
Potassium Improves Drought Stress Tolerance in Plants by Affecting Root Morphology, Root Exudates, and Microbial Diversity

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
2.1. Experiment I
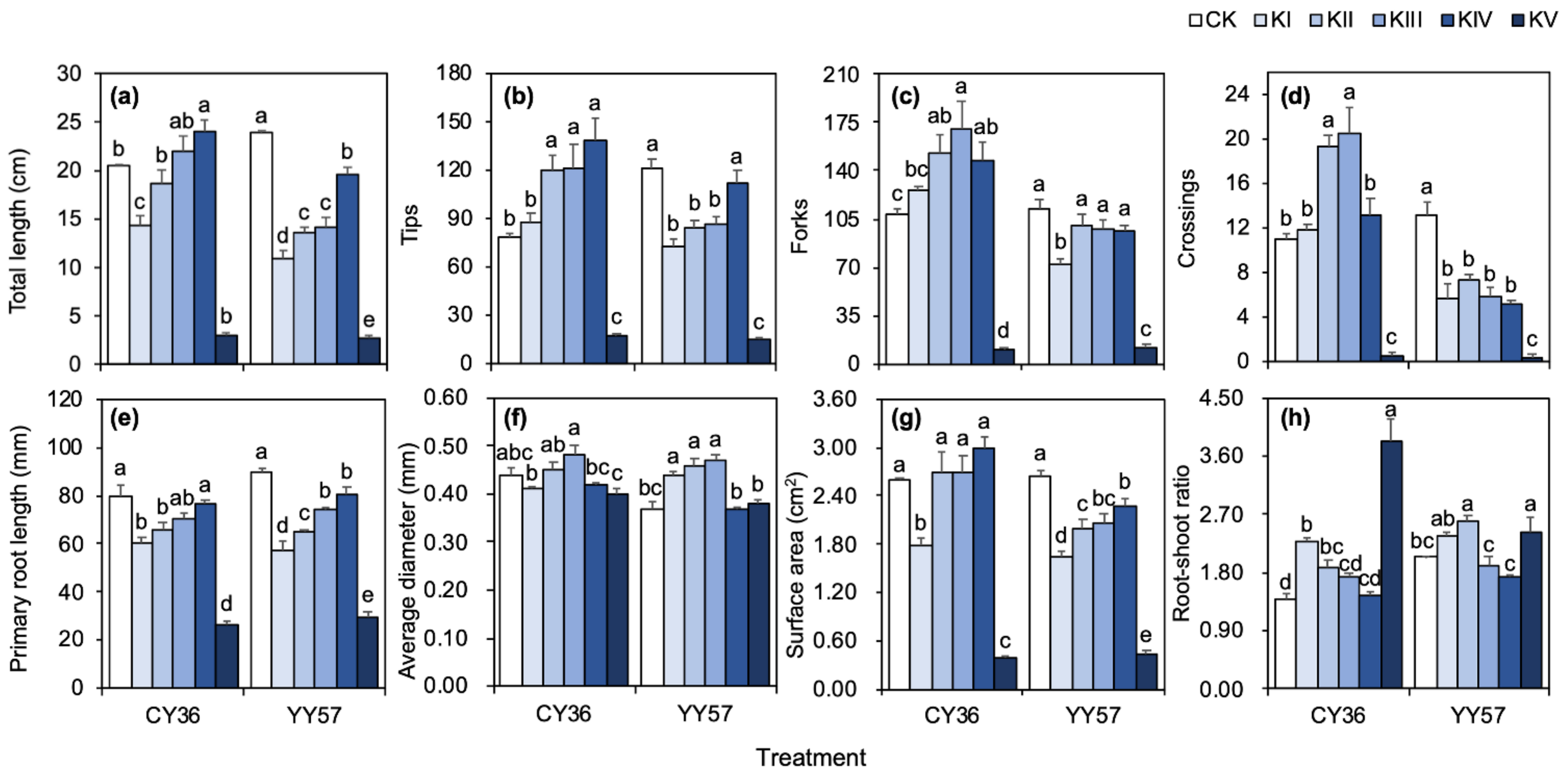
2.1.1. Root Morphological Characters under Different K Supply Levels
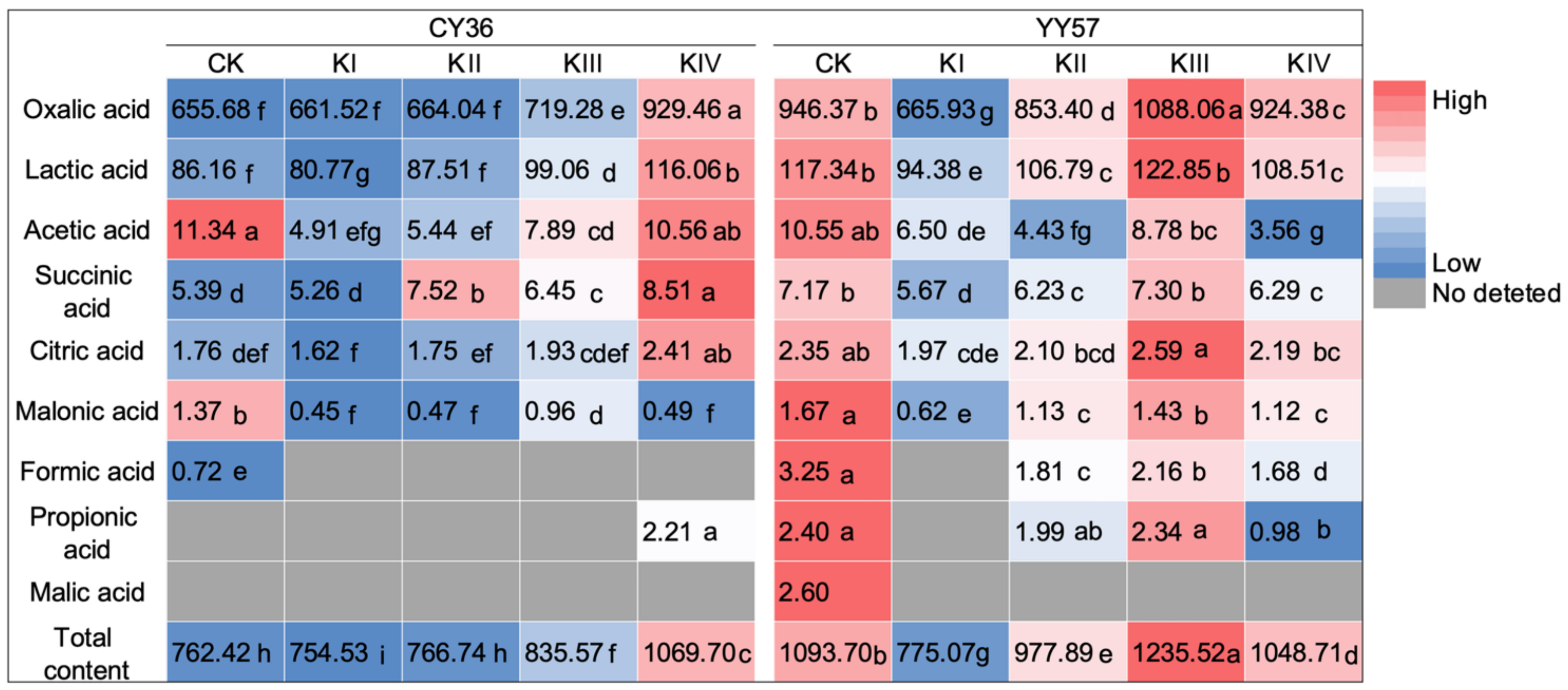
2.1.2. Changes in the Quantity and Composition of Organic Acids with K Supply
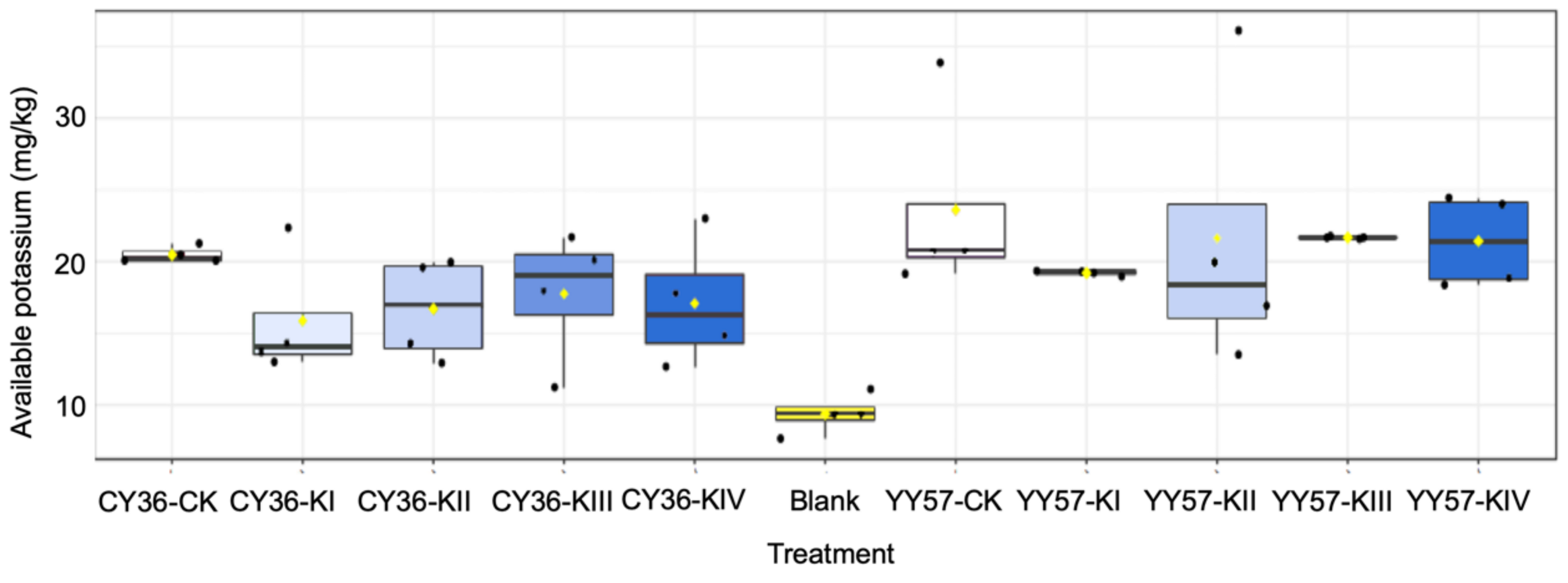
2.1.3. Soil Available K Activated by Organic Acids
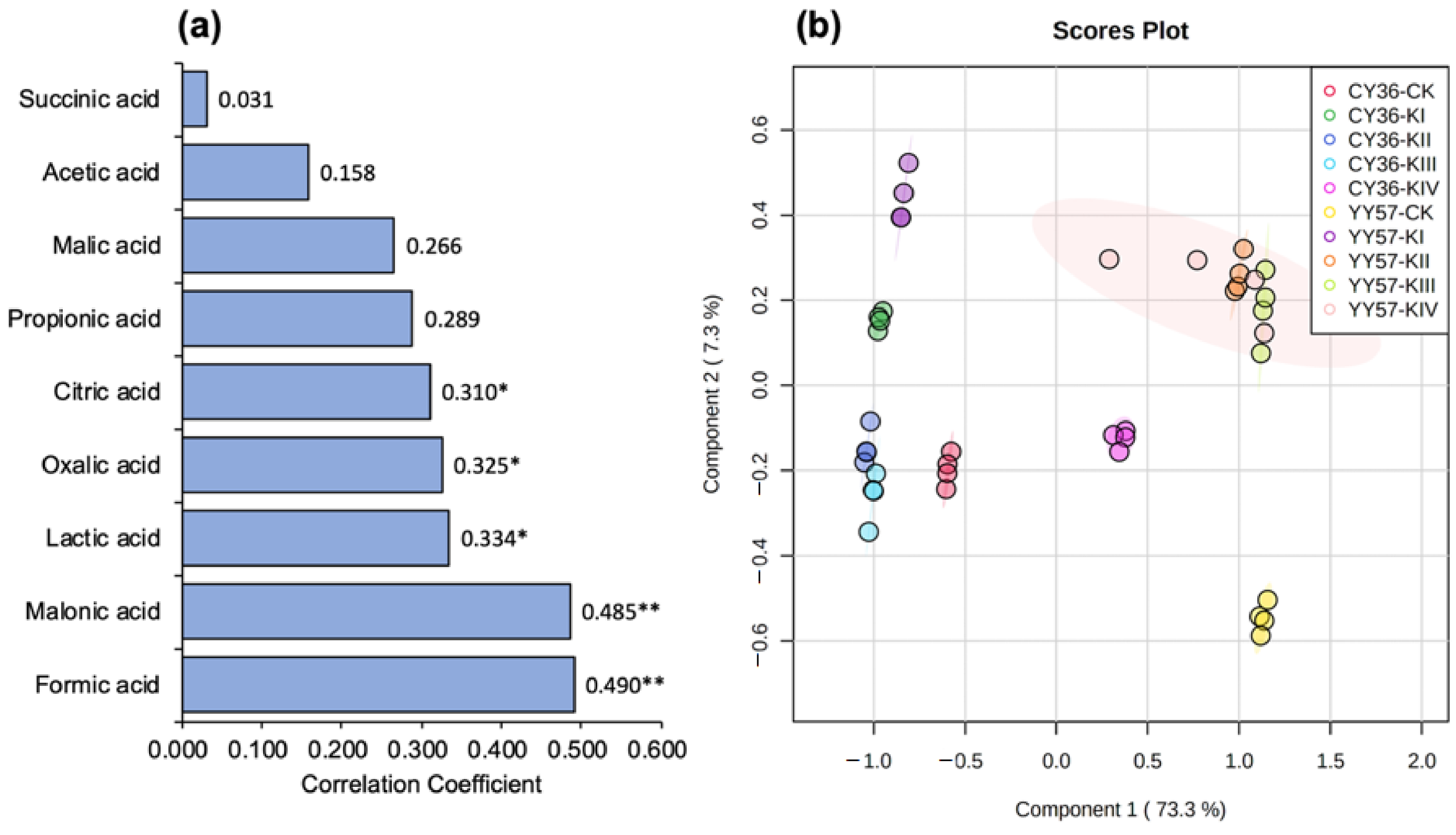
2.1.4. Partial Least Squares Discriminant Analysis
2.2. Experiment II
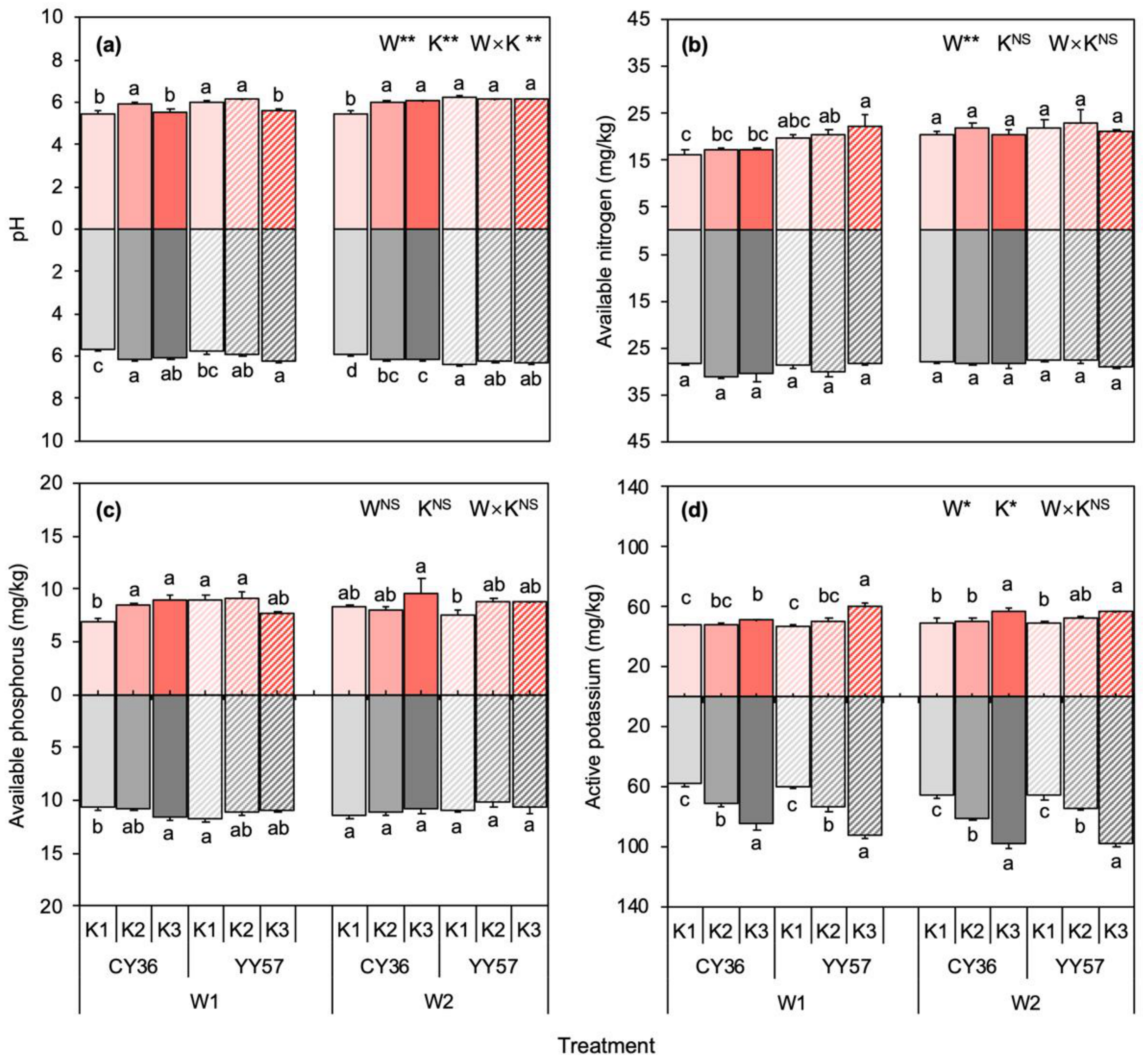
2.2.1. Physicochemical Properties of Rhizosphere and Non-Rhizosphere Soil
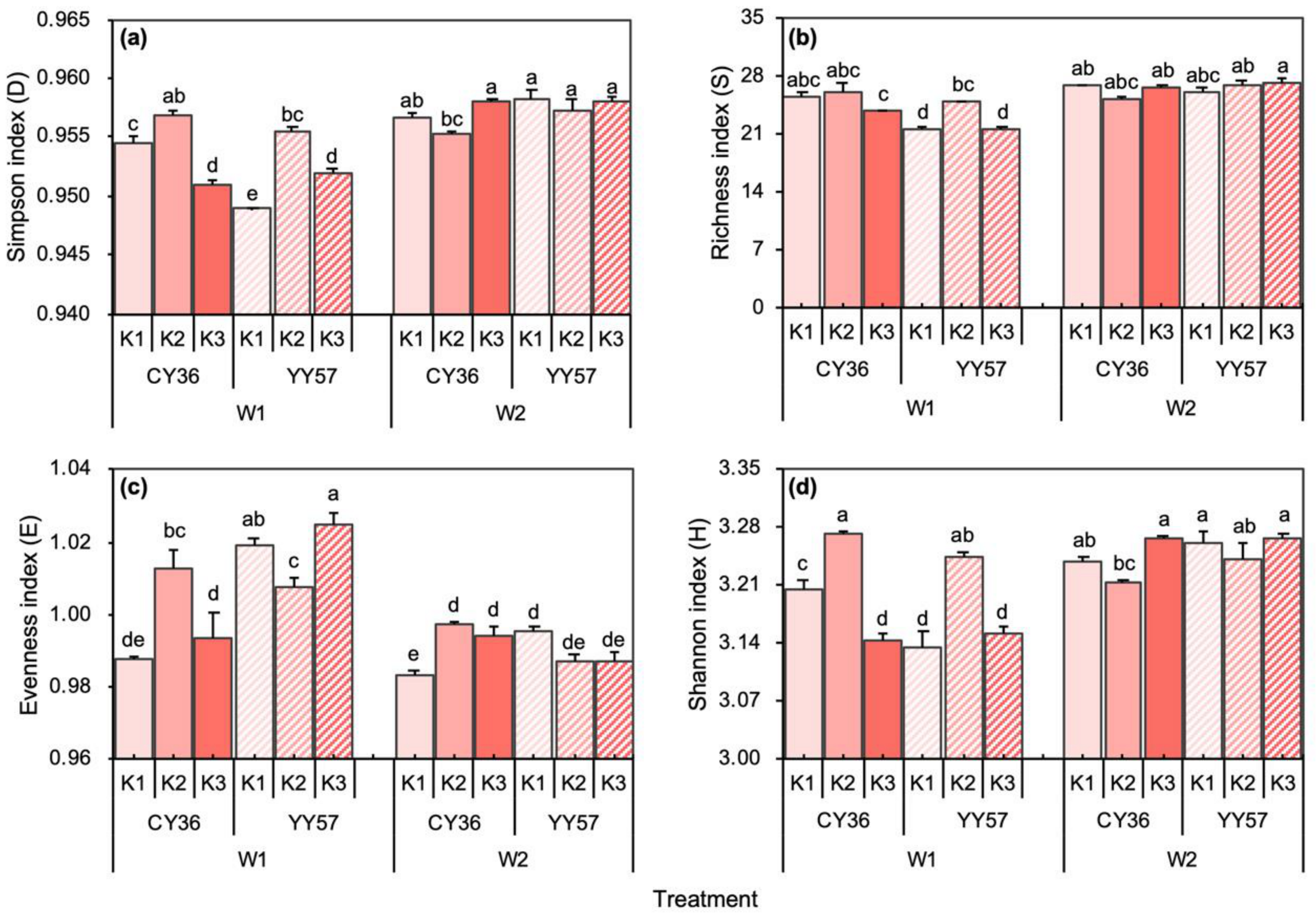
2.2.2. Microbial Diversity Indexes under Water-Potassium Combination
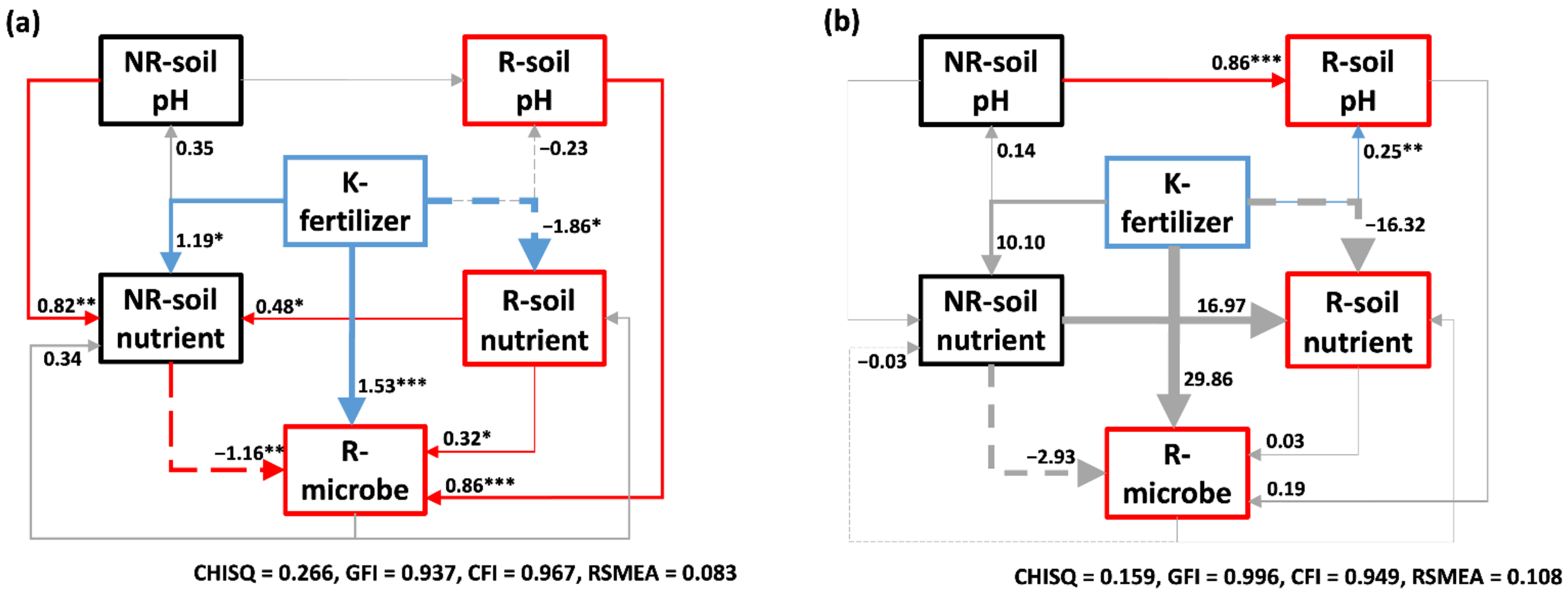
2.2.3. Relationship in the Soil Ecosystem
3. Discussion
4. Materials and Methods
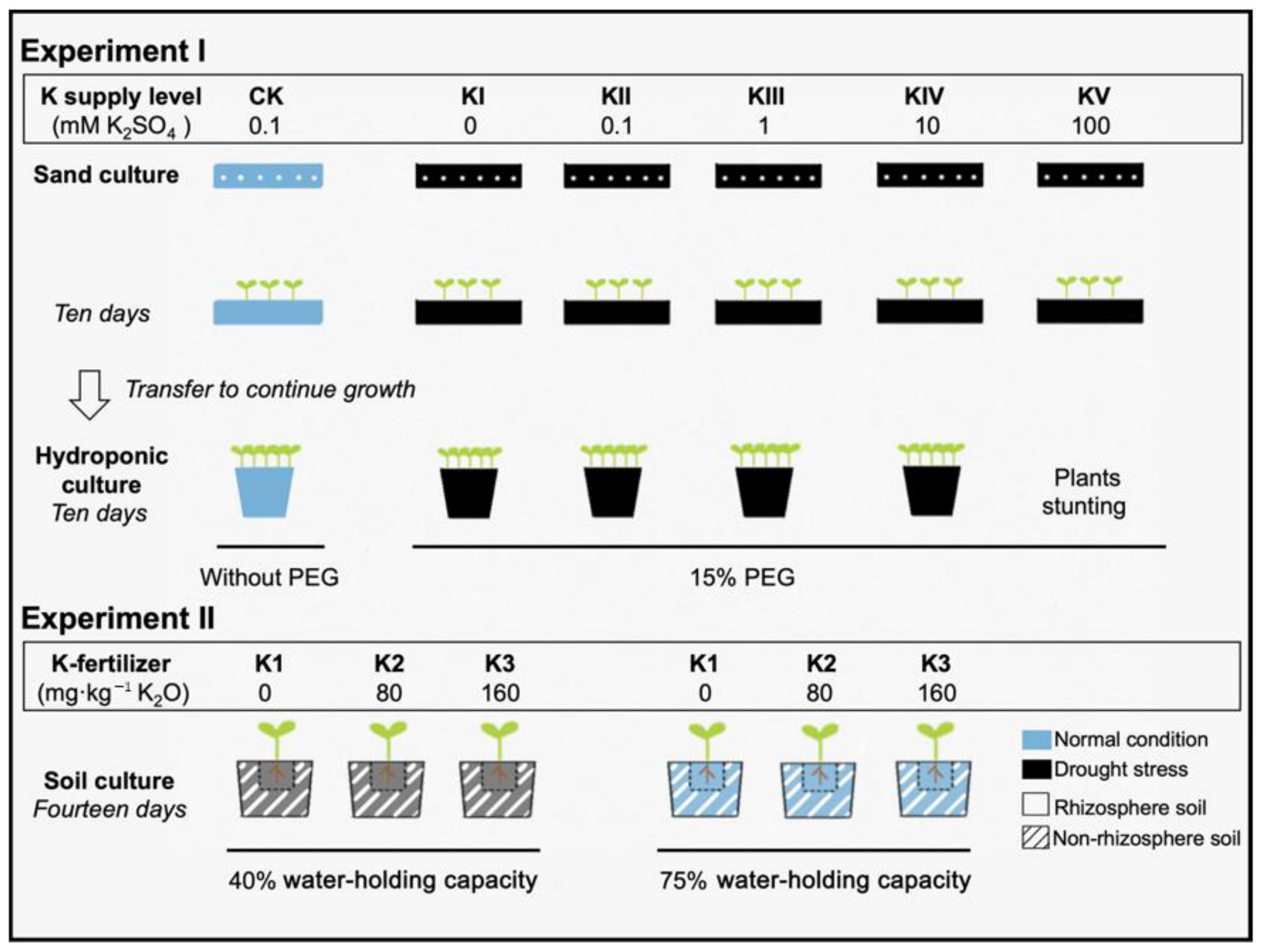
4.1. Experimental Setup
4.1.1. Experiment I: Effects of K Supply Level on Root Morphology and Root Exudates under Drought Stress
4.1.2. Experiment II: Effects of the Water-Potassium Combination on Soil Nutrient and Microbes
4.2. Measurements
4.2.1. Root Morphological Traits
4.2.2. Organic Acids in Root Exudates and Activation on Soil AK
4.2.3. Soil Sampling and Physicochemical Properties
4.2.4. Soil Microbial Community Functional Diversity
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lau, J.A.; Lennon, J.T. Rapid responses of soil microorganisms improve plant fitness in novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Rhodes, C.J. The 2015 Paris climate change conference: COP21. Sci. Prog. 2016, 99, 97–104. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-Deficit stress-induced anatomical changes in higher plants. Cr. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Husen, A.; Iqbal, M.; Aref, I.M. Growth, water status, and leaf characteristics of Brassica carinata under drought and rehydration conditions. Braz. J. Bot. 2014, 37, 217–227. [Google Scholar] [CrossRef]
- Oliet, J.; Planelles, R.; Segura, M.L.; Artero, F.; Jacobs, D.F. Mineral nutrition and growth of containerized Pinus halepensis seedlings under controlled-release fertilizer. Sci. Hortic Amst. 2004, 103, 113–129. [Google Scholar] [CrossRef]
- Zhu, B.; Xu, Q.W.; Zou, Y.G.; Ma, S.M.; Zhang, X.D.; Xie, X.Y.; Wang, L.C. Effect of potassium deficiency on growth, antioxidants, ionome and metabolism in rapeseed under drought stress. Plant Growth Regul. 2020, 90, 455–466. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.R.; Abid, M.; Zhao, W.Q.; Wang, Y.H.; Zhou, Z.G. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Zoerb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Zahoor, R.; Zhao, W.; Abid, M.; Dong, H.; Zhou, Z. Title: Potassium application regulates nitrogen metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress. J. Plant Physiol. 2017, 215, 30–38. [Google Scholar] [CrossRef]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Shahid, A.M.; Saleem, M.F.; Khan, I.H.; Ahmad, S.; Ali, M.; Iqbal, R. Effects and management strategies to mitigate drought stress in oilseed rape (Brassica napus L.): A review. Zemdirbyste 2017, 104, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, W.; Inanaga, S.; Araki, H.; Morita, S.; An, P.; Sonobe, K. Development and distribution of root system in two grain sorghum cultivars originated from Sudan under drought stress. Plant Prod. Sci. 2005, 8, 553–562. [Google Scholar] [CrossRef]
- Vries, F.T.; Williams, A.; Stringer, F.; Willcocks, R.; McEwing, R.; Langridge, H.; Straathof, A.L. Changes in root-exudate-induced respiration reveal a novel mechanism through which drought affects ecosystem carbon cycling. New Phytol. 2019, 224, 132–145. [Google Scholar] [CrossRef]
- Song, F.B.; Han, X.Y.; Zhu, X.C.; Herbert, S.J. Response to water stress of soil enzymes and root exudates from drought and non-drought tolerant corn hybrids at different growth stages. Can. J. Soil Sci. 2012, 92, 501–507. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.H.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Jones, D.L. Organic acids in the rhizosphere—A critical review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Zolla, G.; Badri, D.V.; Bakker, M.G.; Manter, D.K.; Viyanco, J.M. Soil microbiomes vary in their ability to confer drought tolerance to Arab. Appl. Soil Ecol. 2013, 68, 1–9. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic-stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92. [Google Scholar] [CrossRef]
- Magnani, F.; Mencuccini, M.; Grace, J. Age-Related decline in stand productivity: The role of structural acclimation under hydraulic constraints. Plant Cell Environ. 2010, 23, 251–263. [Google Scholar] [CrossRef]
- Muszynska, E.; Labudda, M. Dual role of metallic trace elements in stress biology-from negative to beneficial impact on plants. Int. J. Mol. Sci. 2019, 20, 3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J. Hormesis: Principles and applications. Homeopathy 2015, 104, 69–82. [Google Scholar] [CrossRef]
- Marschner, H. Functions of mineral nutrients: Micronutrients. Miner. Nutr. High. Plants 1995, 313–404. [Google Scholar]
- Yurtseven, E.; Kesmez, G.D.; Unlukara, A. The effects of water salinity and potassium levels on yield, fruit quality and water consumption of a native central anatolian tomato species (Lycopersicon esculantum). Agr. Water Manag. 2005, 78, 128–135. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Jones, D.L.; Darrah, P.R. Influx and efflux of organic-acids across the soil-root interface of Zea-Mays and its implications in rhizosphere C flow. Plant Soil 1995, 173, 103–109. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Randall, P.J. Characterization of Al-stimulated efflux of malate from the apices of Al-tolerant wheat roots. Planta 1995, 196, 103–110. [Google Scholar] [CrossRef]
- Samuels, A.L.; Fernando, M.; Glass, A.D.M. Immunofluorescent localization of plasma membrane H+-ATPase in barley roots and effects of K nutrition. Plant Physiol. 1992, 99, 1509–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.; Roberts, J.K.M. Cytoplasmic malate levels in maize root tips during K+ ion uptake determined by 13C-NMR spectroscopy. Biochim. Biophys. Acta 1991, 1092, 29–34. [Google Scholar] [CrossRef]
- Li, X.; Dong, J.; Chu, W.; Chen, Y.; Duan, Z. The relationship between root exudation properties and root morphological traits of cucumber grown under different nitrogen supplies and atmospheric CO2 concentrations. Plant Soil 2018, 425, 415–432. [Google Scholar] [CrossRef]
- Jiang, C.; Yang, J.; Wei, C.; Xie, D.; Qu, M. Effect of low-molecular-weight organic acids on potassium release from purple rocks. Plant Nutr. Fertil. Sci. 2002, 8, 441–446. [Google Scholar]
- Hocking, P.J. Organic acids exuded from roots in phosphorus uptake and aluminum tolerance of plants in acid soils. Adv. Agron. 2001, 74, 63–97. [Google Scholar]
- Yuan, X.; Geng, S.; Zheng, M.; Xi, X.; Song, D.; Huang, F. Effects of faba bean (Vicia fabaL.) root exudate on soil available nutrients and microbial population in different purple soils. Chin. J. Eco-Agric. 2016, 24, 910–917. [Google Scholar]
- Zhan, L.; Cong, R.; Li, X.; Lu, J.; Wang, J.; Wang, Z.; Liao, Z. Effect of low-molecular-weight organic acids on K+ adsorption kinetics of red soil and yellow-cinnamon soil. Acta Pedol. Sin. 2012, 49, 1147–1157. [Google Scholar]
- Zheng, Y.; Chen, X.; Gong, X.; Bonkowski, M.; Wang, S.; Griffiths, B.; Hu, F.; Liu, M. The geophagous earthworm Metaphire guillelmi effects on rhizosphere microbial community structure and functioning vary with plant species. Geoderma 2020, 379, 114647. [Google Scholar] [CrossRef]
- Tian, J.; Lou, Y.; Gao, Y.; Fang, H.; Liu, S.; Xu, M.; Blagodatskaya, E.; Kuzyakov, Y. Response of soil organic matter fractions and composition of microbial community to long-term organic and mineral fertilization. Biol. Fertil. Soils 2017, 53, 523–532. [Google Scholar] [CrossRef]
- Jiang, L.; Bonkowski, M.; Luo, L.; Kardol, P.; Zhang, Y.; Chen, X.; Li, D.; Xiao, Z.; Hu, F.; Liu, M. Combined addition of chemical and organic amendments enhances plant resistance to aboveground herbivores through increasing microbial abundance and diversity. Biol. Fertil. Soils 2020, 56, 1007–1022. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, D.; Xu, Q.F.; Fuhrmann, J.J.; Li, L.Q.; Pan, G.X.; Li, Y.F.; Qin, H.; Liang, C.F.; Sun, X. Organic carbon quality, composition of main microbial groups, enzyme activities, and temperature sensitivity of soil respiration of an acid paddy soil treated with biochar. Biol. Fertil. Soils 2019, 55, 185–197. [Google Scholar] [CrossRef]
- Bell, C.W.; Fricks, B.E.; Rocca, J.D.; Steinweg, J.M.; McMahon, S.K.; Wallenstein, M.D. High-Throughput fluorometric measurement of potential soil extracellular enzyme activities. Jove J. Vis. Exp. 2013. [Google Scholar] [CrossRef]
- Lan, Z.; Lin, X.; Zhang, W.; Zhang, H.; Wu, Y. Effect of P deficiency on the emergence of Astragalus L. root exudates and mobilization of sparingly soluble phosphorus. Sci. Agric. Sin. 2012, 45, 1521–1531. [Google Scholar]
- Lu, Y.; Abraham, W.R.; Conrad, R. Spatial variation of active microbiota in the rice rhizosphere revealed by in situ stable isotope probing of phospholipid fatty acids. Environ. Microbiol. 2007, 9, 474–481. [Google Scholar] [CrossRef]
- Sun, B.; Chi, J.; Fan, Z.; He, X.; Li, Y.; Tong, C. Determination of organic acids from integrated system of soil and plant. Environ. Sci. Technol. 2010, 33, 130–134. [Google Scholar]
- Li, T.; Ma, G.; Zhang, X. Root exudates of potassium-enrichment genotype grain amaranth and their activation on soil mineral potassium. J. Appl. Ecol. 2006, 17, 368–372. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Zhang, K.; Wang, Z.; Xu, Q.; Liu, B.; Duan, M.; Wang, L. Effect of controlled-release urea fertilizers for oilseed rape (Brassica napus L.) on soil carbon storage and CO2 emission. Environ. Sci. Pollut. Res. 2020, 27, 31983–31994. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Lin, X.G.; Wang, J.H.; Dai, J.; Chen, R.R.; Zhang, J.B.; Wong, M.H. Microbial functional diversity, metabolic quotient, and invertase activity of a sandy loam soil as affected by long-term application of organic amendment and mineral fertilizer. J. Soils Sediments 2011, 11, 271–280. [Google Scholar] [CrossRef]
- Doncaster, C.P. Structural equation modeling and natural systems. Fish Fish. 2006, 8, 368–369. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Fu, H.; Zhu, B.; Hussain, H.A.; Zhang, K.; Tian, X.; Duan, M.; Xie, X.; Wang, L. Potassium Improves Drought Stress Tolerance in Plants by Affecting Root Morphology, Root Exudates, and Microbial Diversity. Metabolites 2021, 11, 131. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11030131
Xu Q, Fu H, Zhu B, Hussain HA, Zhang K, Tian X, Duan M, Xie X, Wang L. Potassium Improves Drought Stress Tolerance in Plants by Affecting Root Morphology, Root Exudates, and Microbial Diversity. Metabolites. 2021; 11(3):131. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11030131
Chicago/Turabian StyleXu, Qiwen, Hao Fu, Bo Zhu, Hafiz Athar Hussain, Kangping Zhang, Xiaoqing Tian, Meichun Duan, Xiaoyu Xie, and Longchang Wang. 2021. "Potassium Improves Drought Stress Tolerance in Plants by Affecting Root Morphology, Root Exudates, and Microbial Diversity" Metabolites 11, no. 3: 131. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11030131