
Selection of Potential Yeast Probiotics and a Cell Factory for Xylitol or Acid Production from Honeybee Samples

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Isolated Yeasts from Honey
2.2. Determination of Thermo-, Low PH and Bile Salt Tolerance of Yeast Isolates
2.3. Autoaggregation Ability of Yeast Isolates
2.4. Utilization of Sugars
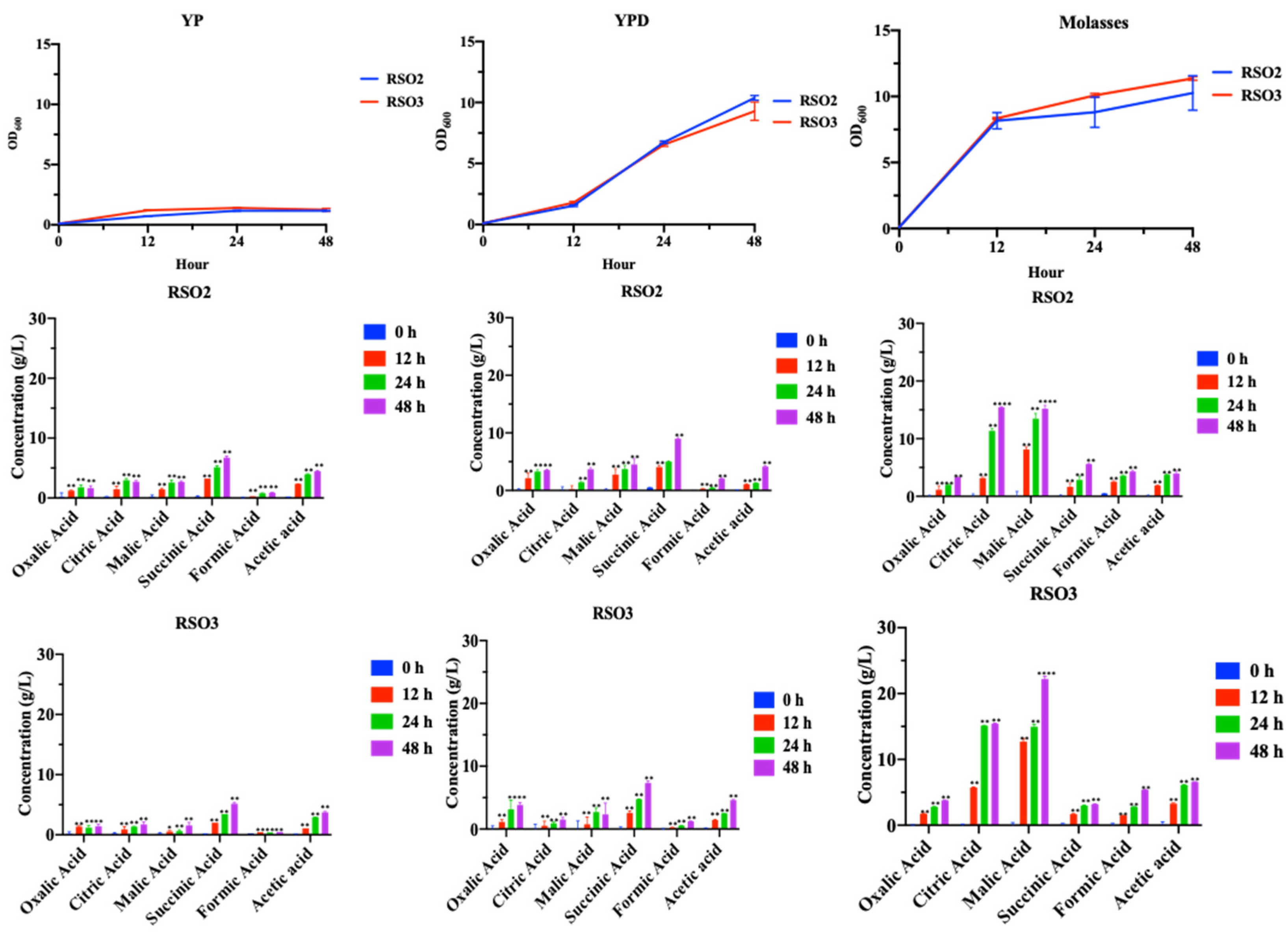
2.5. Organic Acid Production by S. cerevisiae RSO Strains
2.6. Xytitol Production by M. guilliermondii MX Strain
3. Materials and Methods
3.1. Sample Collection and Yeast Isolation
3.2. Sequence Analysis of the ITS Regions
3.3. Phylogenic Tree Analysis Check
3.4. Growth Assays at Increased Temperatures, Low PHS, and 0.3% of Bile Salt
3.5. Autoaggregation Assay
3.6. Carbohydrate Utilization
3.7. Metabolite and Cell Density Analyses
3.8. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afrc, R.F. Probiotics In Man And Animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [CrossRef]
- Clancy, R. Immunobiotics And The Probiotic Evolution. FEMS Immunol. Med. Microbiol. 2003, 38, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H.J.F.M.L. Probiotics, Their Health Benefits And Applications For Developing Healthier Foods: A Review. FEMS Microbiol. Lett. 2020, 334, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forssten, S.D.; Sindelar, C.W.; Ouwehand, A.C. Probiotics From An Industrial Perspective. Anaerobe 2011, 17, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, B.; Rani, G.S.; Kumar, B.K.; Chandrasekhar, B.; Krishna, K.V.; Devi, T.A.; Bhima, B. Evaluating The Probiotic And Therapeutic Potentials Of Saccharomyces Cerevisiae Strain (Obs2) Isolated From Fermented Nectar Of Toddy Palm. AMB Express 2017, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Coelho, B.; Fiorda-Mello, F.; De Melo Pereira, G.V.; Thomaz-Soccol, V.; Rakshit, S.K.; De Carvalho, J.C.; Soccol, C.R.J.F. In Vitro Probiotic Properties And Dna Protection Activity Of Yeast And Lactic Acid Bacteria Isolated From A Honey-Based Kefir Beverage. Foods 2019, 8, 485. [Google Scholar] [CrossRef] [Green Version]
- Otles, S.; Cagindi, O.J.P.J.O.N. Kefir: A Probiotic Dairy-Composition, Nutritional And Therapeutic Aspects. Pak. J. Nutr. 2003, 2, 54–59. [Google Scholar] [CrossRef]
- Ogunremi, O.; Sanni, A.; Agrawal, R.J.J.O.A.M. Probiotic Potentials Of Yeasts Isolated From Some Cereal-Based N Igerian Traditional Fermented Food Products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Heber, D.; Lembertas, A.; Lu, Q.-Y.; Bowerman, S.; Go, V.L.W.J.T.J.O.A.; Medicine, C. An Analysis Of Nine Proprietary Chinese Red Yeast Rice Dietary Supplements: Implications Of Variability In Chemical Profile And Contents. J. Altern. Complementary Med. 2001, 7, 133–139. [Google Scholar] [CrossRef]
- Menezes, A.G.T.; Ramos, C.L.; Dias, D.R.; Schwan, R.F.J.F.R.I. Combination Of Probiotic Yeast And Lactic Acid Bacteria As Starter Culture To Produce Maize-Based Beverages. Food Res. Int. 2018, 111, 187–197. [Google Scholar] [CrossRef]
- Huang, J.S.; Bousvaros, A.; Lee, J.W.; Diaz, A.; Davidson, E.J. Efficacy Of Probiotic Use In Acute Diarrhea In Children: A Meta-Analysis. Dig. Dis. Sci. 2002, 47, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Moslehi-Jenabian, S.; Lindegaard, L.; Jespersen, L. Beneficial Effects Of Probiotic And Food Borne Yeasts On Human Health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Pothoulakis, C. Efficacy And Safety Of The Probiotic Saccharomyces Boulardii For The Prevention And Therapy Of Gastrointestinal Disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerucka, D.; Piche, T.; Rampal, P. Yeast As Probiotics–Saccharomyces Boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, S.; Klinder, A.; Brigidi, P.; Cavina, P.; Costabile, A. Potential Probiotic Kluyveromyces Marxianus B0399 Modulates The Immune Response In Caco-2 Cells And Peripheral Blood Mononuclear Cells And Impacts The Human Gut Microbiota In An In Vitro Colonic Model System. Appl. Environ. Microbiol. 2012, 78, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Moradi, R.; Nosrati, R.; Zare, H.; Tahmasebi, T.; Saderi, H.; Owlia, P.J.I.J.O.M. Screening And Characterization Of In-Vitro Probiotic Criteria Of Saccharomyces And Kluyveromyces Strains. Iran. J. Microbiol. 2018, 10, 123. [Google Scholar] [PubMed]
- Qvirist, L.A.; De Filippo, C.; Strati, F.; Stefanini, I.; Sordo, M.; Andlid, T.; Felis, G.E.; Mattarelli, P.; Cavalieri, D. Isolation, Identification And Characterization Of Yeasts From Fermented Goat Milk Of The Yaghnob Valley In Tajikistan. Front. Microbiol. 2016, 7, 1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illiano, P.; Brambilla, R.; Parolini, C.J.T.F.J. The Mutual Interplay Of Gut Microbiota, Diet And Human Disease. FEBS J. 2020, 287, 833–855. [Google Scholar] [CrossRef]
- Kurtzman, C.; Robnett, C.J.J.O.C.M. Identification Of Clinically Important Ascomycetous Yeasts Based On Nucleotide Divergence In The 5′end Of The Large-Subunit (26s) Ribosomal Dna Gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- De Smet, I.; Van Hoorde, L.; Vande Woestyne, M.; Christiaens, H.; Verstraete, W.J.J.o.A.B. Significance of bile salt hydrolytic activities of lactobacilli. J. Appl. Bacteriol. 1995, 79, 292–301. [Google Scholar] [CrossRef]
- Fadahunsi, I.F.; Olubodun, S.J.B.O.T.N.R.C. Antagonistic Pattern Of Yeast Species Against Some Selected Food-Borne Pathogens. Bull. Natl. Res. Cent. 2021, 45, 1–19. [Google Scholar] [CrossRef]
- Auclair, E. Yeast As An Example Of The Mode Of Action Of Probiotics In Monogastric And Ruminant Species. 2001. pp. 45–53. Available online: https://om.ciheam.org/om/pdf/c54/01600010.pdf (accessed on 13 May 2021).
- Ferronato, G.; Prandini, A.J.A. Dietary Supplementation Of Inorganic, Organic, And Fatty Acids In Pig: A Review. Anaimals 2020, 10, 1740. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.; Kalam, A.; Sarker, M.; Li, T.; Yin, J. Probiotic Species In The Modulation Of Gut Microbiota: An Overview. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, T.S.; Gibson, G.R. Prebiotics. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.S.; Rabadzhiev, Y.; Eller, M.R.; Iliev, I.; Ivanova, I.; Santana, W.C. Microorganisms In Honey. Honey Anal. 2017, 500, 1–378. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.; Quek, S.-Y.; Gutierrez-Maddox, N.; Gao, Y.; Shu, Q.J.F.Q. Effect Of Honey In Improving The Gut Microbial Balance. Food Qual. Saf. 2017, 1, 107–115. [Google Scholar] [CrossRef]
- Cliver, D.; Snowdown, J.J.I.J.F.M. Microorganisms In Honey. Int. J. Food Microbiol. 1996, 31, 1–26. [Google Scholar] [CrossRef]
- Spor, A.; Kvitek, D.J.; Nidelet, T.; Martin, J.; Legrand, J.; Dillmann, C.; Bourgais, A.; De Vienne, D.; Sherlock, G.; Sicard, D.J.E. Phenotypic And Genotypic Convergences Are Influenced By Historical Contingency And Environment In Yeast. Evolution 2014, 68, 772–790. [Google Scholar] [CrossRef]
- Orfao, A. Phenotypic Analysis. Curr. Protoc. Cytom. 2021. [Google Scholar] [CrossRef]
- Offei, B.; Vandecruys, P.; De Graeve, S.; Foulquié-Moreno, M.R.; Thevelein, J.M.J.G.R. Unique Genetic Basis Of The Distinct Antibiotic Potency Of High Acetic Acid Production In The Probiotic Yeast Saccharomyces Cerevisiae Var. Boulardii. Genome Res. 2019, 29, 1478–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, C.; Meirinho, S.; Estevinho, M.; Choupina, A.J.A.D.Z. Yeast Species Associated With Honey: Different Identification Methods. Arch. Zootec. 2010, 59, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Wang, L.; Jin, Y.; Zhang, J.; Su, L.; Zhang, X.; Zhou, J.; Li, Y.J.F.I.M. The Microbial Community Dynamics During The Vitex Honey Ripening Process In The Honeycomb. Front. Microbiol. 2017, 8, 1649. [Google Scholar] [CrossRef] [PubMed]
- Gerardy, R.; Debecker, D.P.; Estager, J.; Luis, P.; Monbaliu, J.-C.M.J.C.R. Continuous Flow Upgrading Of Selected C2–C6 Platform Chemicals Derived From Biomass. Chem. Rev. 2020, 120, 7219–7347. [Google Scholar] [CrossRef]
- Cadete, R.M.; Melo-Cheab, M.A.; Viana, A.L.; Oliveira, E.S.; Fonseca, C.; Rosa, C.A.J.W.J.O.M. The Yeast Scheffersomyces Amazonensis Is An Efficient Xylitol Producer. World J. Microbiol. Biotechnol. 2016, 32, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, B.; Wang, D.; Gao, X.; Hong, J.J.B.T. Xylitol Production At High Temperature By Engineered Kluyveromyces Marxianus. Bioresour. Technol. 2014, 152. [Google Scholar] [CrossRef]
- Atzmüller, D.; Ullmann, N.; Zwirzitz, A.J.A.E. Identification Of Genes Involved In Xylose Metabolism Of Meyerozyma Guilliermondii And Their Genetic Engineering For Increased Xylitol Production. AMB Express 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Zou, Y.-Z.; Qi, K.; Chen, X.; Miao, X.-L.; Zhong, J.-J.J.J.O.B. Favorable Effect Of Very Low Initial Kla Value On Xylitol Production From Xylose By A Self-Isolated Strain Of Pichia Guilliermondii. J. Biosci. Bioeng. 2010, 109, 149–152. [Google Scholar] [CrossRef]
- Tokatlı, M.; Gülgör, G.; Bağder Elmacı, S.; Arslankoz İşleyen, N.; Özçelik, F. In Vitro Properties Of Potential Probiotic Indigenous Lactic Acid Bacteria Originating From Traditional Pickles. Biomed. Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef] [Green Version]
- Kast, C.; Roetschi, A.J.F.M. Evaluation Of Baker’s Yeast In Honey Using A Real-Time Pcr Assay. Food Microbiol. 2017, 62, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.R.; Lopes, A.C.A.; Pereira, R.A.; Cardoso, P.G.; Duarte, W.F.J.A.O.M. Selection Of Potentially Probiotic Kluyveromyces Lactis For The Fermentation Of Cheese Whey–Based Beverage. Ann. Microbiol. 2019, 69, 1361–1372. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.J.D. Finding Needles In Haystacks: Linking Scientific Names, Reference Specimens And Molecular Data For Fungi. Database 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, A.M.; Dlauchy, D.; Péter, G.J.I.J.O.S.; Microbiology, E. Meyerozyma Amylolytica Sp. Nov. From Temperate Deciduous Trees And The Transfer Of Five Candida Species To The Genus Meyerozyma. Int. J. Syst. Evol. Microbiol. 2017, 67, 3977–3981. [Google Scholar] [CrossRef]
- Chang, C.-F.; Lee, C.-F.; Liu, S.-M.J.A.V.L. Candida Neustonensis Sp. Nov., A Novel Ascomycetous Yeast Isolated From The Sea Surface Microlayer In Taiwan. Antonie Leeuwenhoek 2010, 97, 35–40. [Google Scholar] [CrossRef]
- Zacchi, L.; Vaughan-Martini, A.J.A.O.M. Yeasts Associated With Insects In Agricultural Areas Of Perugia, Italy. Ann. Microbiol. 2002, 52, 237–244. [Google Scholar]
- Vaughan-Martini, A.; Kurtzman, C.P.; Meyer, S.A.; O′neill, E.B.J.F.Y.R. Two New Species In The Pichia Guilliermondii Clade: Pichia Caribbica Sp. Nov., The Ascosporic State Of Candida Fermentati, And Candida Carpophila Comb. Nov. FEMS Yeast Res. 2005, 5, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.-V.; Boekhout, T.J.F.Y.R. Characterization Of Saccharomyces Uvarum (Beijerinck, 1898) And Related Hybrids: Assessment Of Molecular Markers That Predict The Parent And Hybrid Genomes And A Proposal To Name Yeast Hybrids. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; De Vries, M.; Verkleij, G.; Crous, P.; Boekhout, T.J.S.I.M. Dna Barcoding Analysis Of More Than 9 000 Yeast Isolates Contributes To Quantitative Thresholds For Yeast Species And Genera Delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Mccullough, M.J.; Clemons, K.V.; Mccusker, J.H.; Stevens, D.A.J.J.O.C.M. Species Identification And Virulence Attributes Of Saccharomyces Boulardii (Nom. Inval.). J. Clin. Microbiol. 1998, 36, 2613–2617. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, I.S.J.Y. Tailoring Wine Yeast For The New Millennium: Novel Approaches To The Ancient Art Of Winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Pedersen, L.L.; Owusu-Kwarteng, J.; Thorsen, L.; Jespersen, L.J.I.J.O.F.M. Biodiversity And Probiotic Potential Of Yeasts Isolated From Fura, A West African Spontaneously Fermented Cereal. Int. J. Food Microbiol. 2012, 159, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Fietto, J.L.; Araújo, R.S.; Valadão, F.N.; Fietto, L.G.; Brandão, R.L.; Neves, M.J.; Gomes, F.C.; Nicoli, J.R.; Castro, I.M.J.C.J.O.M. Molecular And Physiological Comparisons Between Saccharomyces Cerevisiae And Saccharomyces Boulardii. Can. J. Microbiol. 2004, 50, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotcheva, V.; Hristozova, E.; Hristozova, T.; Guo, M.; Roshkova, Z.; Angelov, A.J.F.B. Assessment Of Potential Probiotic Properties Of Lactic Acid Bacteria And Yeast Strains. Food Biotechnol. 2002, 16, 211–225. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; De Cadiñanos, L.P.G.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C.J.F.R.I. Adhesion Abilities Of Dairy Lactobacillus Plantarum Strains Showing An Aggregation Phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Golubev, W. Antagonistic Interactions Among Yeasts. In Biodiversity And Ecophysiology Of Yeasts; Springer: Berlin/Heidelberg, Germany, 2006; pp. 197–219. [Google Scholar] [CrossRef]
- Michaëlsson, K.; Wolk, A.; Langenskiöld, S.; Basu, S.; Lemming, E.W.; Melhus, H.; Byberg, L. Milk Intake And Risk Of Mortality And Fractures In Women And Men: Cohort Studies. BMJ 2014, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caggia, C.; Restuccia, C.; Pulvirenti, A.; Giudici, P. Identification Of Pichia Anomala Isolated From Yoghurt By Rflp Of The Its Region. Int. J. Food Microbiol. 2001, 71, 71–73. [Google Scholar] [CrossRef]
- Liu, J.-J.; Zhang, G.-C.; Kong, I.I.; Yun, E.J.; Zheng, J.-Q.; Kweon, D.-H.; Jin, Y.-S. A Mutation In Pgm2 Causing Inefficient Galactose Metabolism In The Probiotic Yeast Saccharomyces Boulardii. Appl. Environ. Microbiol. 2018, 84, 02858-17. [Google Scholar] [CrossRef] [Green Version]
- Cevrimli, B.S.; Kariptas, E.; Ciftci, H.J.A.J.O.C. Effects Of Fermentation Conditions On Citric Acid Production From Beet Molasses By Aspergillus Niger. Asian J. Chem. 2009, 21, 3211–3218. [Google Scholar]
- Hesham, A.E.-L.; Mostafa, Y.S.; Alsharqi, L.E.O.J.M. Optimization Of Citric Acid Production By Immobilized Cells Of Novel Yeast Isolates. Mycobiology 2020, 48, 122–132. [Google Scholar] [CrossRef]
- Alonso, S.; Rendueles, M.; Díaz, M.J.C.R.I.B. Microbial Production Of Specialty Organic Acids From Renewable And Waste Materials. Crit. Rev. Biotechnol. 2015, 35, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, L.; Isar, J.; Meghwanshi, G.; Saxena, R.J.J.O.A.M. A Cost Effective Fermentative Production Of Succinic Acid From Cane Molasses And Corn Steep Liquor By Escherichia Coli. J. Appl. Microbiol. 2006, 100, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Papon, N.; Savini, V.; Lanoue, A.; Simkin, A.J.; Crèche, J.; Giglioli-Guivarc’h, N.; Clastre, M.; Courdavault, V.; Sibirny, A.A.J.C.G. Candida Guilliermondii: Biotechnological Applications, Perspectives For Biological Control, Emerging Clinical Importance And Recent Advances In Genetics. Curr. Genet. 2013, 59, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Goodwin Tw, P.S. Studies On The Biosynthesis Of Riboflavin; Nitrogen Metabolism and Flavinogenesis in Eremothecium Ashbyii. Biochem. J. 1945, 57, 631. [Google Scholar] [CrossRef] [Green Version]
- Gong, F.; Sheng, J.; Chi, Z.; Li, J.J.J.O.I.M. Inulinase Production By A Marine Yeast Pichia Guilliermondii And Inulin Hydrolysis By The Crude Inulinase. J. Ind. Microbiol. Biotechnol. 2007, 34, 179–185. [Google Scholar] [CrossRef]
- Wang, G.-Y.; Chi, Z.; Song, B.; Wang, Z.-P.; Chi, Z.-M.J.B.T. High Level Lipid Production By A Novel Inulinase-Producing Yeast Pichia Guilliermondii Pcla22. Bioresour. Technol. 2012, 124, 77–82. [Google Scholar] [CrossRef]
- Carneiro, C.V.G.; Almeida, J.R.J.M. Xylitol Production: Identification And Comparison Of New Producing Yeasts. Microorganisms 2019, 7, 484. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-L.; Liao, Y.-Y.; Zhang, J.; Wang, X.-L.J.B. Comparative Transcriptome Analysis Of Salt Tolerance Mechanism Of Meyerozyma Guilliermondii W2 Under Nacl Stress. 3 Biotech. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Buck, J.D.; Cleverdon, R.C.J.L. The spread plate as a method for the enumeration of marine bacteria. Limnol. Oceanogr. 1960, 5, 78–80. [Google Scholar] [CrossRef]
- Muir, A.; Harrison, E.; Wheals, A.J.F.Y.R. A Multiplex Set Of Species-Specific Primers For Rapid Identification Of Members Of The Genus Saccharomyces. FEMS Yeast Res. 2011, 11, 552–563. [Google Scholar] [CrossRef]
- Kanbe, T.; Horii, T.; Arishima, T.; Ozeki, M.; Kikuchi, A.J.Y. Pcr-Based Identification Of Pathogenic Candida Species Using Primer Mixes Specific To Candida Dna Topoisomerase Ii Genes. Yeast 2002, 19, 973–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciardo, D.E.; Lucke, K.; Imhof, A.; Bloemberg, G.V.; Böttger, E.C.J.J.O.C.M. Systematic Internal Transcribed Spacer Sequence Analysis For Identification Of Clinical Mold Isolates In Diagnostic Mycology: A 5-Year Study. J. Clin. Microbiol. 2010, 48, 2809–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryce, T.; Palladino, S.; Kay, I.; Coombs, G.J.M.M. Rapid Identification Of Fungi By Sequencing The Its1 And Its2 Regions Using An Automated Capillary Electrophoresis System. Med. Mycol. 2003, 41, 369–381. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.J.P.P.A.G.T.M. Amplification And Direct Sequencing Of Fungal Ribosomal Rna Genes For Phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K.J.M.B. Mega7: Molecular Evolutionary Genetics Analysis Version 7.0 For Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Rodríguez, A.M.; Carrascosa, A.V.; Requena, T.J.L.-F.S. Yeasts in foods and beverages: In vitro characterisation of probiotic traits. Food Sci. Technol. 2015, 64, 1156–1162. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Isolate Name | Temp. 30 °C | Temp. 37 °C | Temp. 42 °C | PH 2.5 | PH 2.0 | 0.3% Bile Salt | Auto- Aggregation Ability (%) | Galactose Utilization Ability | Genus/Species Identification |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | RSO1 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 100 a | ✓ | Saccharomyces cerevisiae |
| 2 | RSO2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 100 a | ✓ | Saccharomyces cerevisiae |
| 3 | RSO3 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 100 a | ✓ | Saccharomyces cerevisiae |
| 4 | RSO4 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 98 a | ✓ | Saccharomyces cerevisiae |
| 5 | KTF1 | ✓ | ✓ | - | ✓ | ✓ | ✓ | 80 b | ✓ | Non- Saccharomyces |
| 6 | KTF2 | ✓ | ✓ | - | ✓ | ✓ | ✓ | 86 b | ✓ | Non- Saccharomyces |
| 7 | KTF3 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 91 a | ✓ | Meyerozyma guilliermondii |
| 8 | KTF5 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 95 a | ✓ | Meyerozyma guilliermondii |
| 9 | KTF6 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 73 c | ✓ | Non- Saccharomyces |
| 10 | KTF16 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 78 c | ✓ | Non- Saccharomyces, Meyerozyma |
| 11 | KTF18 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 72 c | ✓ | Meyerozyma guilliermondii |
| 12 | RBF19 | ✓ | ✓ | - | ✓ | ✓ | ✓ | 89 b | ✓ | Non- Saccharomyces |
| 13 | RBF20 | ✓ | ✓ | - | ✓ | ✓ | ✓ | 68 d | ✓ | Meyerozyma guilliermondii |
| 14 | RBF27 | ✓ | ✓ | - | ✓ | ✓ | ✓ | 81 b | ✓ | Non- Saccharomyces |
| 15 | RBF37 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 52 d | ✓ | Meyerozyma guilliermondii |
| 16 | RBF47 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 70 c | ✓ | Meyerozyma guilliermondii |
| 17 | RBF56 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 88 b | ✓ | Meyerozyma guilliermondii |
| 18 | BY4742 | ✓ | - | - | ✓ | ✓ | ✓ | 100 a | ✓ | Saccharomyces cerevisiae |
| 19 | SB | ✓ | ✓ | - | ✓ | ✓ | ✓ | 96 a | - | Saccharomyces var. boulardii |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahoor, F.; Sooklim, C.; Songdech, P.; Duangpakdee, O.; Soontorngun, N. Selection of Potential Yeast Probiotics and a Cell Factory for Xylitol or Acid Production from Honeybee Samples. Metabolites 2021, 11, 312. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050312
Zahoor F, Sooklim C, Songdech P, Duangpakdee O, Soontorngun N. Selection of Potential Yeast Probiotics and a Cell Factory for Xylitol or Acid Production from Honeybee Samples. Metabolites. 2021; 11(5):312. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050312
Chicago/Turabian StyleZahoor, Farah, Chayaphathra Sooklim, Pattanan Songdech, Orawan Duangpakdee, and Nitnipa Soontorngun. 2021. "Selection of Potential Yeast Probiotics and a Cell Factory for Xylitol or Acid Production from Honeybee Samples" Metabolites 11, no. 5: 312. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050312