
Untargeted Metabolic Profiling of 4-Fluoro-Furanylfentanyl and Isobutyrylfentanyl in Mouse Hepatocytes and Urine by Means of LC-HRMS

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. 4F-FUF Metabolic Profile In Vitro and In Vivo
2.2. Isobutyrylfentanyl Metabolic Profile In Vitro and In Vivo
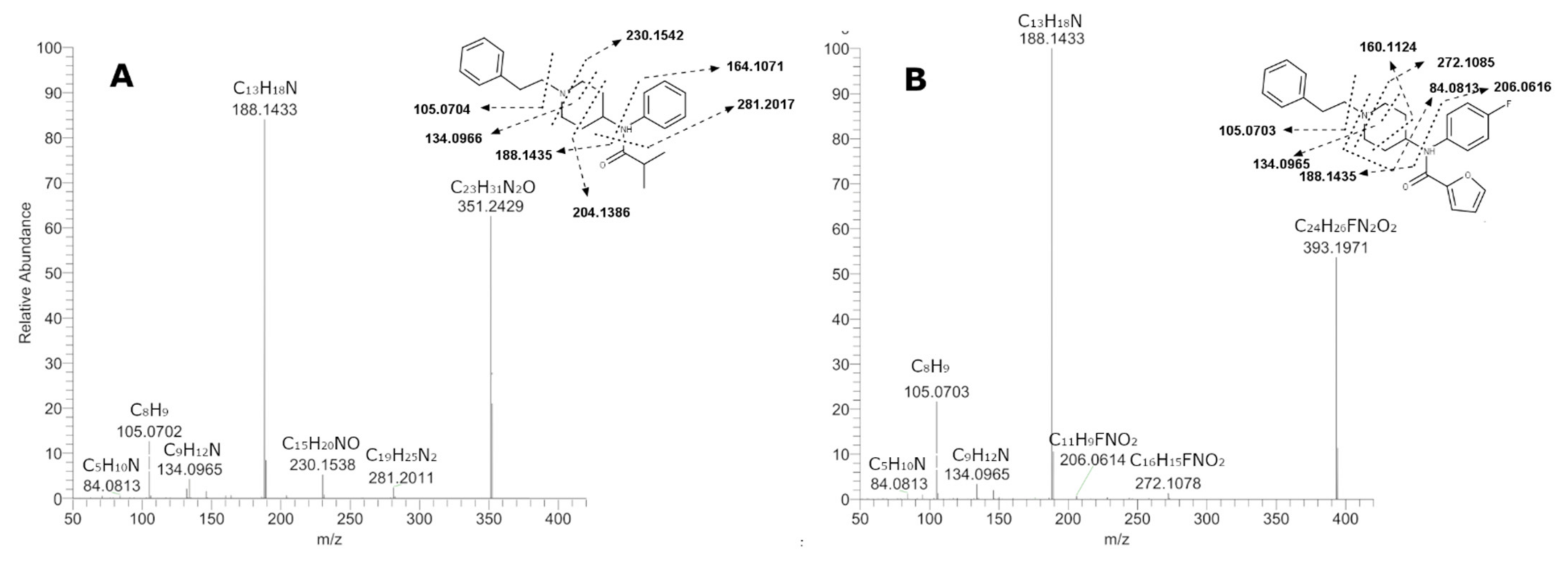
2.3. Elucidation of Metabolites Structure
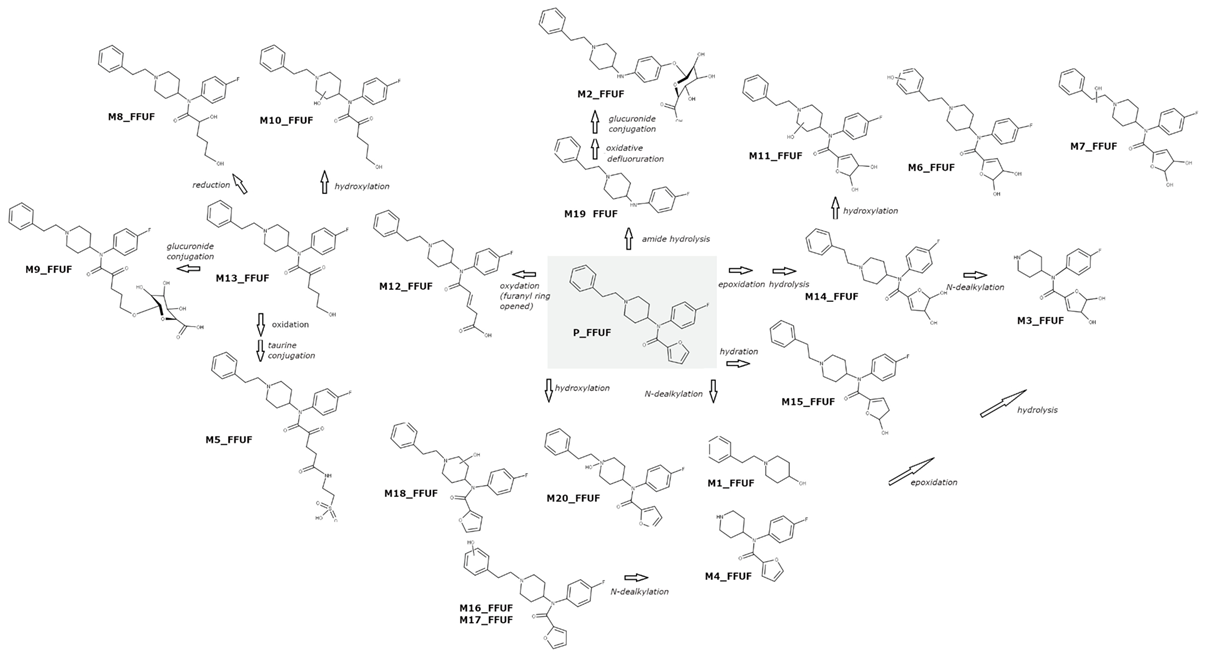
2.3.1. 4F-FUF Metabolites
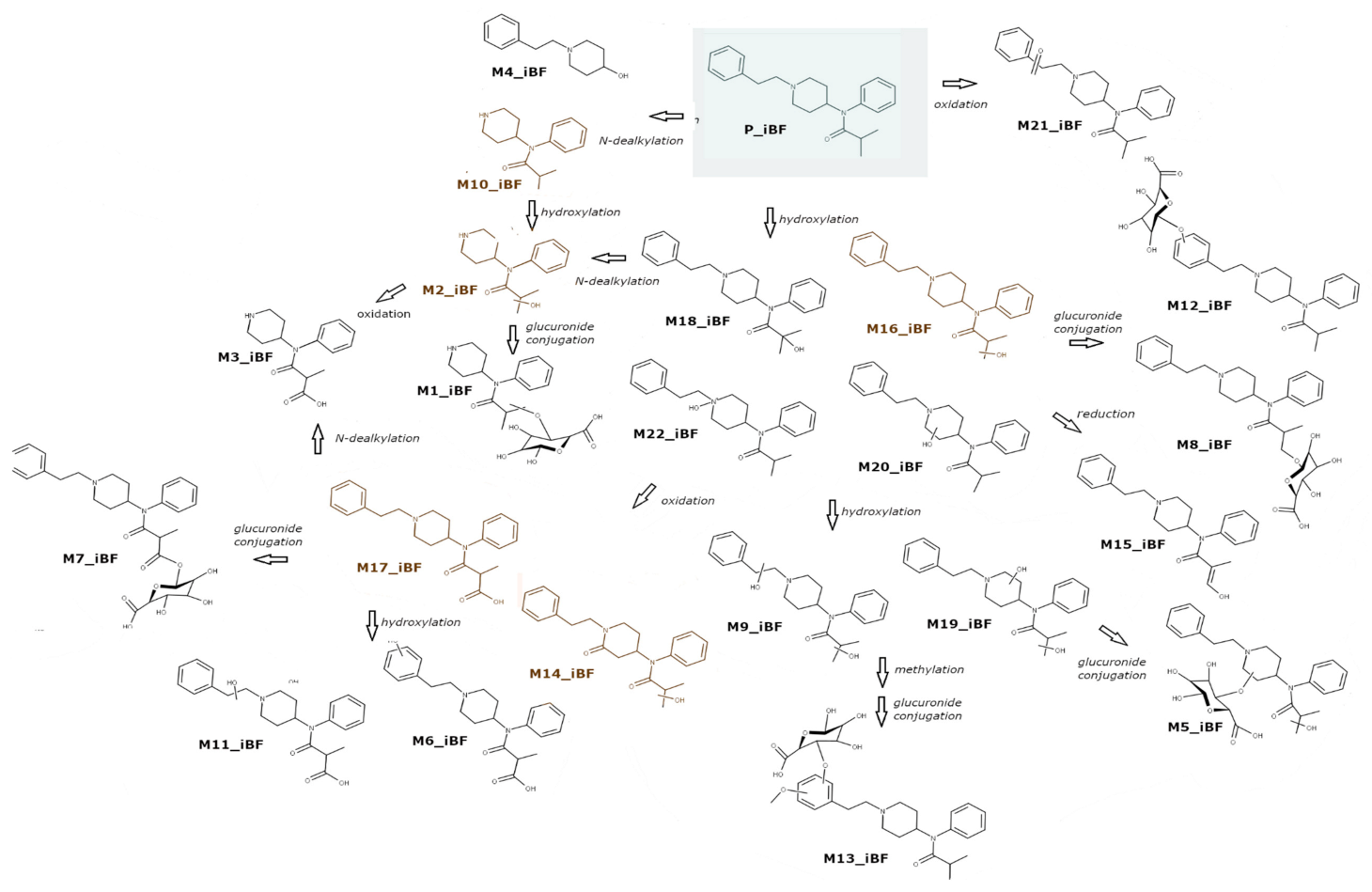
2.3.2. iBF Metabolites
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. In Vitro Incubation Using Mouse Hepatocytes
3.3. In Vivo Study on Mice
3.4. LC-HRMS Analysis
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- King, L.A.; Kicman, A.T. A brief history of “new psychoactive substances”. Drug Test. Anal. 2011, 3, 401–403. [Google Scholar] [CrossRef]
- Van Hout, M.C.; Hearne, E. New psychoactive substances (NPS) on cryptomarket fora: An exploratory study of characteristics of forum activity between NPS buyers and vendors. Int. J. Drug Policy 2017, 40, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Favretto, D.; Pascali, J.P.; Tagliaro, F. New challenges and innovation in forensic toxicology: Focus on the “New Psychoactive Substances”. J. Chromatogr. A 2013, 1287, 84–95. [Google Scholar] [CrossRef]
- Vincenti, F.; Montesano, C.; Cellucci, L.; Gregori, A.; Fanti, F.; Compagnone, D.; Curini, R.; Sergi, M. Combination of pressurized liquid extraction with dispersive liquid liquid micro extraction for the determination of sixty drugs of abuse in hair. J. Chromatogr. A 2019, 1605, 360348. [Google Scholar] [CrossRef]
- Montesano, C.; Vannutelli, G.; Gregori, A.; Ripani, L.; Compagnone, D.; Curini, R.; Sergi, M. Broad screening and identification of novel psychoactive substances in plasma by high-performance liquid chromatography-high-resolution mass spectrometry and post-run library matching. J. Anal. Toxicol. 2016, 40, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richeval, C.; Gaulier, J.-M.; Romeuf, L.; Allorge, D.; Gaillard, Y. Case report: Relevance of metabolite identification to detect new synthetic opioid intoxications illustrated by U-47700. Int. J. Legal Med. 2019, 133, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhang, H.; Humphreys, W.G. Drug metabolite profiling and identification by high-resolution mass spectrometry. J. Biol. Chem. 2011, 286, 25419–25425. [Google Scholar] [CrossRef] [Green Version]
- Swortwood, M.J.; Ellefsen, K.N.; Wohlfarth, A.; Diao, X.; Concheiro-Guisan, M.; Kronstrand, R.; Huestis, M.A. First metabolic profile of PV8, a novel synthetic cathinone, in human hepatocytes and urine by high-resolution mass spectrometry. Anal. Bioanal. Chem. 2016, 408, 4845–4856. [Google Scholar] [CrossRef] [PubMed]
- Montesano, C.; Vannutelli, G.; Fanti, F.; Vincenti, F.; Gregori, A.; Togna, A.R.; Canazza, I.; Marti, M.; Sergi, M. Identification of MT-45 metabolites: In silico prediction, in vitro incubation with rat hepatocytes and in vivo confirmation. J. Anal. Toxicol. 2017, 41, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagmann, L.; Frankenfeld, F.; Park, Y.M.; Herrmann, J.; Fischmann, S.; Westphal, F.; Müller, R.; Flockerzi, V.; Meyer, M.R. How to Study the Metabolism of New Psychoactive Substances for the Purpose of Toxicological Screenings—A Follow-Up Study Comparing Pooled Human Liver S9, HepaRG Cells, and Zebrafish Larvae. Front. Chem. 2020, 8, 539. [Google Scholar] [CrossRef] [PubMed]
- Grafinger, K.E.; Wilke, A.; König, S.; Weinmann, W. Investigating the ability of the microbial model Cunninghamella elegans for the metabolism of synthetic tryptamines. Drug Test. Anal. 2019, 11, 721–729. [Google Scholar] [CrossRef]
- Richter, L.H.J.; Herrmann, J.; Andreas, A.; Park, Y.M.; Wagmann, L.; Flockerzi, V.; Müller, R.; Meyer, M.R. Tools for studying the metabolism of new psychoactive substances for toxicological screening purposes—A comparative study using pooled human liver S9, HepaRG cells, and zebrafish larvae. Toxicol. Lett. 2019, 305, 73–80. [Google Scholar] [CrossRef]
- Wohlfarth, A.; Scheidweiler, K.B.; Pang, S.; Zhu, M.; Castaneto, M.; Kronstrand, R.; Huestis, M.A. Metabolic characterization of AH-7921, a synthetic opioid designer drug: In vitro metabolic stability assessment and metabolite identification, evaluation of in silico prediction, and in vivo confirmation. Drug Test. Anal. 2016, 8, 779–781. [Google Scholar] [CrossRef] [Green Version]
- EMCDDA Trendspotter Study on Fentanyl in Europe Report from an EMCDDA Expert Meeting. 2012. Available online: https://www.emcdda.europa.eu/system/files/publications/748/TD3112230ENN_Fentanyl_400350.pdf (accessed on 28 December 2020).
- Shoff, E.N.; Zaney, M.E.; Kahl, J.H.; Hime, G.W.; Boland, D.M. Qualitative identification of fentanyl analogs and other opioids in postmortem cases by UHPLC-Ion Trap-MSn. J. Anal. Toxicol. 2017, 41, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Manral, L.; Gupta, P.K.; Ganesan, K.; Malhotra, R.C. Gas chromatographic retention indices of fentanyl and analogues. J. Chromatogr. Sci. 2008, 46, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Staeheli, S.N.; Baumgartner, M.R.; Gauthier, S.; Gascho, D.; Jarmer, J.; Kraemer, T.; Steuer, A.E. Time-dependent postmortem redistribution of butyrfentanyl and its metabolites in blood and alternative matrices in a case of butyrfentanyl intoxication. Forensic Sci. Int. 2016, 266, 170–177. [Google Scholar] [CrossRef] [PubMed]
- European Monitoring Centre for Drugs European Drug Report; European Union: Brussels, Belgium, 2019; ISBN 9789291686926.
- Watanabe, S.; Vikingsson, S.; Roman, M.; Green, H.; Kronstrand, R.; Wohlfarth, A. In Vitro and In Vivo Metabolite Identification Studies for the New Synthetic Opioids Acetylfentanyl, Acrylfentanyl, Furanylfentanyl, and 4-Fluoro-Isobutyrylfentanyl. AAPS J. 2017, 19, 1102–1122. [Google Scholar] [CrossRef] [Green Version]
- Goggin, M.M.; Nguyen, A.; Janis, G.C. Identification of unique metabolites of the designer opioid furanyl fentanyl. J. Anal. Toxicol. 2017, 41, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Gaulier, J.; Richeval, C.; Gicquel, T.; Hugbart, C.; Le Dare, B.; Allorge, D.; Morel, I. In vitro characterization of NPS metabolites produced by human liver microsomes and the HepaRG cell line using liquid chromatography-high resolution mass spectrometry (LC-HRMS) analysis: Application to furanyl fentanyl. Curr. Pharm. Biotechnol. 2017, 18, 806–814. [Google Scholar] [CrossRef]
- Kanamori, T.; Iwata, Y.T.; Segawa, H.; Yamamuro, T.; Kuwayama, K.; Tsujikawa, K.; Inoue, H. Metabolism of butyrylfentanyl in fresh human hepatocytes: Chemical synthesis of authentic metabolite standards for definitive identification. Biol. Pharm. Bull. 2019, 42, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Steuer, A.E.; Williner, E.; Staeheli, S.N.; Kraemer, T. Studies on the metabolism of the fentanyl-derived designer drug butyrfentanyl in human in vitro liver preparations and authentic human samples using liquid chromatography-high resolution mass spectrometry (LC-HRMS). Drug Test. Anal. 2017, 9, 1085–1092. [Google Scholar] [CrossRef]
- Gundersen, P.O.M.; Åstrand, A.; Gréen, H.; Josefsson, M.; Spigset, O.; Vikingsson, S. Metabolite Profiling of Ortho-, Meta- And Para-Fluorofentanyl by Hepatocytes and High-Resolution Mass Spectrometry. J. Anal. Toxicol. 2019, 44, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Armenian, P.; Vo, K.T.; Barr-Walker, J.; Lynch, K.L. Fentanyl, fentanyl analogs and novel synthetic opioids: A comprehensive review. Neuropharmacology 2018, 134, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincenti, F.; Pagano, F.; Montesano, C.; Sciubba, F.; Di Cocco, M.E.; Gregori, A.; Rosa, F.D.; Lombardi, L.; Sergi, M.; Curini, R. Multi-analytical characterization of 4-fluoro-furanyl fentanyl in a drug seizure. Forensic Chem. 2020, 21, 100283. [Google Scholar] [CrossRef]
- Varshneya, N.B.; Walentiny, D.M.; Moisa, L.T.; Walker, T.D.; Akinfiresoye, L.R.; Beardsley, P.M. Opioid-like antinociceptive and locomotor effects of emerging fentanyl-related substances. Neuropharmacology 2019, 151, 171–179. [Google Scholar] [CrossRef]
- Wallgren, J.; Vikingsson, S.; Rautio, T.; Nasr, E.; Åstrand, A.; Watanabe, S.; Kronstrand, R.; Gréen, H.; Dahlén, J.; Wu, X.; et al. Structure Elucidation of Urinary Metabolites of Fentanyl and Five Fentanyl Analogs using LC-QTOF-MS, Hepatocyte Incubations and Synthesized Reference Standards. J. Anal. Toxicol. 2020, 21, 993–1003. [Google Scholar] [CrossRef]
- Kokko, H.; Hall, P.D.; Afrin, L.B. Fentanyl-associated syndrome of inappropriate antidiuretic hormone secretion. Pharmacotherapy 2002, 22, 1188–1192. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A.; Rubio, V.Y.; Garrett, T.J. Impact of matrix effects and ionization efficiency in non-quantitative untargeted metabolomics. Metabolomics 2019, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- Park, B.K.; Kitteringham, N.R.; O’Neill, P.M. Metabolism of fluorine-containing drugs. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 443–470. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.A. Reactive metabolites in the biotransformation of molecules containing a furan ring. Chem. Res. Toxicol. 2013, 26, 6–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chieffi, C.; Camuto, C.; De-Giorgio, F.; de la Torre, X.; Diamanti, F.; Mazzarino, M.; Trapella, C.; Marti, M.; Botrè, F. Metabolic profile of the synthetic drug 4,4′-dimethylaminorex in urine by LC–MS-based techniques: Selection of the most suitable markers of its intake. Forensic Toxicol. 2020. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Biotransformation | Rt (min) | Formula | Measured m/z | Mass Error (PPM) | Diagnostic Ions |
|---|---|---|---|---|---|---|
| - | ||||||
| M1_FFUF | Oxidative N-dealkylation | 4.05 | C13H20NO | 206.1547 | 1.02 | 188.1433, 105.0702, 56.0603 |
| M2_FFUF | Amide hydrolysis + oxidative defluorination + glucuronidation | 4.08 | C25H33N2O7 | 473.2289 | 0.26 | 188.1433, 297.1960, 105.0702 |
| M3_FFUF | Dihydrodiol formation + N-dealkylation | 4.10 | C16H20FN2O4 | 323.1404 | −0.96 | 84.0814, 194.0810, 166.0863 |
| M4_FFUF | Oxidative N-dealkylation | 5.29 | C16H18FN2O2 | 289.1346 | −2.18 | 84.0814, 206.0611, 56.0503 |
| M5_FFUF | Oxidation (furanyl ring opened) + oxidation to carbonyl metabolite + taurine conjugation | 5.35 | C26H33FN3O6S | 534.2069 | −0.95 | 188.1433, 105.0703, 299.1917, 409.1917 |
| M6_FFUF | Dihydrodiol formation + hydroxylation | 5.37 | C24H28FN2O5 | 443.1973 | −2.09 | 121.0650, 204.1382, 323.1401 |
| M7_FFUF | Dihydrodiol formation + hydroxylation | 5.64 | C24H28FN2O5 | 443.1969 | −2.99 | 204.1382, 121.0650, 335.1400 |
| M8_FFUF | Oxidation (furanyl ring opened) + reduction | 5.65 | C24H32FN2O3 | 415.2384 | −3.12 | 188.0702, 105.0702, 299.1918 |
| M9_FFUF | Oxidation (furanyl ring opened + glucuronidation) | 5.67 | C30H39FN2O9 | 589.2552 | −1.59 | 188.1433, 105.0702, 413.2233, 299.1917 |
| M10_FFUF | Oxidation (furanyl ring opened) + hydroxylation | 5.80 | C24H30FN2O4 | 429.2186 | −0.84 | 186.1277, 204.1386, 299.1918 |
| M11_FFUF | Dihydrodiol formation + hydroxylation | 5.90 | C24H28FN2O5 | 443.1976 | −1.41 | 425.1868, 186.1277, 134.0965 |
| M12_FFUF | Oxidation (furanyl ring opened) | 6.50 | C24H28FN2O3 | 411.2078 | −1.45 | 188.1434, 105.0703, 299.1921 |
| M13_FFUF | Oxidation (furanyl ring opened) | 6.60 | C24H30FN2O3 | 413.2237 | −0.84 | 188.1434, 105.0703, 299.1916 |
| M14_FFUF | Dihydrodiol formation | 6.77 | C24H28FN2O4 | 427.2027 | −1.43 | 188.1434, 105.0703, 335.1401 |
| M15_FFUF | Hydration | 6.92 | C24H28FN2O3 | 411.2085 | 0.25 | 188.1434, 105.0703 |
| M16_FFUF | Hydroxylation | 7.02 | C24H26FN2O3 | 409.1915 | −3.04 | 204.1382, 121.0650 |
| M17_FFUF | Hydroxylation | 7.35 | C24H26FN2O3 | 409.1915 | −3.04 | 204.1382, 121.0650 |
| M18_FFUF | Hydroxylation | 7.87 | C24H26FN2O3 | 409.1912 | −3.78 | 204.1385, 186.1278, 391.1817 |
| M19_FFUF | Amide hydrolysis | 9.43 | C19H24FN2 | 299.1918 | −1.84 | 105.0703, 204.1385, 186.1277 |
| P_FFUF | - | 9.62 | C24H26FN2O2 | 393.1974 | −1.10 | - |
| M20_FFUF | N-oxygenation | 10.58 | C24H26FN2O3 | 409.1912 | −3.78 | 186.1277, 204.1386, 349.2273 |
| ID | Average (n = 2) Peak Area in Hepatocyte Samples | Average Area in Urine Samples (CV%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.5 h | 1 h | 2 h | 3 h | 1 h | 2 h | 3 h | 4 h | 5 h | 6 h | 6–12 h (n = 6) | 12–24 h (n = 5) | 24–31 h (n = 5) | |
| M1_FFUF | 4.35 × 106 (13) | 5.86 × 106 (6) | 5.52 × 106 (9) | 2.30 × 106 (11) | NS | NS | NS | NS | NS | NS | 1.08 × 107 (51) | 1.15 × 105 (67) | 2.28 × 105 (63) |
| M2_FFUF | 2.77 × 106 (11) | 1.09 × 107 (8) | 2.21 × 107 (16) | 3.39 × 106 (6) | NS | NS | NS | NS | NS | NS | 6.96 × 108 (50) | 4.71 × 107 (44) | 5.54 × 107 (60) |
| M3_FFUF | 1.58 × 106 (5) | 5.42 × 106 (9) | 7.49 × 106 (12) | 2.62 × 106 (10) | NS | NS | NS | NS | NS | NS | 5.55 × 109 (42) | 2.29 × 108 (40) | 2.83 × 108 (77) |
| M4_FFUF | 9.34 × 107 (11) | 1.66 × 108 (16) | 1.66 × 108 (13) | 7.12 × 107 (8) | NS | NS | NS | NS | NS | NS | 3.91 × 108 (47) | 1.79 × 107 (38) | 1.82 × 107 (54) |
| M5_FFUF | NF | 7.53 × 104 (18) | 1.1 × 105 (11) | NF | NS | NS | NS | NS | NS | NS | 7.89 × 107 (51) | 6.6 × 106 (44) | 6.32 × 106 (64) |
| M6_FFUF | NF | NF | NF | NF | NS | NS | NS | NS | NS | NS | 1.96 × 109 (32) | 2.27 × 108 (39) | 3.21 × 108 (56) |
| M7_FFUF | NF | NF | NF | NF | NS | NS | NS | NS | NS | NS | 1.01 × 108 (29) | 1.05 × 107 (44) | 1.84 × 107 (32) |
| M8_FFUF | 5.63 × 106 (9) | 1.07 × 107 (11) | 9.98 × 106 (11) | 5.62 × 106 (13) | NS | NS | NS | NS | NS | NS | 3.05 × 107 (50) | 1.26 × 106 (14) | 1.37 × 106 (56) |
| M9_FFUF | 2.94 × 105 (8) | 7.74 × 105 (13) | 1.16 × 105 (14) | 2.03E5 (11) | NS | NS | NS | NS | NS | NS | 3.81 × 107 (66) | 4.89 × 106 (55) | 3.11 × 106 (49) |
| M10_FFUF | 3.08 × 106 (15) | 6.57 × 106 (12) | 5.33 × 106 (7) | 4.52 × 106 (16) | NS | NS | NS | NS | NS | NS | 5.66 × 108 (63) | 3.94 × 107 (43) | 5.70 × 107 (55) |
| M11_FFUF | 3.47 × 106 (6) | 8.55 × 106 (15) | 1.01 × 107 (18) | 3.17 × 106 (18) | NS | NS | NS | NS | NS | NS | 7.06 × 108 (44) | 9.71 × 107 (21) | 2.23 × 108 (41) |
| M12_FFUF | 6.09 × 106 (9) | 1.28 × 107 (17) | 3.35 × 106 (11) | 2.56 × 106 (15) | NS | NS | NS | NS | NS | NS | 9.22 × 106 (73) | 1.26 × 104 (51) | 5.66 × 104 (62) |
| M13_FFUF | 8.02 × 107 (10) | 1.4 × 108 (9) | 6.60 × 107 (11) | 7.67 × 107 (20) | NS | NS | NS | NS | NS | NS | 1.55 × 108 (69) | 2.04 × 106 (31) | 2.44 × 106 (55) |
| M14_FFUF | 3.59 × 108 (4) | 6.33 × 108 (18) | 7.15 × 108 (12) | 2.57 × 108 (12) | NS | NS | NS | NS | NS | NS | 1.77 × 1010 (55) | 2.85 × 109 (19) | 3.33 × 109 (50) |
| M15_FFUF | NF | NF | NF | NF | NS | NS | NS | NS | NS | NS | 8.11 × 106 (47) | 9.88 × 105 (33) | 6.44 × 105 (49) |
| M16_FFUF | 5.11 × 105 (16) | 1.11 × 106 (5) | 4.31 × 106 (7) | 8.31 × 105 (17) | NS | NS | NS | NS | NS | NS | 8.26 × 106 (75) | 3.23 × 106 (59) | 1.51 × 106 (72) |
| M17_FFUF | NF | NF | NF | NF | NS | NS | NS | NS | NS | NS | 2.91 × 106 (59) | NF | NF |
| M18_FFUF | 6.00 × 106 (18) | 1.20 × 107 (9) | 5.75 × 106 (14) | 7.57 × 106 (14) | NS | NS | NS | NS | NS | NS | 3.27 × 106 (61) | NF | NF |
| M19_FFUF | 7.87 × 106 (11) | 4.19 × 107 (16) | 3.78 × 107 (7) | 5.02 × 107 (9) | NS | NS | NS | NS | NS | NS | 3.40 × 108 (32) | 1.69 × 107 (44) | 2.09 × 107 (19) |
| P_FFUF | 4.05 × 108 (12) | 7.47 × 108 (6) | 2.83 × 108 (9) | 3.91 × 108 (13) | NS | NS | NS | NS | NS | NS | 2.60 × 109 (69) | 5.95 × 107 (71) | 8.62 × 107 (61) |
| M20_FFUF | 1.2 × 106 (8) | 8.86 × 106 (19) | 5.63 × 106 (21) | 1.02 × 107 (12) | NS | NS | NS | NS | NS | NS | 5.60 × 106 (81) | 4.53 × 106 (79) | 1.98 × 106 (85) |
| ID | Biotransformation | Rt (min) | Formula | m/z | Mass Error (PPM) | Diagnostic Ions |
|---|---|---|---|---|---|---|
| - | ||||||
| M1_iBF | N-dealkylation + hydroxylation + gluconidation | 3.61 | C21H31N2O8 | 439.2088 | 1.73 | 263.1753, 84.0814, 180.1019 |
| M2_iBF | N-dealkylation + hydroxylation | 3.86 | C15H23N2O2 | 263.1758 | −0.58 | 84.0814, 245.1648, 177.1387 |
| M3_iBF | N-dealkylation + oxidation | 3.89 | C15H21N2O3 | 277.1546 | −2.23 | 84.0814, 233.1648 |
| M4_iBF | Oxidative N-dealkylation | 4.05 | C13H20NO | 206.1542 | −1.40 | 188.1433, 105.0702 |
| M5_iBF | Dihydroxylation + glucuronidation | 4.78 | C29H39N2O9 | 559.2642 | −2.42 | 263.1390, 383.2328, 204.1382, 116.0709 |
| M6_iBF | Oxidation + hydroxylation | 4.94 | C23H29N2O4 | 397.2119 | −2.10 | 204.1384, 121.0651, 353.2221 |
| M7_iBF | Oxidation to carbonyl metabolite + glucuronidation | 5.34 | C29H37N2O9 | 557.2490 | −1.63 | 188.1434, 337.2272, 105.0703 |
| M8_iBF | Hydroxylation + glucuronidation | 5.37 | C29H39N2O8 | 543.2706 | −0.08 | 188.1433, 367.2378, 105.0702 |
| M9_iBF | Dihydroxylation | 5.37 | C23H31N2O3 | 383.2330 | −1.22 | 204.1384, 186.1278, 365.2223 |
| M10_iBF | Oxidative N-dealkylation | 5.39 | C15H23N2O | 247.1808 | −0.96 | 84.0813, 177.1386, 164.1073 |
| M11_iBF | Oxidation to carbonyl metabolite+ hydroxylation | 5.45 | C23H29N2O4 | 397.2119 | −2.10 | 204.1384, 353.2222, 121.0651 |
| M12_iBF | Hydroxylation +glucuronidation | 5.56 | C29H39N2O8 | 543.2707 | 0.11 | 367.2376, 204.1382, 121.0650 |
| M13_iBF | Dihydroxylation + methylation + glucuronidation | 5.64 | C30H41N2O9 | 573.2809 | −0.53 | 397.2534, 410.1813, 234.1488 |
| M14_iBF | Oxidation + hydroxylation | 5.77 | C23H29N2O3 | 381.2181 | 0.74 | 202.1229, 148.0759, 105.0703 |
| M15_iBF | Hydroxylation | 5.87 | C23H29N2O2 | 365.2225 | −1.10 | 188.1436, 105.0704, 244.1332 |
| M16_iBF | Hydroxylation | 6.09 | C23H31N2O2 | 367.2381 | −1.23 | 188.1434, 105.0703, 246.1486 |
| M17_iBF | Oxidation to carbonyl metabolite | 6.16 | C23H29N2O3 | 381.2190 | 3.10 | 188.1434, 105.0703, 281.2011, 337.2272 |
| M18_iBF | Hydroxylation | 6.43 | C23H31N2O2 | 367.2381 | −1.23 | 188.1434, 105.0703, 281.2013 |
| M19_iBF | Dihydroxylation | 7.81 | C23H31N2O3 | 383.2342 | 1.91 | 105.0703, 186.1274, 275.1753 |
| M20_iBF | Hydroxylation | 7.84 | C23H31N2O2 | 367.2381 | −1.23 | 186.1274, 204.1386, 105.0703 |
| M21_iBF | Oxidation | 8.80 | C23H29N2O2 | 365.2226 | −0.83 | 202.1230, 195.1808, 230.1536 |
| P_iBF | - | 9.61 | C23H31N2O | 351.2428 | −2.39 | - |
| M22_iBF | N-oxidation | 10.54 | C23H29N2O2 | 367.2381 | −1.23 | 186.1274, 105.0703, 204.1386 |
| ID | Average (n = 2) Peak Area in Hepatocyte Samples (CV%) | Average Area in Urine Samples (CV%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.5 h | 1 h | 2 h | 3 h | 1 h (n = 1) | 2 h (n = 2) | 3 h (n = 6) | 4 h | 5 h (n = 6) | 6 h (n = 5) | 6–12 h (n = 6) | 12–24 h (n = 6) | 24–31 h (n = 6) | |
| M1_iBF | NF | NF | NF | NF | 4.08 × 107 | 1.53 × 108 (17) | 9.85 × 108 (74) | NS | 2.65 × 108 (50) | 1.51 × 108 (51) | 8.41 × 107 (63) | 1.94 × 107 (53) | 2.86 × 107 (52) |
| M2_iBF | 1.49 × 107 (5) | 1.64 × 107 (9) | 3.60 × 107 (8) | 2.00 × 107 (10) | 8.09 × 108 | 2.01 × 109 (32) | 3.31 × 109 (51) | NS | 2.76 × 108 (49) | 2.06 × 108 (50) | 1.14 × 109 (31) | 3.55 × 108 (33) | 4.26 × 108 (55) |
| M3_iBF | NF | NF | NF | NF | 3.58 × 107 | 5.58 × 107 (15) | 2.03 × 108 (39) | NS | 1.93 × 108 (46) | 1.20 × 108 (49) | 7.93 × 107 (29) | 1.51 × 107 (43) | 2.01 × 107 (58) |
| M4_iBF | 2.87 × 107 (18) | 1.86 × 107 (10) | 3.88 × 107 (8) | 1.76 × 107 (12) | 9.98 × 107 | 3.37 × 107 (26) | 1.88 × 108 (71) | NS | 6.59 × 107 (33) | 2.80 × 107 (39) | 1.94 × 107 (61) | 3.54 × 106 (44) | 4.88 × 106 (62) |
| M5_iBF | NF | NF | NF | NF | 5.55 × 107 | 3.49 × 107 (32) | 1.43 × 108 (49) | NS | 9.10 × 107 (39) | 8.16 × 107 (61) | 7.42 × 107 (62) | 1.24 × 107 (38) | 2.63 × 107 (43) |
| M6_iBF | NF | NF | NF | NF | 3.44 × 107 | 1.21 × 107 (18) | 1.03 × 108 (52) | NS | 4.97 × 107 (40) | 3.36 × 107 (59) | 2.48 × 107 (36) | 9.57 × 106 (52) | 1.81 × 107 (43) |
| M7_iBF | NF | NF | NF | NF | 1.18 × 108 | 3.91 × 107 (19) | 2.41 × 108 (50) | NS | 2.49 × 108 (33) | 2.26 × 108 (39) | 1.51 × 108 (44) | 2.24 × 107 (42) | 4.14 × 107 (51) |
| M8_iBF | 2.76 × 105 (19) | 1.39 × 106 (11) | 1.45 × 106 (9) | 7.50 × 105 (11) | 1.28 × 108 | 6.02 × 107 (21) | 1.54 × 108 (38) | NS | 1.50 × 108 (52) | 2.75 × 108 (44) | 1.66 × 108 (32) | 3.70 × 107 (41) | 4.94 × 107 (54) |
| M9_iBF | 1.51 × 107 (7) | 1.65 × 107 (9) | 2.12 × 107 (6) | 1.41 × 107 (18) | 4.69 × 106 | 1.60 × 107 (32) | 4.15 × 107 (30) | NS | 5.29 × 107 (53) | 3.59 × 107 (41) | 3.66 × 107 (61) | 3.45 × 106 (59) | 5.55 × 106 (37) |
| M10_iBF | 2.04 × 109 (8) | 2.01 × 109 (6) | 2.68 × 109 (8) | 1.62 × 109 (5) | 6.78 × 109 | 1.21 × 1010 (18) | 1.40 × 1010 (29) | NS | 1.08 × 1010 (50) | 1.16 × 1010 (39) | 6.96 × 109 (51) | 2.35 × 109 (41) | 2.83 × 109 (38) |
| M11_iBF | NF | NF | NF | NF | 1.27 × 108 | 1.74 × 107 (44) | 2.12 × 108 (61) | NS | 1.88 × 108 (48) | 1.18 × 108 (41) | 1.27 × 108 (52) | 2.86 × 107 (32) | 5.49 × 107 (34) |
| M12_iBF | 1.59 × 106 (6) | 1.61 × 106 (5) | 3.47 × 106 (6) | 1.45 × 106 (7) | 7.01 × 107 | 5.06 × 107 (32) | 1.05 × 108 (50) | NS | 9.86 × 107 (33) | 1.53 × 108 (41) | 1.19 × 108 (54) | 3.70 × 107 (61) | 4.84 × 107 (39) |
| M13_iBF | NF | NF | NF | NF | 9.79 × 105 | 1.62 × 106 (11) | 3.54 × 106 (41) | NS | 4.44 × 106 (39) | 1.47 × 107 (39) | 3.41 × 106 (41) | 8.66 × 105 (34) | 8.55 × 105 (31) |
| M14_iBF | 1.73 × 106 (6) | 2.00 × 106 (9) | 2.55 × 106 (10) | 1.70 × 106 (12) | 1.38 × 109 | 5.46 × 108 (36) | 3.40 × 109 (38) | NS | 2.61 × 109 (41) | 2.15 × 109 (49) | 1.51 × 109 (61) | 3.93 × 108 (36) | 6.15 × 108 (18) |
| M15_iBF | 6.61 × 106 (9) | 3.98 × 105 (8) | 6.63 × 106 (9) | 7.50 × 106 (7) | 4.84 × 105 | 8.69 × 105 (18) | 1.45 × 106 (42) | NS | 6.20 × 105 (52) | 1.21 × 106 (51) | 2.93 × 105 (55) | 4.49 × 104 (31) | 2.30 × 104 (54) |
| M16_iBF | 2.71 × 108 (8) | 2.52 × 108 (10) | 2.74 × 108 (8) | 1.42 × 108 (11) | 5.77 × 108 | 2.93 × 108 (21) | 1.26 × 109 (51) | NS | 7.99 × 108 (51) | 4.74 × 108 (59) | 2.93 × 108 (45) | 4.81 × 107 (39) | 7.05 × 107 (56) |
| M17_iBF | 3.29 × 106 (5) | 5.50 × 106 (12) | 1.52 × 107 (7) | 8.62 × 106 (16) | 2.37 × 109 | 9.25 × 108 (32) | 6.17 × 109 (33) | NS | 4.48 × 109 (39) | 3.45 × 109 (57) | 2.57 × 109 (49) | 6.43 × 108 (41) | 1.05 × 109 (51) |
| M18_iBF | 2.54 × 108 (10) | 2.06 × 108 (7) | 2.02 × 108 (6) | 1.20 × 108 (13) | 3.52 × 107 | 1.85 × 106 (33) | 9.57 × 107 (49) | NS | 4.12 × 107 (42) | 2.32 × 107 (33) | 1.63 × 107 (48) | 4.53 × 106 (42) | 5.72 × 106 (49) |
| M19_iBF | 1.82 × 106 (9) | 1.95 × 106 (8) | 1.96 × 106 (5) | 1.13 × 106 (11) | 2.70 × 107 | 4.08 × 107 (19) | 6.93 × 107 (50) | NS | 4.34 × 107 (43) | 2.54 × 107 (39) | 1.51 × 107 (32) | 2.52 × 106 (37) | 5.44 × 106 (48) |
| M20_iBF | 1.40 × 108 (7) | 1.04 × 108 (10) | 9.07 × 107 (7) | 4.57 × 107 (12) | 2.21 × 107 | 5.10 × 107 (21) | 6.80 × 107 (51) | NS | 5.84 × 106 (55) | 4.54 × 106 (37) | 2.21 × 106 (18) | 1.10 × 106 (37) | 8.85 × 105 (51) |
| M21_iBF | 9.47 × 106 (17) | 9.19 × 106 (21) | 6.68 × 106 (10) | 2.24 × 106 (11) | 1.32 × 106 | 8.40 × 106 (22) | 9.30 × 106 (63) | NS | 5.15 × 106 (37) | 3.04 × 106 (35) | 3.56 × 106 (22) | 2.05 × 106 (58) | 5.05 × 106 (58) |
| P_iBF | 1.69 × 109 (11) | 8.78 × 108 (18) | 5.31 × 108 (8) | 2.39 × 108 13) | 2.28 × 109 | 3.82 × 108 (29) | 5.82 × 109 (41) | NS | 1.05 × 109 (39) | 1.23 × 109 (38) | 7.56 × 108 (32) | 8.86 × 107 (56) | 2.33 × 107 (38) |
| M22_iBF | NF | NF | NF | NF | 2.44 × 107 | 2.92 × 107 (31) | 5.78 × 107 (42) | NS | 1.98 × 107 (41) | 1.09 × 107 (48) | 8.65 × 106 (61) | 6.08 × 105 (32) | 1.75 × 106 (42) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montesano, C.; Vincenti, F.; Fanti, F.; Marti, M.; Bilel, S.; Togna, A.R.; Gregori, A.; Di Rosa, F.; Sergi, M. Untargeted Metabolic Profiling of 4-Fluoro-Furanylfentanyl and Isobutyrylfentanyl in Mouse Hepatocytes and Urine by Means of LC-HRMS. Metabolites 2021, 11, 97. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020097
Montesano C, Vincenti F, Fanti F, Marti M, Bilel S, Togna AR, Gregori A, Di Rosa F, Sergi M. Untargeted Metabolic Profiling of 4-Fluoro-Furanylfentanyl and Isobutyrylfentanyl in Mouse Hepatocytes and Urine by Means of LC-HRMS. Metabolites. 2021; 11(2):97. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020097
Chicago/Turabian StyleMontesano, Camilla, Flaminia Vincenti, Federico Fanti, Matteo Marti, Sabrine Bilel, Anna Rita Togna, Adolfo Gregori, Fabiana Di Rosa, and Manuel Sergi. 2021. "Untargeted Metabolic Profiling of 4-Fluoro-Furanylfentanyl and Isobutyrylfentanyl in Mouse Hepatocytes and Urine by Means of LC-HRMS" Metabolites 11, no. 2: 97. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020097