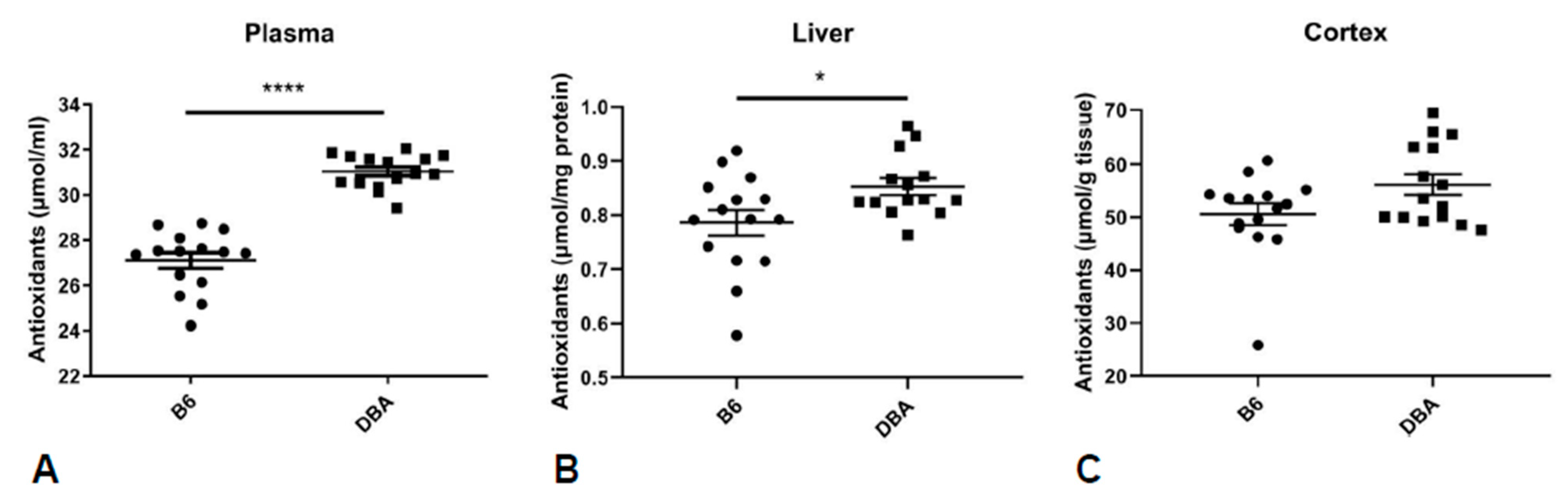
2.4. Common Metabolite Network Differences in B6 vs. DBA Mice
3. Discussion
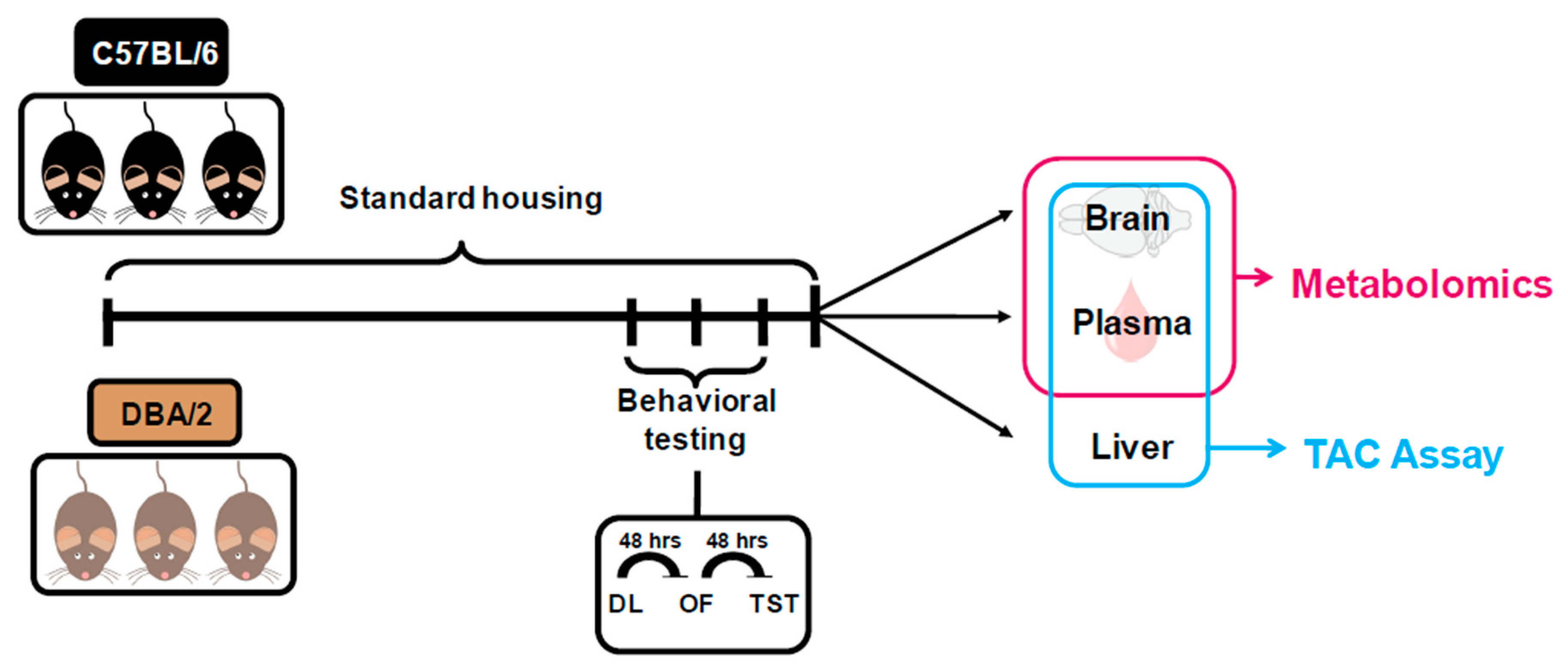
4. Material and Methods
4.1. Animals
4.2. Behavioral Testing and Sample Collection
4.3. Targeted Metabolomics Sample Preparation and Measurement
4.4. Targeted Metabolomics Data Analysis
4.5. Total Antioxidant Capacity
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DL | dark-light box |
| FDR | false discovery rate |
| NMDA | N-methyl-D-aspartic acid |
| OF | open field |
| PLSDA | partial least square discriminant analysis |
| SAM | significance analysis of microarrays |
| SRM | selected reaction monitoring |
| TAC | total antioxidant capacity |
| TCA | tricarboxylic acid cycle |
| TST | tail suspension test |
References
- Griebel, G.; Belzung, C.; Perrault, G.; Sanger, D.J. Differences in anxiety-related behaviours and in sensitivity to diazepam in inbred and outbred strains of mice. Psychopharmacology 2000, 148, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Ogundele, O.M.; Lee, C.C. Contrasting characteristic behaviours among common laboratory mouse strains. R. Soc. Open Sci. 2019, 6, 190574. [Google Scholar] [CrossRef] [Green Version]
- Ohl, F.; Roedel, A.; Binder, E.; Holsboer, F. Impact of high and low anxiety on cognitive performance in a modified hole board test in C57BL/6 and DBA/2 mice. Eur. J. Neurosci. 2003, 17, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Podhorna, J.; Brown, R.E. Strain differences in activity and emotionality do not account for differences in learning and memory performance between C57BL/6 and DBA/2 mice. Genes Brain Behav. 2002, 1, 96–110. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Gunn, R.K.; Brown, R.E. What are we measuring when we test strain differences in anxiety in mice? Behav. Genet. 2013, 43, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Sanger, D.J.; Perrault, G. Genetic differences in the mouse defense test battery. Aggress Behav. 1997, 23, 19–31. [Google Scholar] [CrossRef]
- Võikar, V.; Polus, A.; Vasar, E.; Rauvala, H. Long-term individual housing in C57BL/6J and DBA/ mice: Assessment of behavioral consequences. Genes Brain Behav. 2005, 4, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Kajiwara, Y.; Hirano, K.; Yamada, S.; Tagawa, N.; Kobayashi, Y.; Hotta, Y.; Yamada, J. Mouse strain differences in immobility and sensitivity to fluvoxamine and desipramine in the forced swimming test: Analysis of serotonin and noradrenaline transporter binding. Eur. J. Pharmacol. 2008, 592, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Naumenko, V.S.; Tibeikina, M.A.; Kulikov, A.V. Serotonin transporter, 5-HT1A receptor, and behavior in DBA/2J mice in comparison with four inbred mouse strains. J. Neurosci. Res. 2009, 87, 3649–3657. [Google Scholar] [CrossRef]
- Yuan, M.; Breitkopf, S.B.; Yang, X.M.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filiou, M.D.; Asara, J.M.; Nussbaumer, M.; Teplytska, L.; Landgraf, R.; Turck, C.W. Behavioral extremes of trait anxiety in mice are characterized by distinct metabolic profiles. J. Psychiatr. Res. 2014, 58, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Humer, E.; Probst, T.; Pieh, C. Metabolomics in Psychiatric Disorders: What We Learn from Animal Models. Metabolites 2020, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Pu, J.; Liu, Y.; Gui, S.; Zhong, X.; Song, X.; Xu, S.; Zhang, H.; Wang, H.; Zhou, W.; et al. Metabolomic analysis of animal models of depression. Metab. Brain Dis. 2020, 35, 979–990. [Google Scholar] [CrossRef]
- Park, D.I.; Dournes, C.; Sillaber, I.; Uhr, M.; Asara, J.M.; Gassen, N.C.; Rein, T.; Ising, M.; Webhofer, C.; Filiou, M.D.; et al. Purine and pyrimidine metabolism: Convergent evidence on chronic antidepressant treatment response in mice and humans. Sci. Rep. 2016, 6, 35317. [Google Scholar] [CrossRef] [Green Version]
- Vlaikou, A.M.; Nussbaumer, M.; Komini, C.; Lambrianidou, A.; Konidaris, C.; Trangas, T.; Filiou, M.D. Exploring the crosstalk of glycolysis and mitochondrial metabolism in psychiatric disorders and brain tumors. Eur. J. Neurosci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Sandi, C. Anxiety and Brain Mitochondria: A Bidirectional Crosstalk. Trends Neurosci. 2019, 42, 573–588. [Google Scholar] [CrossRef]
- Filiou, M.D.; Zhang, Y.; Teplytska, L.; Reckow, S.; Gormanns, P.; Maccarrone, G.; Frank, E.; Kessler, M.S.; Hambsch, B.; Nussbaumer, M.; et al. Proteomics and metabolomics analysis of a trait anxiety mouse model reveals divergent mitochondrial pathways. Biol. Psychiatry 2011, 70, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Iris, F.; Filiou, M.; Turck, C.W. Differential proteomics analyses reveal anxiety-associated molecular and cellular mechanisms in cingulate cortex synapses. AJPN 2014, 2, 25–42. [Google Scholar]
- Zhang, Y.; Filiou, M.D.; Reckow, S.; Gormanns, P.; Maccarrone, G.; Kessler, M.S.; Frank, E.; Hambsch, B.; Holsboer, F.; Landgraf, R.; et al. Proteomic and metabolomic profiling of a trait anxiety mouse model implicate affected pathways. Mol. Cell Proteom. 2011, 10. [Google Scholar] [CrossRef] [Green Version]
- Lopes, S.; Teplytska, L.; Vaz-Silva, J.; Dioli, C.; Trindade, R.; Morais, M.; Webhofer, C.; Maccarrone, G.; Almeida, O.F.X.; Turck, C.W.; et al. Tau Deletion Prevents Stress-Induced Dendritic Atrophy in Prefrontal Cortex: Role of Synaptic Mitochondria. Cereb. Cortex 2017, 27, 2580–2591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, M.; McEwen, B.S.; Epel, E.S.; Sandi, C. An energetic view of stress: Focus on mitochondria. Front. Neuroendocrinol. 2018, 49, 72–85. [Google Scholar] [CrossRef]
- Weckmann, K.; Deery, M.J.; Howard, J.A.; Feret, R.; Asara, J.M.; Dethloff, F.; Filiou, M.D.; Iannace, J.; Labermaier, C.; Maccarrone, G.; et al. Ketamine’s antidepressant effect is mediated by energy metabolism and antioxidant defense system. Sci. Rep. 2017, 7, 15788. [Google Scholar] [CrossRef] [Green Version]
- Misiewicz, Z.; Iurato, S.; Kulesskaya, N.; Salminen, L.; Rodrigues, L.; Maccarrone, G.; Martins, J.; Czamara, D.; Laine, M.A.; Sokolowska, E.; et al. Multi-omics analysis identifies mitochondrial pathways associated with anxiety-related behavior. PLoS Genet. 2019, 15, e1008358. [Google Scholar] [CrossRef]
- Nussbaumer, M.; Asara, J.M.; Teplytska, L.; Murphy, M.P.; Logan, A.; Turck, C.W.; Filiou, M.D. Selective Mitochondrial Targeting Exerts Anxiolytic Effects In Vivo. Neuropsychopharmacology 2016, 41, 1751–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, S.A.; Copeland, C.S.; Uebele, V.N.; Thomson, F.J.; Salt, T.E. Modulation of hippocampal synaptic transmission by the kynurenine pathway member xanthurenic acid and other VGLUT inhibitors. Neuropsychopharmacology 2013, 38, 1060–1067. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P.; Parikh, V. Abnormal neurotransmitter release underlying behavioral and cognitive disorders: Toward concepts of dynamic and function-specific dysregulation. Neuropsychopharmacology 2007, 32, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, C.; Vazquez, M.; Fagiolino, P. Potential Therapeutic Role of Carnitine and Acetylcarnitine in Neurological Disorders. Curr. Pharm. Des. 2020, 26, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Cherix, A.; Larrieu, T.; Grosse, J.; Rodrigues, J.; McEwen, B.; Nasca, C.; Gruetter, R.; Sandi, C. Metabolic signature in nucleus accumbens for anti-depressant-like effects of acetyl-L-carnitine. eLife 2020, 9. [Google Scholar] [CrossRef]
- Pancotto, L.; Mocelin, R.; Marcon, M.; Herrmann, A.P.; Piato, A. Anxiolytic and anti-stress effects of acute administration of acetyl-L-carnitine in zebrafish. PeerJ 2018, 6, e5309. [Google Scholar] [CrossRef] [Green Version]
- Erabi, H.; Okada, G.; Shibasaki, C.; Setoyama, D.; Kang, D.; Takamura, M.; Yoshino, A.; Fuchikami, M.; Kurata, A.; Kato, T.A.; et al. Kynurenic acid is a potential overlapped biomarker between diagnosis and treatment response for depression from metabolome analysis. Sci. Rep. 2020, 10, 16822. [Google Scholar] [CrossRef]
- Liu, H.; Ding, L.; Zhang, H.; Mellor, D.; Wu, H.; Zhao, D.; Wu, C.; Lin, Z.; Yuan, J.; Peng, D. The Metabolic Factor Kynurenic Acid of Kynurenine Pathway Predicts Major Depressive Disorder. Front. Psychiatry 2018, 9, 552. [Google Scholar] [CrossRef] [PubMed]
- Jong, C.J.; Azuma, J. Schaffer, Mechanism underlying the antioxidant activity of taurine: Prevention of mitochondrial oxidant production. Amino Acids 2012, 42, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, Z.; Vlaikou, A.M.; Theodoridou, D.; Komini, C.; Chalkiadaki, G.; Vafeiadi, M.; Margetaki, K.; Trangas, T.; Turck, C.W.; Syrrou, M.; et al. Unraveling the Serum Metabolomic Profile of Post-partum Depression. Front. Neurosci. 2019, 13, 833. [Google Scholar] [CrossRef] [PubMed]
- Frank, E.; Kessler, M.S.; Filiou, M.D.; Zhang, Y.; Maccarrone, G.; Reckow, S.; Bunck, M.; Heumann, H.; Turck, C.W.; Landgraf, R.; et al. Stable isotope metabolic labeling with a novel 15N-enriched bacteria diet for improved proteomic analyses of mouse models for psychopathologies. PLoS ONE 2009, 4, e7821. [Google Scholar] [CrossRef]
- Filiou, M.D.; Teplytska, L.; Otte, D.M.; Zimmer, A.; Turck, C.W. Myelination and oxidative stress alterations in the cerebellum of the G72/G30 transgenic schizophrenia mouse model. J. Psychiatr. Res. 2012, 46, 1359–1365. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hippocampus Metabolites | p Value | q Value | Plasma Metabolites | p Value | q Value | |
|---|---|---|---|---|---|---|
| 2-hydroxy-2-methylbutanedioic acid | 0.0004 | 0.0002 | 1-methyl-histidine | 0.0211 | 0.0078 | |
| 2-hydroxygluterate | 0.0049 | 0.0014 | 7-methylguanosine | 0.0006 | 0.0004 | |
| 2-keto-isovalerate | 0.0347 | 0.0043 | acetylcarnitine DL | 0.0003 | 0.0002 | |
| 2-oxobutanoate* | 0.0021 | 0.0007 | alanine | 0.0086 | 0.0039 | |
| 3-methylphenylacetic acid | 0.0256 | 0.0034 | anthranilate* | 0.0003 | 0.0002 | |
| 4-aminobutyrate | 0.0212 | 0.0030 | carnitine | <0.0001 | <0.0001 | |
| adenine* | 0.0032 | 0.0010 | DL-Pipecolic acid | 0.0002 | 0.0002 | |
| adenosine* | <0.0001 | <0.0001 | glutamine* | <0.0001 | <0.0001 | |
| allantoate | <0.0001 | <0.0001 | glycerate | 0.0009 | 0.0006 | |
| allantoin | 0.0179 | 0.0030 | guanidoacetic acid* | <0.0001 | <0.0001 | |
| aminoadipic acid | 0.0425 | 0.0050 | indole | 0.0164 | 0.0066 | |
| atrolactic acid | 0.0439 | 0.0051 | isocitrate | 0.0030 | 0.0017 | |
| betaine | 0.0126 | 0.0025 | kynurenic acid | 0.0003 | 0.0002 | |
| citrate | 0.0465 | 0.0052 | lactate* | <0.0001 | <0.0001 | |
| creatinine* | <0.0001 | <0.0001 | myo-inositol | 0.0113 | 0.0047 | |
| deoxyribose-phosphate | 0.0244 | 0.0033 | N-acetyl-glutamine | 0.0019 | 0.0011 | |
| D-glucarate | 0.0212 | 0.0030 | orotate | 0.0111 | 0.0047 | |
| D-glyceraldehdye-3-phosphate | 0.0007 | 0.0004 | p-aminobenzoate* | 0.0003 | 0.0002 | |
| dimethylglycine* | 0.0004 | 0.0002 | phenylalanine | 0.0003 | 0.0002 | |
| fructose-1,6-bisphosphate* | <0.0001 | <0.0001 | purine | 0.0003 | 0.0002 | |
| fumarate | 0.0339 | 0.0043 | pyrophosphate | 0.0244 | 0.0087 | |
| glyoxylate* | 0.0018 | 0.0006 | ribose-phosphate | 0.0066 | 0.0032 | |
| homocysteic acid | 0.0212 | 0.0030 | S-adenosyl-L-methionine* | 0.0002 | 0.0002 | |
| hypoxanthine | 0.0147 | 0.0028 | sn-glycerol-3-phosphate* | <0.0001 | <0.0001 | |
| inosine | 0.0089 | 0.0022 | taurine* | 0.0066 | 0.0032 | |
| isocitrate* | 0.0060 | 0.0015 | tryptophan | 0.0039 | 0.0021 | |
| lactate | 0.0126 | 0.0025 | uric acid | 0.0208 | 0.0078 | |
| myo-inositol* | <0.0001 | <0.0001 | ||||
| oxaloacetate* | 0.0109 | 0.0025 | ||||
| pantothenate* | 0.0007 | 0.0004 | ||||
| phosphorylcholine | 0.0121 | 0.0025 | ||||
| proline | 0.0161 | 0.0030 | ||||
| pyroglutamic acid | 0.0058 | 0.0015 | ||||
| pyrophosphate | 0.0181 | 0.0030 | ||||
| SBP | 0.0018 | 0.0006 | ||||
| S-ribosyl-L-homocysteine* | 0.0014 | 0.0006 | ||||
| taurine | 0.0186 | 0.0030 | ||||
| thymine* | 0.0014 | 0.0006 | ||||
| urea | 0.0212 | 0.0030 | ||||
| xanthine | 0.0356 | 0.0043 | ||||
| xanthurenic acid | 0.0209 | 0.0030 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filiou, M.D.; Nussbaumer, M.; Teplytska, L.; Turck, C.W. Behavioral and Metabolome Differences between C57BL/6 and DBA/2 Mouse Strains: Implications for Their Use as Models for Depression- and Anxiety-Like Phenotypes. Metabolites 2021, 11, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020128
Filiou MD, Nussbaumer M, Teplytska L, Turck CW. Behavioral and Metabolome Differences between C57BL/6 and DBA/2 Mouse Strains: Implications for Their Use as Models for Depression- and Anxiety-Like Phenotypes. Metabolites. 2021; 11(2):128. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020128
Chicago/Turabian StyleFiliou, Michaela D., Markus Nussbaumer, Larysa Teplytska, and Christoph W. Turck. 2021. "Behavioral and Metabolome Differences between C57BL/6 and DBA/2 Mouse Strains: Implications for Their Use as Models for Depression- and Anxiety-Like Phenotypes" Metabolites 11, no. 2: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020128