
Exploring the Gut Microbiota and Cardiovascular Disease
by
 , and
, and
Kiera Murphy
1,2 ,
,
Aoife N. O’Donovan
1,2,3,
Noel M. Caplice
2,4,
R. Paul Ross
2 and
Catherine Stanton
1,2,3,* 1
Teagasc Food Research Centre, Moorepark, Co. Cork P61 C996, Ireland
2
APC Microbiome Ireland, Biosciences Institute, University College Cork, Cork T12 YT20, Ireland
3
VistaMilk SFI Research Centre, Teagasc, Moorepark, Co. Cork P61 C996, Ireland
4
Centre for Research in Vascular Biology, Biosciences Institute, University College Cork, Cork T12 YT20, Ireland
*
Author to whom correspondence should be addressed.
Metabolites 2021, 11(8), 493; https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080493
Submission received: 27 May 2021
/
Revised: 24 July 2021
/
Accepted: 27 July 2021
/
Published: 29 July 2021
(This article belongs to the Special Issue Nutrition, Microbiota and Metabolism)
Abstract
:Cardiovascular disease (CVD) has been classified as one of the leading causes of morbidity and mortality worldwide. CVD risk factors include smoking, hypertension, dyslipidaemia, obesity, inflammation and diabetes. The gut microbiota can influence human health through multiple interactions and community changes are associated with the development and progression of numerous disease states, including CVD. The gut microbiota are involved in the production of several metabolites, such as short-chain fatty acids (SCFAs), bile acids and trimethylamine-N-oxide (TMAO). These products of microbial metabolism are important modulatory factors and have been associated with an increased risk of CVD. Due to its association with CVD development, the gut microbiota has emerged as a target for therapeutic approaches. In this review, we summarise the current knowledge on the role of the gut microbiome in CVD development, and associated microbial communities, functions, and metabolic profiles. We also discuss CVD therapeutic interventions that target the gut microbiota such as probiotics and faecal microbiota transplantation.
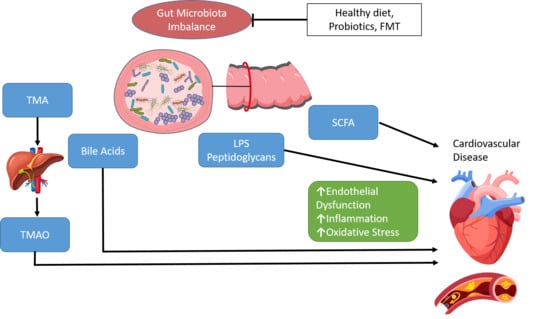
1. Introduction
Cardiovascular disease (CVD) is a general term for a number of pathologies including coronary heart disease (CHD), cerebrovascular disease, peripheral artery disease, congenital and rheumatic heart disease and venous thromboembolism [1]. CVD is a chronic progressive condition, often leading to irreversible damage to vascular structures in the form of atherosclerosis and detrimental clinical outcomes such as arterial thrombosis, myocardial infarction (MI) and stroke. Elevated levels of serum cholesterol (hypercholesterolemia), particularly low-density lipoprotein cholesterol (LDL-C) is a well-documented risk factor for CVD. LDL-C deposits cholesterol into artery walls and this plaque build-up can lead to atherosclerosis [2]. CVD has been classified as one of the leading causes of morbidity and mortality worldwide, which has led to extensive health and economic burdens accounting for approximately one in every three deaths in the United States and one in every four deaths in Europe [3].
The World Health Organisation (WHO) has estimated that over 75% of premature CVD is avoidable and that risk factor improvement can aid in the reduction in the growing incidence of CVD [4]. The INTERHEART study established a case–control study of acute MI from 52 countries worldwide. The study identified nine easily assessed risk factors which were highly and significantly associated with an increased risk of acute MI [5]. Smoking, dyslipidaemia, hypertension, diabetes, abdominal obesity, psychosocial factors, daily fruits and vegetables, exercise, alcohol intake and apolipoprotein (Apo)B/ApoA1 ratio accounted for over 90% of the risk of acute MI.
The human body harbours trillions of microbial cells that form a complex microbial community in the intestine known as the gut microbiota. The gut is predominantly inhabited by two phyla, Bacteriodetes and Firmicutes, which account for 90% of the total inhabiting microbes. The remaining 10% consists of Actinobacteria, Cyanobacteria, Fusobacteria, Proteobacteria and Verrucomicrobia [6,7]. Beginning at birth, there are multiple factors that can influence how the gut is colonised, such as mode of birth (vaginal vs. caesarean), feeding method (breast feeding vs. infant formula), exposure to antibiotics, hygiene standards and geographical location [8,9]. It has been shown that the gut microbiota of infants who are vaginally delivered predominantly consist of Bifidobacterium in the weeks and months after birth, whereas those infants born via caesarean section predominantly consist initially of maternal skin microbiota, mainly Staphylococcus [10]. As the infant ages, the initial dominant aerobic gut environment matures and evolves to form an anaerobic environment, resulting in greater abundances of Bifidobacterium and Clostridia [7]. The gut microbiota begin to stabilise and evolve toward an adult-like composition by the second year of life [10].
The gut microbiota perform essential metabolic functions, serve as a source of essential nutrients and vitamins and assist in energy and nutrient extraction from food [11,12]. The gut microbiota also work in conjunction with the host’s defence and immune systems to protect against pathogen infiltration and colonisation. An imbalance in gut microbial composition can result in these mechanisms becoming disrupted and can contribute to increased intestinal permeability or a ‘leaky gut’ [13]. In the leaky gut, the tight junctions which support the epithelial lining barrier are disrupted, allowing infiltration of microbes, toxins or antigens into the tissues beneath. This may trigger inflammation through the activation of local and systemic immune responses. Shifts in the composition of the gut microbiota disrupt homeostasis and have been associated with long-term consequences, leading to disorders including CVD. 16S rRNA gene sequencing and whole-metagenome shotgun sequencing have added to the growing body of evidence that suggests that alterations of the gut microbiome composition affect the pathogenesis of CVD [7,14,15,16]. In addition to the composition of the gut microbiota, metabolites produced by these microbes can enter into the circulation and act as a modifiers of gut microbial effects on the host [17,18]. A number of microbiome studies have shown a potential link between microbial metabolites including short-chain fatty acids (SCFAs), bile acids (BA) and trimethylamine N-oxide (TMAO) and CVD [19,20,21].
The gut microbiota are involved in the digestion of food through two catabolic pathways, the saccharolytic pathway and the proteolytic pathway [22]. The saccharolytic pathway uses the gut microbiota to help break down sugars in the ingested foods, leading to SCFA production. The proteolytic pathway is characterised by the fermentation of ingested proteins, leading to SCFA production as well as formation of other metabolites including ammonia, thiols, amines, phenols and indoles. Diet is an important risk factor for CVD and the complex interactions between food and gut microbiota, and resulting metabolites play a key role in cardiovascular health. Given the growing body of evidence linking the gut microbiota with development of CVD, therapies aimed at modulating the gut microbiota have been suggested among the most promising strategies to prevent CVD.
2. Gut Microbiota and CVD
Many metabolites in the human body originate from intestinal microbes and there is growing evidence supporting the relationship between these metabolites and the development of CVD [18,23]. Through these metabolites, the gut microbiota influence host metabolic pathways such as cholesterol metabolism, in addition to inflammatory reactions and oxidative stress. Gut microbe-dependent metabolites may be absorbed into the bloodstream through the intestinal epithelium, which in turn influences the function of several organs and bodily systems [24]. Production of TMAO by the gut microbiota is a key mechanism of CVD. Dietary choline, phosphatidylcholine and carnitine are metabolised by gut microbes to produce trimethylamine (TMA). TMA is transported to the liver and further converted to TMAO via hepatic enzymes, flavin-containing monooxygenases [25,26]. TMAO has been shown to modulate cholesterol and sterol metabolism, cholesterol transport and bile acids levels [19,20,21,27]. Elevated serum levels of TMAO are associated with early atherosclerosis, severity of peripheral artery disease and with high risk of CVD mortality [28,29]. In a study of 4007 patients undergoing elective coronary angiography, increased plasma levels of TMAO were associated with an increased risk of death, MI, and stroke during three years of follow-up [21]. Dietary supplementation of TMAO to mice reduced the expression of Cyp7a1, the main bile acid synthetic enzyme and rate-limiting step in the catabolism of cholesterol, resulting in significantly reduced cholesterol transport [19]. In a study by Wang et al., associations were observed between CAD, peripheral vascular disease and MI and the gut metabolites choline, TMAO and betaine [20]. A TMAO-enriched diet expanded the atherosclerotic lesion area in atherosclerosis-prone mice (C57BL/6J Apoe−/−), while suppression of gut microbiota in atherosclerosis-prone mice inhibited dietary-choline-enhanced atherosclerosis. More recently, a murine study showed that both healthy and unhealthy diets can increase plasma levels of TMAO, with the gut itself being a significant site for oxidative production of TMAO [30]. Importantly this study reconciles some contradictory data on TMAO by showing no direct association between plasma TMAO and atherosclerosis extent but a clear association between TMAO levels and atherosclerotic plaque instability. This suggests that TMAO is a marker of cardiovascular risk.
SCFAs are the main metabolites produced by the gut microbiota through the fermentation of indigestible complex polysaccharides and proteins in the large intestine. The most abundant SCFAs are acetate (C2), propionate (C3), and butyrate (C4). SCFAs play important roles in gut health through maintenance of intestinal epithelial barrier integrity, reduction in inflammation, mucus production, and directly affecting immune cells. SCFAs bind to G protein-coupled receptors, the best-studied of which are free fatty acid receptors FFAR2 and FFAR3. FFAR2 is an essential receptor for neutrophil recruitment and through binding with FFAR2, SCFAs can help mediate inflammation [31]. SCFAs can induce satiety by stimulating the secretion of the gut hormones glucagon-like peptide-1 (GLP-1) and peptide YY (PYY) [32,33]. SCFAs can also influence blood pressure (BP) regulation by binding with FFAR2, FFAR3 and olfactory receptor 78 (OLFR78) [34]. Stimulation of OLFR78 has been demonstrated to elevate BP, while stimulation of FFAR3 lowered BP [35]. SCFAs regulate cholesterol distribution in liver and blood by inhibiting liver adipose synthetase activity. As SCFAs play a role in reducing serum cholesterol levels, decreases in SCFA production have been associated with dyslipidaemia. Concentrations of SFCAs have also been observed to be lower in patients with atherosclerotic vascular disease or hypertension [36].
The conversion of primary BAs to secondary BAs is also dependent on the gut microbiota. The gut microbiota produce bile salt hydrolase which catalyses the deconjugation of conjugated BAs and the release of free BAs and amino acids. A high intake of saturated fat leads to increased BA secretion and the presence of excess BAs in the intestine. Primary BAs are converted to hydrophobic secondary BAs, such as deoxycholic acid (DCA). Secondary BAs have been linked with increased hydrophobicity and enhanced membrane lipid binding and may cause changes in gut microbiota composition and structure [37,38]. Secondary BAs that enter the circulation can act as hormones and regulate signalling pathways including metabolism, inflammation and energy expenditure. Bile acid secretion disorders can cause blood lipid abnormalities and have been implicated in the development of atherosclerosis and other cardiometabolic diseases [3,38].
Components from the gut microbiota such as lipopolysaccharide (LPS) and peptidoglycan can trigger numerous immunomodulatory functions. LPS, a component of the Gram-negative bacterial cell membrane, is an endotoxin which promotes chronic systemic inflammation. LPS induces expression of inflammatory mediators and activates an immune response through Toll-like receptors (TLRs) and the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-ĸB) pathway [39]. LPS may enter the circulation via intestinal absorption leading to endotoxemia, elevated levels of endotoxins in the blood. Endotoxemia has been associated with the pathogenesis of several disorders including atherosclerosis, obesity and insulin resistance [40]. In particular, endotoxemia has been shown to be associated with an increased risk of developing atherosclerosis in smokers and subjects with infectious disease [41].
The gut microbiota may also influence CVD development via oxidative stress. High oxygen levels lead to the formation of reactive oxygen species (ROS) including superoxide anion radicals, hydroxyl radicals, and hydrogen peroxide. During oxidative stress, ROS cause free-radical chain reactions, resulting in damage to proteins, DNA, and lipids, and even cell death [42]. Oxidative stress has been linked to several CVD risk factors including obesity, hypertension dyslipidaemia and diabetes. Endothelial dysfunction is a key hallmark underlying CVD and nitric oxide (NO) is the major factor accounting for endothelium-dependent relaxation in the aorta [43]. Increased ROS bioavailability reduces NO bioavailability and promotes inflammation, thereby affecting endothelial function [43]. Diets rich in foods with a high content of antioxidants and polyphenols have been shown to decrease oxidative stress markers. Elevated levels of glucose lead to an increase in oxidative stress, resulting in the oxidation of lipoproteins and production of adhesion molecules [44]. Gut microbes can influence oxidative stress through the regulation of uric acid metabolism. Elevated plasma uric acid levels may lead to an increase in the production of oxygen free radicals and promote oxidative stress and endothelial dysfunction [45].
3. Gut Microbiota Composition in Cardiovascular Disease
Next-generation sequencing (NGS) technology has greatly enhanced our understanding of the human gut microbiome. In particular, 16S rRNA gene sequencing has been extensively employed to profile the gut microbiota composition. The 16S rRNA gene is highly conserved between bacteria and contains nine hypervariable regions with bacterial species-specific variations. Alterations in the intestinal microbiota composition are believed to contribute to the pathogenesis of many diseases including CVD. Table 1 summarises the main findings of studies examining the relationship between the gut microbiota and CVD. Kelly et al. examined the association between the gut microbiota and lifetime CVD risk [46]. Prevotella 2, Prevotella 7, Tyzzerella and Tyzzerella 4 were found to be enriched in subjects with high CVD risk compared with low risk subjects. By contrast, Alloprevotella and Catenibacterium were depleted among subjects with high versus low CVD risk. Alpha diversity was inversely associated with lifetime CVD risk.
The gut microbiota have been suggested to be an important contributor to the development of obesity, a key risk factor for CVD. Bäckhed et al. initially proposed that the gut microbiota was an important environmental factor that affects predisposition towards energy storage and fat mass gain [47]. Conventional mice were observed to have a higher body fat content and higher metabolic rate than germ-free (GF) counterparts. The same group sought to examine the mechanisms underlying the protection against obesity in GF mice [48]. Here, they demonstrated that in contrast to conventional mice, GF mice were resistant to diet-induced obesity, even if the mice ingested the same amount of calories. The lean phenotype of GF mice was associated with mechanisms that result in increased fatty acid metabolism. Ley et al. reported a reduced abundance of Bacteroidetes and a proportionally increased abundance of Firmicutes in obese subjects [49]. An increased Bacteroidetes/Firmicutes ratio has been replicated in numerous studies analysing the microbiota composition in obese subjects [50,51,52,53]. However, in contrast to these findings, there have been many studies which have not observed an increase in the Bacteroidetes/Firmicutes ratio and in some cases have reported a decreased Bacteroidetes/Firmicutes ratio [54,55,56,57,58,59,60]. Obese subjects have also been shown to present with reduced microbial diversity compared with lean controls [61,62,63]. Several studies have reported that Akkermansia muciniphila, a mucin-degrading bacterium, is inversely correlated with obesity [64,65,66,67,68]. Ozato et al. reported an inverse association between the genus Blautia and visceral fat accumulation in Japanese adults [69]. Indeed, Blautia was the only genus significantly and inversely associated with visceral fat accumulation. A 2020 study reported a positive association between Acidaminococcus and body mass index (BMI), as well as waist and hip circumferences in Bangladeshi adults [70]. Companys et al. demonstrated that anthropometric parameters were positively associated with Collinsella aerofaciens, Dorea formicigenerans and Dorea longicatena, which had higher abundances in overweight/obese subjects [71]. Saturated fatty acids were negatively associated with the Intestinimonas, a biomarker of the overweight/obese subjects. By contrast, saturated fatty acids were positively associated with Roseburia, a biomarker for the lean subjects.
The gut microbiota also have the capacity to contribute to dyslipidaemia. Martinez-Guryn et al. showed that GF mice exhibit decreased plasma triglyceride and LDL-C compared to control mice [72]. Karlsson et al. reported a positive correlation between high-density lipoprotein cholesterol (HDL-C) and Clostridium species and a negative correlation between serum triglycerides and Clostridum species [73]. Rebolledo et al. assessed differences in gut microbial composition between hypercholesterolemic subjects and controls [74]. Hypercholesterolemic subjects harboured a lower richness and diversity of bacterial communities than controls. Fu et al. examined the gut microbiota composition in 893 subjects from the LifeLines-DEEP population cohort, a Dutch population-based study [75]. Thirty-four bacterial taxa were associated with BMI and blood lipids. In particular, Eggerthella was associated with increased triglyceride and decreased HDL-C, and family Pasteurellaceae with decreased triglyceride. The authors reported that the microbiome had little effect on LDL-C or total cholesterol (TC). A more recent 2021 study showed that genera of the Lachnospiraceae family are negatively associated with lipid CVD risk factors including body fat, LDL-C and TC [71]. Liang et al. demonstrated that a high-fat and high-cholesterol diet (HFHCD) significantly decreased the alpha diversity of the gut microbiota in mice compared with high-fat diet (HFD) alone [76]. In addition, HFHCD mice had reduced abundances of Oscillospira, Odoribacter, Bacteroides, and Prevotella compared with HFD and greater Ruminococcus and Akkermansia. In 2021, Zhang et al. reported that mice fed a HFHCD had increased Mucispirillum, Desulfovibrio, Anaerotruncus and Desulfovibrionaceae, while Bacteroides and Bifidobacterium were depleted [77]. This finding was further corroborated in human hypercholesteremia patients.
With regard to CAD, Emoto et al. reported that the phylum Bacteroidetes was decreased and the order Lactobacillales was increased in CAD patients compared with controls [78]. Mitra et al. demonstrated differences in microbial composition between symptomatic and asymptomatic atherosclerotic plaques [79]. Asymptomatic atherosclerotic plaques contained increased abundances of Porphyromonadaceae, Bacteroidaceae, Micrococcacaea, and Streptococcacaea. In contrast, symptomatic plaques had an increased abundance of Helicobacteracaea, Neisseriaceae, and Thiotrichacaea. However, it remains unclear whether these bacteria are active or passive participants in plaque erosion/rupture. Collinsella was found to be enriched in patients with symptomatic atherosclerosis, while Roseburia and Eubacterium were enriched in healthy controls [80]. More recently, in a study by Zhu et al., decreased diversity and richness were observed in CAD patients [81]. Escherichia-Shigella were significantly enriched, while Faecalibacterium, Subdoligranulum, Roseburia and Eubacterium rectale were significantly reduced in CAD patients. Jie et al. showed an increase in the relative abundances of Escherichia coli, Enterobacter aerogenes, Klebsiella and Streptococcus in faecal samples from patients with atherosclerotic CVD than in healthy controls [82]. The relative abundances of bacteria typically associated with the oral cavity, such as Streptococcus, Lactobacillus salivarius, Solobacterium moorei, and Atopobium parvulum, were also higher in atherosclerotic patients than in healthy controls. Periodontal disease has been associated with atherosclerosis, suggesting that oral pathogens may contribute to the pathogenesis of atherosclerosis [83]. Koren et al. compared the microbial composition of oral, gut, and atherosclerotic plaque samples in patients with atherosclerosis [84]. Chryseomonas was detected in all atherosclerotic plaque samples. A correlation was noted between the combined abundances of Veillonella and Streptococcus in atherosclerotic plaques and in samples from the oral cavity. In addition, several OTUs were shared between the atherosclerotic plaque and oral or gut samples within the same individual. Hyvarinen et al. investigated the association between CAD and salivary levels of four major periodontal pathogens, Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, and Tannerella forsythia [85]. High salivary levels of A. actinomycetemcomitans were associated with an increased risk for both acute and stable CAD.
A number of studies have investigated whether changes in intestinal microbiota composition affect hypertension. Moghadamrad et al. demonstrated that GF mice presented with significantly lower BP when compared with conventional mice, suggesting a contribution of the intestinal microbiota to the regulation of hypertension [86]. An increased Firmicutes/Bacteroidetes ratio in hypertensive animal models and patients has been widely reported [87,88]. Callejo et al. analysed the faecal microbiota composition in a rat model of pulmonary arterial hypertension (PAH) using 16S rRNA sequencing [89]. No differences in alpha and beta diversities were observed. However, PAH rats had a significantly increased Firmicutes/Bacteroidetes ratio. A 2015 study examined the gut microbiota composition in both animal and human hypertension [87]. Microbial richness, diversity, and evenness were observed to be significantly decreased in the spontaneously hypertensive rat (SHR) and in a chronic angiotensin II infusion rat model. An increased Firmicutes/Bacteroidetes ratio and decreases in acetate- and butyrate-producing bacteria were also observed. The microbiota of the human hypertensive subjects was observed to have reduced diversity compared with that of control subjects. Li et al. found decreased microbial richness and diversity, and an increase in Prevotella and Klebsiella in both pre-hypertensive and hypertensive populations compared to healthy controls [90]. A 2020 study used a multivariable-adjusted model to examine the association between gut microbiota composition and BP in 6953 subjects in Finland [91]. The associations between overall microbial composition and BP were reported as weak. However, mainly positive significant associations between BP indexes and 45 genera (27 belonging to the phylum Firmicutes) were observed. Strong negative associations between 19 Lactobacillus species and BP were also found. In particular, Lactobacillus paracasei, a well-studied probiotic, was associated with lower mean arterial pressure and lower dietary sodium intake.
4. Gut Microbiota Function in Cardiovascular Disease
In contrast to 16S compositional analysis, metagenomic approaches facilitate the comprehensive analysis of microbial community functional potential as well as strain-level analysis. The gut microbiota are believed to encode at least 150-fold more genes than the human genome [92,93]. Metabolomic analysis has become an important complementary approach in microbiome studies, allowing researchers to directly measure the metabolites present in the intestine. Many metabolites in the human body originate from intestinal microbes and there is growing evidence supporting the relationship between these metabolites and the development of CVD [18,23].
A 2021 study identified high-fat diet (HFD)-induced alterations of 19 metabolites in the liver and 43 metabolites in cecal contents of mice as potential biomarkers related to obesity [94]. HFD was associated with increased lipid profiles and total bile acid. Hong et al. studied the gut functional and metabolite profile in C57BL/6J mice that were fed with chow or HFD with or without Astragalus polysaccharides supplementation (APS) [95]. HFD was associated with an increase in six bacterial pathways, while eight pathways were reduced. These included tryptophan metabolism and methane metabolism, which were significantly enriched by HFD and reversed by APS supplementation. In addition, the nicotinate and nicotinamide metabolism pathway was reduced by HFD intake and increased by APS. Purine metabolism and glutathione metabolism were reduced and increased by HFD, respectively. Twenty species, including Streptococcus equi and Bizionia argentinensis, were negatively correlated with glutamic acid and pyroglutamic acid. Medina et al. analysed microbial functional patterns in obese and lean people from six different regions and observed that microbial contribution to functional pathways were region specific [96]. In addition, the authors analysed functional changes in the gut microbiota of obese patients after bariatric surgery and reported that changes were specific to the type of obesity treatment received. In 2019, Tran et al. used a metaproteomic approach to identify microbial factors that may correlate to diet induced obesity in mice prior to and following administration of an obesogenic high-fat low-fibre diet for 8 weeks [97]. Large shifts in the overall structure of the metaproteome were observed following HFD, which appeared to be driven by proteins derived from Clostridiales and Bacteroidales. Functionally a decreased abundance of flagellin proteins was observed following HFD.
Karlsson et al. utilised shotgun sequencing to characterise the functional capacity in patients with symptomatic atherosclerosis. The gut microbiome of symptomatic atherosclerosis was enriched with proinflammatory peptidoglycan synthesis encoding genes and showed reduced production of anti-inflammatory β-carotenes compared with healthy controls [80]. Jie et al. performed a metagenome-wide association study on 218 patients with atherosclerotic CVD and 187 healthy controls [82]. The atherosclerotic patients’ faecal metagenome differed from that of the controls in the potential for transport or metabolism of several molecules associated with cardiovascular health. Genes required for the formation of TMA and the O-antigen of LPS were enriched in atherosclerotic samples. Zhu et al. utilised PICRUSt to predict bacterial functional potential based on 16S gene sequencing profiles from 70 patients with CAD and 98 healthy controls [81]. Genes related to the phosphotransferase system, propanoate metabolism, LPS biosynthesis, and amino acid metabolism were increased in CAD patients, indicative of the inflammation observed in CAD. Feng et al. utilised both metagenomic and metabolomic approaches to identify microbial metabolites associated with CHD from plasma and urine of CHD patients and healthy controls [98]. Several metabolites were identified as novel candidate biomarkers, including GlcNAc-6-P, mannitol and 15 plasma cholines. Spearman correlation analysis of those biomarkers with microbial species showed that GlcNAc-6-P and mannitol were positively associated with Clostridium sp. HGF2, Streptococcus sp. M334 and Streptococcus sp. M143. Zhou et al. characterised the blood microbiome in a cohort of 199 subjects with CHD, ST-segment elevation myocardial infarction (STEMI) or healthy controls using shotgun sequencing [99]. In STEMI patients, higher microbial richness and diversity were observed. Lactobacillus, Bacteroides, and Streptococcus were found in more than 12% of post-STEMI blood samples. In comparison to control and CHD patients, STEMI patients’ LPS and d-lactate levels increased and correlated with systemic inflammation and predicted adverse cardiovascular events.
Kim et al. performed shotgun metagenomics on faecal samples from patients with high BP and healthy controls [100]. In particular, altered butyrate production in patients with high BP was observed. Significant increases in plasma biomarkers of gut epithelial barrier dysfunction and altered immune status including intestinal fatty acid-binding protein (I-FABP), LPS, and augmented gut-targeting proinflammatory T helper 17 (Th17) were also observed in high BP patients. In addition, Zonulin, a gut epithelial tight junction protein regulator, was elevated in high BP patients and strongly correlated with systolic blood pressure (SBP). A 2017 metagenomic and metabolomic analyses of 56 subjects with pre-hypertension, 99 individuals with primary hypertension and 41 healthy controls, reported microbial functional alterations in the gut microbiota of both pre-hypertensive and hypertensive patients [90]. The metabolic functions and metabolites identified in pre-hypertensive and hypertensive adults were closely linked to inflammation. Wang et al. characterised microbial composition and function in two subtypes of hypertension; isolated systolic hypertension (ISH) and isolated diastolic hypertension (IDH) [101]. A reduction in bacterial richness and evenness was reported in IDH patients compared with ISH and healthy controls. Patients with IDH or ISH had increased abundances of Rothia mucilaginosa and reduced Clostridium sp. ASBs410 compared to controls. Functionally, KEGG modules such as sodium transport system were significantly decreased in IDH patients compared with control. In ISH patients, functions related to biotin biosynthesis were decreased. A recent 2021 study used whole-metagenome shotgun sequencing to analyse stool samples from patients diagnosed with both depression and hypertension (DEP-HTN) [102]. The endotype of DEP-HTN patients was distinctive from DEP or HTN alone and defined by co-occurrence of Eubacterium siraeum, Alistipes obesi, Holdemania filiformis, and Lachnospiraceae bacterium 1.1.57FAA with Streptococcus salivariu. Metagenomes of DEP-HTN patients engaged pathways that degrade beneficial SCFAs and gamma amino butyric acid (GABA) and were associated with enhanced inflammasome activities and sodium absorption.
Hayashi et al. reported an increase in TMAO levels in patients with both compensated and decompensated heart failure in addition to an altered gut microbiota composition [103]. In 2021, the same researchers used metagenome-wide shotgun sequencing to elucidate which of the key genes of the TMA synthesis pathways contributes to the increase in TMAO levels [104]. The abundance of cntA/B, derived mainly from Escherichia and Klebsiella was positively correlated with TMAO, in heart failure patients. No correlation was observed for cutC/D or betaine reductase. Cui et al. performed metagenomics and metabolomic analyses of faecal and plasma samples from patients with chronic heart failure (CHF) and controls [105]. A decreased abundance of Faecalibacterium prausnitzii and an increase in Ruminococcus gnavus were observed in CHF patients. Microbial genes involved in the metabolism of protective metabolites such as butyrate were significantly reduced in the metagenome of CHF patients. Meanwhile, genes for potentially harmful metabolites such as TMAO were significantly upregulated in CHF patients.
5. Gut Microbiota as Therapeutic Strategies for Cardiovascular Disease
5.1. Probiotics
Probiotics are defined by the FAO/WHO as “live microorganisms which, when administered in adequate amounts, confer a health benefit on the host” [106]. A growing body of evidence demonstrates the beneficial health impacts of probiotic supplementation and they could be considered as a potential therapeutic tool for CVD. Probiotics have been investigated in several studies for their use as anti-obesity treatments. Pedret et al. examined the effect of Bifidobacterium animalis subsp. lactis CECT 8145 (Ba8145) on 135 obese subjects. Ba8145 decreased BMI when compared with a baseline and placebo group and was associated with an increase in Akkermansia. A three-week RCT involving 25 obese hypertensive participants investigated if a hypocaloric diet (1500 kcals/day) supplemented with a probiotic cheese containing the Lactiplantibacillus plantarum strain TENSIA could reduce symptoms of metabolic syndrome (MetS) [107]. BMI was reported to be significantly reduced in the participants that received probiotic cheese versus the control group. A 12 week RCT randomised 210 healthy Japanese adults with large amounts of visceral fat to receive fermented milk containing Lactobacillus gasseri SBT2055 (LG2055) [108]. Participants who received LG2055 exhibited a significant lowering in abdominal visceral fat, waist and hip circumference, BMI and body fat mass when compared with those in the control group.
Another RCT investigated the impact of daily supplementation of Lactobacillus rhamnosus CGMCC1.3724 (LPR) with oligofructose and inulin for 24 weeks in 125 obese adults combined with an energy-restricted diet for the first 12 weeks [109]. LPR supplementation did not significantly affect weight loss compared to placebo. However, a significant reduction in body weight was observed in the female participants as a result of dietary intervention compared with the placebo. Cai et al. reported that L. plantarum FRT10 supplementation alleviated HFD-induced obesity in mice partly related to the activation of the peroxisome proliferator-activated receptor-α (PPARα)/carnitine palmitoyltransferase-1α (CPT1α) pathway [110]. FRT10 reduced fat weight, body weight gain, liver triacylglycerols and alanine aminotransferase concentrations. Supplementation with FRT10 was also associated with significantly increased abundances of Butyricicoccus, Butyricimonas, Intestinimonas, Odoribacter, and Alistipes, and decreased abundances of Desulfovibrionaceae, Roseburia, and Lachnoclostridium. Depommier et al. conducted a randomised controlled pilot study in 32 overweight/obese insulin-resistant individuals [111]. Daily supplementation of A. muciniphila significantly improved insulin sensitivity and reduced insulinemia and plasma TC compared to placebo.
Probiotic bacteria have been studied as a therapeutic to improve lipid profiles. Probiotic bacteria with bile salt hydrolase activity may reduce serum cholesterol levels through deconjugation of BAs and increasing their excretion [112,113]. This increases the need for de novo bile synthesis, leading to increased conversion of cholesterol into BAs in the liver, thus reducing serum cholesterol levels. SCFAs have been shown to regulate cholesterol metabolism and reduce hepatic cholesterol synthesis [114]. Other potential mechanisms of action include probiotic-driven cholesterol assimilation and increasing cholesterol binding to cell walls of probiotics in the small intestine, thereby decreasing the amount absorbed by the body [115]. A 2021 study showed that L. plantarum K50 supplementation in HFD-induced obese mice decreased the serum triglyceride level and increased HDL-C levels [116]. Ahn et al. evaluated the triglyceride-lowering effect of supplementation with Lactobacillus curvatus HY7601 and L. plantarum KY1032. The RCT was performed over 12 weeks and involved 92 participants with hypertriglyceridaemia. Probiotic supplementation resulted in a significant 20% reduction in serum triglycerides and a 25% increase in apo A-V. Another RCT studied the effect of a fermented soy product containing Enterococcus faecium CRL 183 and Lactobacillus helveticus 416 and isoflavone on CVD risk markers in moderately hypercholesteraemic men for 42 days [117]. Consumption of the probiotic soy product with isoflavone improved total cholesterol (TC), LDL-C and electronegative LDL concentrations. The levels of high-density lipoprotein-cholesterol (HDL-C) was unaffected and the level of fibrinogen and C-reactive protein (CRP) levels were not improved. The effect of a combination of Lactobacillus acidophilus and Bifidobacterium bifidum or placebo for six weeks on lowering serum cholesterol in 70 hypercholesteraemic patients was evaluated [118]. The probiotic group exhibited decreases in TC, HDL-C and LDL-C levels compared with the control at the end of treatment. We previously demonstrated the potential of exopolysaccharide (EPS)-producing Lactobacillus mucosae DPC 6426 as hypocholesterolaemia therapy in apo E deficient (apoE−/−) mice, fed a high fat/high cholesterol diet [119]. In another study, we fed apoE−/− mice either a high-fat/cholesterol diet alone or in conjunction with plant sterol ester, oat β-glucan (OBG), BSH active Lactobacillus reuteri APC 2587, or the drug atorvastatin [120]. OBG intervention resulted in the most favourable shifts in microbiome composition and functionality, and OBG mice appeared to be protected from the high-fat/cholesterol-induced atherogenesis.
Pregnancy is associated with elevated levels of lipid concentrations. In a RCT conducted by Hoppu et al., 256 pregnant women were randomised into three study groups: dietary counselling with probiotics (L. rhamnosus GG and Bifidobacterium lactis) or dietary counselling with placebo or control in the first trimester of pregnancy [121]. Similar lipid levels were observed in all groups during pregnancy. However, postpartum LDL-C and TC were lower in both the probiotic and placebo dietary counselling groups compared with controls. Another RCT examined the effects of daily supplementation of a probiotic yoghurt containing Streptococcus thermophilus, Lactobacillus bulgaricus, L. acidophilus LA-5, and Bifidobacterium animalis BB12 among 70 pregnant women in the third trimester for 9 weeks [122]. Consumption of the probiotic yoghurt resulted in a significant reduction in TC, LDL-C and HDL-C levels, as well as serum triglyceride concentration. However, no significant differences were found in serum lipid profiles.
Probiotics have also been studied for their putative antihypertensive effects. Probiotics may exert this antihypertensive effect through reducing blood glucose levels, insulin resistance and by improving cholesterol levels and endothelial dysfunction [123,124]. Seppo et al. evaluated the BP-lowering effect of milk fermented by L. helveticus LBK-16H in 39 hypertensive subjects [125]. L. helveticus LBK-16H supplementation resulted in decreases in both SBP and diastolic blood pressure (DBP) compared with control group. Ivey et al. examined the effect of L. acidophilus La5 and B. lactis Bb12 on BP in 156 overweight men and women over 55 years [126]. When compared to control, probiotic yoghurt did not significantly alter BP, heart rate or serum lipid concentrations. A 2015 RCT examined the effects of probiotic soy milk containing L. plantarum A7 on BP in type 2 diabetic (T2D) patients [127]. Probiotic soymilk was reported to significantly decrease SBP and DBP. Aihara et al. demonstrated that daily consumption of fermented milk with L. helveticus CM4 in subjects with high–normal BP or mild hypertension decreased SBP and DBP without any adverse effects [128]. In a 2020 study, administration of L. plantarum WJL during pregnancy and lactation in dams was reported to reduce BP and prevent cardiovascular dysfunction in male offspring [129].
The potential for probiotic bacteria to decrease ROS and oxidative stress has also been examined. Probiotics may exert antioxidant effects through regulation of antioxidant enzymes, scavenging of ROS or chelating of metal ions [42]. LeBlanc et al. demonstrated that mice receiving catalase or superoxide dismutase-producing Lactobacillus casei BL23 had a faster recovery of initial weight loss, increased enzymatic activities in the gut and less colonic inflammation compared to animals that received wild-type strain [130]. Ejtahed et al. assessed the effect of a probiotic yogurt containing L. acidophilus La5 and B. lactis Bb12 on blood glucose and antioxidant status in 64 T2D patients [131]. Subjects receiving probiotic yogurt had significantly increased erythrocyte superoxide dismutase and glutathione peroxidase activities and total antioxidant status compared with the control group. Gómez-Guzmán et al. demonstrated that administration of Lactobacillus fermentum CECT5716, or Lactobacillus coryniformis CECT5711 plus Lactobacillus gasseri CECT5714 improved endothelial dysfunction, vascular inflammation and vascular oxidative stress, in SHR. Xin et al. reported that supplementation of Lactobacillus johnsonii BS15 alleviated HFD-induced oxidative stress in non-alcoholic fatty liver disease (NAFLD) mice, suggesting that this strain can improve host redox state [132]. Friques et al. evaluated the effects of kefir treatment for 60 days on endothelial function and vascular responsiveness in SHR [133]. Kefir was reported to attenuate increased ROS production and decreased nitric oxide NO bioavailability in SHR, thereby improving endothelial function.
Malik et al. examined whether oral L. plantarum 299v supplementation for six weeks in 24 men with stable CAD improves vascular endothelial function and decreases systematic inflammation [128]. Lp299v supplementation resulted in improvements in endothelium-dependent vasodilation and increased NO bioavailability. In addition, a decrease in systemic inflammation with reduced IL-8, IL-12 and leptin levels was observed. It was also found that the SCFA propionate increased with Lp299v supplementation. A 12 week RCT examined the effect of a multispecies probiotic, Ecologic® Barrier, on endothelial dysfunction in obese postmenopausal women [134]. The probiotic preparation contained Bifidobacterium bifidum W23, B. lactis W51, B. lactis W52, L. acidophilus W37, Lactobacillus brevis W63, Lactobacillus casei W56, Lactobacillus salivarius W24, Lactococcus lactis W19, and Lactococcus lactis W58. Eighty-one women participated in the trial and were randomised to receive either a low dose of probiotic, a high dose or placebo daily. The trial revealed a significant reduction in SBP, vascular endothelial growth factor, interleukin-6 (IL-6), tumour necrosis factor alpha (TNF-α), and thrombomodulin following high-dose probiotic supplementation. Decreased SBP and IL-6 were observed with low doses of probiotic supplementation The anti-inflammatory effects of probiotics in relation to CVD were also examined in the 2019 PROSIR study. This study evaluated the effect of L. reuteri V3401 on gut microbiota composition, biomarkers of inflammation, CVD risk and hepatic steatosis in obese adults with MetS [127]. The RCT included 60 participants, aged 18 to 65 years, who were randomised to receive daily dose of L. reuteri V3401 or placebo for 12 weeks. Consumption of L. reuteri V3401 was associated with lower levels of inflammatory biomarkers IL-6 and soluble intercellular adhesion molecule-1 (sVCAM-1).
As previously discussed, TMAO has been reported to be a risk factor in the development of CVD. A 2015 RCT examined L. casei Shirota (LcS) supplementation on TMAO levels in 13 patients with MetS [135]. There were no significant differences observed in TMAO levels between subjects receiving LcS and controls. Tenore et al. tested the effects of lactofermented Annurca apple puree (lfAAP) containing L. rhamnosus LRH11 and L. plantarum SGL07 on plasma lipid profiles and TMAO levels in 90 individuals with CVD risk factors [136]. lfAAP supplementation significantly increased HDL-C levels and lowered TMAO levels at the end of the intervention period. Malik et al. reported that L. plantarum 299v supplementation in 20 men with stable CAD did not significantly change plasma TMAO concentrations [137]. Qiu et al. reported that L. plantarum ZDY04 significantly reduced serum TMAO and cecal TMA levels, as well as significantly inhibited the development of TMAO-induced atherosclerosis in ApoE−/− 1.3% choline-fed mice compared to controls [138]. Boutagy et al. investigated if supplementation of the multistrain probiotic VSL#3 could attenuate the increase in fasting plasma concentrations of TMAO in 19 healthy males following a HFD [139]. The authors reported that no differences in TMAO levels were observed between the VSL#3 arm and the control arm of the study. Matsumoto et al. investigated the effect of B. lactis LKM512 on colonic TMA in healthy volunteers [140]. Faecal TMA concentration and the proportion of TMA-producing bacteria were found to be lower in the probiotic group than in the placebo. Borges et al. evaluated the effects of S. thermophilus (KB19), L. acidophilus (KB27), and Bifidobacterium longum (KB31) supplementation on TMAO plasma levels in chronic kidney disease patients on haemodialysis [141]. Probiotic supplementation did not appear to change plasma TMAO levels. However, a significant increase in betaine plasma levels was observed.
A 2015 meta-analysis included 15 studies totalling 788 adults to examine the effect of probiotics on the reduction in CVD risk factors [142]. The authors reported a statistically significant pooled effect reduction in TC, LDL-C, BMI, waist circumference and inflammatory markers. A significant reduction in LDL-C was reported when L. acidophilus was used in a trial compared to other strains. Subgroup analysis revealed that probiotics were most effective on TC and LDL-C when consumed in the form of fermented milk or yoghurt compared to capsule form, consumption was at least eight weeks, and were multistrain rather than a single strain intervention. Mo et al. assessed the efficacy of probiotics in lowering serum lipid concentrations in a meta-analysis of 19 RCTs including 967 hypercholesterolemia adults [143]. The meta-analysis indicated that the use of probiotics can lower TC and LDL-C compared to controls. These effects were greatest for longer intervention times, certain probiotic strains, and in younger mildly hypercholesterolemia subjects. No significant effects on TG and HDL-C levels were observed. The effects of probiotics on decreasing TC and LDL-C levels were greater for longer intervention times and in younger mildly hypercholesterolemia subjects. With regard to strain specific effects, L. acidophilus, L. plantarum and L. reuteri significantly decreased TC. L. plantarum, L. helveticus and E. faecium significantly reduced LDL-C. No significant beneficial effects were observed for L. fermentum, L. rhamnosus and mixtures of L. acidophilus and Bifidobacterium.
A more recent and larger 2020 systematic review and meta-analysis examined the efficacy of probiotics on lowering CVD risk factors (obesity, high BP, high cholesterol and triglycerides, elevated HbA1c and serum glucose) in patients with CVD or co-morbidities, such as hypertension, obesity, type II diabetes, MetS and hypercholesterolemia [144]. Thirty-four RCTs with 2177 adults who received probiotic intervention (n = 1176) or control (1001) were included in the analysis. An overall statistically significant reduction in SBP and DBP, TC, LDL-C, serum glucose, HbA1c and BMI with probiotic intake was reported. Subgroup analysis revealed that the statistically significant effects of probiotics on the lowering of risk factors might be related to patient morbidity. Diabetic patients displayed the greatest reduction in SBP, DBP, triglycerides, HDL-C and fasting glucose. Hypercholesterolemia patients displayed the greatest reduction in TC and LDL-C, as did obese patients in reducing BMI. Female populations were reported to have more significant reduction in fasting glucose, TC, LDL-C, HbA1c and BMI compared to males. The authors hypothesise that the oestrogen-gut microbiome axis may contribute to this observed reduction in CVD risk factors. Concerning probiotic formulation, the use of probiotics in yoghurt was statistically favoured for a reduction in TC, HbA1c, fasting glucose and LDL-C. Kefir and powder formulations were associated with significant reductions in BMI. Results favoured a longer duration of >1.5 months for reductions in SBP and DBP, while shorter duration favoured significant reductions in TC, LDL-C HbA1c and fasting glucose. Dosage of probiotics was also important as higher daily doses (>1 × 109 CFU) were reported to be more effective in reducing TC, BMI, HbA1c and fasting glucose. Limitations of this meta-analysis include small population sizes and the use of different probiotic strains and doses in the various studies.
5.2. Faecal Microbiota Transplantation
Faecal microbiota transplantation (FMT) is emerging as an innovative therapeutic solution for many microbiota-associated conditions. FMT involves the collection of filtered faecal samples from carefully screened health donors and its placement into the intestinal tract of the recipient with the aim of restoring the normal functions of the gut microbiota. Eisman et al. first described the use of FMT, via faecal enema, as an adjunct in the treatment of pseudomembranous colitis [145]. In particular, the use of FMT in the treatment of recurrent Clostridium difficile infection has proved extremely successful and has been demonstrated to be significantly more effective than using common treatment methods such as vancomycin antibiotic treatment [146,147]. FMT is also emerging as a potential therapeutic intervention for other microbiota-associated disorders including ulcerative colitis, Crohn’s disease, obesity, type 2 diabetes and CVD.
A pioneering study by Bäckhed et al. showed a 60% increase in body fat content and insulin resistance in GF mice inoculated with FMT from conventionally raised mice [47]. Kootte et al. studied the effect of lean donor versus autologous FMT to male recipients with MetS [148]. Lean donor FMT significantly improved insulin sensitivity accompanied by altered gut microbiota composition at six weeks. The FMT-TRIM pilot trial randomised 24 adults with obesity who were at high risk for development of T2D to receive either weekly oral FMT capsules from lean donors or placebo [149]. No significant improvements in insulin sensitivity or metabolic outcomes in the FMT group compared to the placebo group were observed. Limitations of this study were the small sample size, inclusion of subjects with relatively mild insulin resistance, and lack of concurrent dietary intervention. Allegretti et al. examined the effects of FMT with oral capsules from a lean donor in 22 obese subjects [150]. The microbiota composition and bile acid profiles of subjects who received FMT were observed to have shifted towards that of the donor.
Mell et al. used rat models of hypertension and normotension, Dahl salt-sensitive (S) and Dahl salt-resistant (R) rats, to examine whether differences in microbiota contribute to BP using caecal microbiota transplantation [151]. Caecal transplantation did not alter the SBP of the R rats. By contrast, SBP of the S rats given R rat microbiota was significantly elevated during the rest of their life, and they also had a shorter lifespan. Compositional analysis indicated that reduced faecal Veillonellaceae and increased plasma heptanoate and acetate were associated with the increased BP observed in the S rats given R rat caecal content. Furthermore, it has been demonstrated that transplantation of caecal contents from hypertensive obstructive sleep apnoea rats on HFD into recipient rats on normal chow diet lead to higher BP levels [152]. Recipient rats receiving caecal contents from a sham donor on a HFD had no change in BP. The findings suggest that the hypertensive phenotype of obstructive sleep apnoea rats is transferrable via the caecal contents. Li et al. also reported that high blood pressure could be transferred by FMT [90]. FMT from hypertensive human donors to GF mice resulted in elevated BP. Gregory et al. investigated if caecal microbial transplantation can transmit choline diet-induced TMAO production and atherosclerosis susceptibility [153]. An atherosclerosis-prone and high TMAO-producing model, C57BL/6J, and an atherosclerosis-resistant and low TMAO-producing model, NZW/LacJ, were used as donors into antibiotic treated Apoe−/− mouse mice. Enhanced atherosclerosis was observed in mice receiving caecal microbes from atherosclerosis-prone donors compared to those from atherosclerosis-resistant donors. The increase in atherosclerosis was only observed with a high choline diet. Differences in TMA and TMAO levels were initially reported in the post-transplantation period. However, this was not maintained to the end of the study.
In a study carried out by Hu et al., using an experimental autoimmune myocarditis (EAM) mouse model investigated the use of FMT for the treatment of myocarditis was investigated [154]. FMT treatment was shown to restore the Bacteroidetes population and reduce the incidence of myocarditis. Another preclinical study investigated if changes in the gut microbiota induced by reciprocal FMT from normotensive Wistar-Kyoto (WKY) to SHR could alter neuroinflammation and sympathetic activity and thereby lower BP [155]. The animals were divided into four groups: WKY transplanted with WKY microbiota, SHR with SHR, WKY with SHR, and SHR with WKY. Following FMT from WKY to SHR rats, basal SBP and DBP decreased, however, heart rate did not change significantly. An increase in basal SBP and DBP was also observed after FMT from SHR to WKY rats. Correlation analyses showed a negative correlation between the abundances of Blautia and Odoribacter and high SBP. Kim et al. demonstrated that FMT from healthy donor mice who were fed the polyphenol, Resveratrol, to hypertensive mice had beneficial effects in significantly lowering SBP as well as improving glucose homeostasis during diet-induced obesity [156]. Smits et al. investigated the effects of FMT from vegan donors on TMAO production and vascular inflammation in 20 obese male subjects suffering from MetS [157]. Dietary L-carnitine, which is converted by members of the intestinal microbiota to TMAO, is abundant in red meat, while, individuals with a vegan diet have lower amounts of circulating TMAO [19,158]. Vegan-donor FMT resulted in changes in the gut microbiota composition in some but not all recipients. Beneficial changes in TMAO production or parameters relating to vascular inflammation were not observed.
6. Conclusions and Future Directions
Prevention of CVD is dependent on understanding the mechanisms underlying disease development. Increasing evidence points to an association between alterations in the composition of the gut microbiota and microbial metabolites with the development of CVD. Studies analysing microbial communities have identified several taxa associated with CVD development and have emphasised the importance of oral microbiota in CVD risk assessment and therapeutics. More conclusive evidence is still needed to determine whether alterations in gut microbiota composition are a consequence or a causal factor in the pathogenesis of CVD. Rather than focusing on individual taxa, profiling of microbial metabolic potential and functional genetic alterations in CVD through metagenomic and metabolomic approaches will aid our understanding of the influence of the gut microbiota on CVD pathogenesis. The combined use with other “omics” such as transcriptomics and proteomics will assist in the discovery of novel biomarkers and could act as a form of microbial fingerprinting, facilitating more targeted precision therapies. Therapeutic strategies focusing on the inhibition or blocking of microbial metabolic pathways such as the TMAO signalling pathway could have potential for CVD intervention. Probiotic interventions hold promise in CVD risk prevention, though larger multicentre trials, with information on doses, frequencies of administration and specific strains must be performed to generate more convincing data. The use of FMT in the treatment of CVD has shown some promise as a new therapeutic approach. However, much of the research has been performed in preclinical rather than clinical settings. Further limitations associated with the use of FMT include the possibility of the transfer of endotoxin and infectious agents, resulting in new complications. In conclusion, the gut microbiome plays an important role in the pathogenesis of CVD and targeting the gut microbiota with appropriate therapeutic interventions is a promising strategy for better CVD prevention and management in the future.
Author Contributions
Conceptualization, K.M., A.N.O., R.P.R. and C.S.; writing—original draft preparation, K.M. and A.N.O.; writing—review and editing, K.M., A.N.O., N.M.C., R.P.R. and C.S.; supervision, R.P.R. and C.S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are supported in part by Science Foundation Ireland in the form of a centre grant for APC Microbiome Ireland Grant No. SFI/12/RC/2273-P2 and VistaMilk Grant No. SFI16/RC/3835.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stewart, J.; Manmathan, G.; Wilkinson, P. Primary prevention of cardiovascular disease: A review of contemporary guidance and literature. JRSM Cardiovasc. Dis. 2017, 6, 2048004016687211. [Google Scholar] [CrossRef] [Green Version]
- Linton, M.R.F.; Yancey, P.G.; Davies, S.S.; Jerome, W.G.; Linton, E.F.; Song, W.L.; Doran, A.C.; Vickers, K.C. The Role of Lipids and Lipoproteins in Atherosclerosis. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar]
- Tang, W.H.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [Green Version]
- WHO. The Challenge of Cardiovascular Disease—Quick Statistics. Available online: https://www.euro.who.int/en/health-topics/noncommunicable-diseases/cardiovascular-diseases/data-and-statistics (accessed on 11 May 2021).
- Yusuf, S.; Hawken, S.; Ounpuu, S.; Dans, T.; Avezum, A.; Lanas, F.; McQueen, M.; Budaj, A.; Pais, P.; Varigos, J.; et al. Effect of potentially modifiable risk factors associated with myocardial infarction in 52 countries (the INTERHEART study): Case-control study. Lancet 2004, 364, 937–952. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Novakovic, M.; Rout, A.; Kingsley, T.; Kirchoff, R.; Singh, A.; Verma, V.; Kant, R.; Chaudhary, R. Role of gut microbiota in cardiovascular diseases. World J. Cardiol. 2020, 12, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, J.L.; Liebl, A.L. Early colonization of the gut microbiome and its relationship with obesity. Hum. Microbiome J. 2018, 10, 1–5. [Google Scholar] [CrossRef]
- Sirisinha, S. The potential impact of gut microbiota on your health:Current status and future challenges. Asian Pac. J. Allergy Immunol. 2016, 34, 249–264. [Google Scholar] [CrossRef]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut as a Danger Signal for Autoimmune Diseases. Front. Immunol. 2017, 8, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiattarella, G.G.; Sannino, A.; Esposito, G.; Perrino, C. Diagnostics and therapeutic implications of gut microbiota alterations in cardiometabolic diseases. Trends Cardiovasc. Med. 2019, 29, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Tindall, A.M.; Petersen, K.S.; Kris-Etherton, P.M. Dietary Patterns Affect the Gut Microbiome-The Link to Risk of Cardiometabolic Diseases. J. Nutr. 2018, 148, 1402–1407. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.F.; Dwivedi, G.; O’Gara, F.; Caparros-Martin, J.; Ward, N.C. The gut microbiome and cardiovascular disease: Current knowledge and clinical potential. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H923–H938. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- McCarville, J.L.; Chen, G.Y.; Cuevas, V.D.; Troha, K.; Ayres, J.S. Microbiota Metabolites in Health and Disease. Annu. Rev. Immunol. 2020, 38, 147–170. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpela, K. Diet, Microbiota, and Metabolic Health: Trade-Off Between Saccharolytic and Proteolytic Fermentation. Annu. Rev. Food Sci. Technol. 2018, 9, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, K.; Rey, F.E. The emerging role of gut microbial metabolism on cardiovascular disease. Curr. Opin. Microbiol. 2019, 50, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, G.; Dinakaran, V.; Rajendhran, J.; Swaminathan, K. Blood Microbiota and Circulating Microbial Metabolites in Diabetes and Cardiovascular Disease. Trends Endocrinol. Metab. 2020, 31, 835–847. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, M. Trimethylamine N-Oxide Generated by the Gut Microbiota Is Associated with Vascular Inflammation: New Insights into Atherosclerosis. Mediat. Inflamm. 2020, 2020, 4634172. [Google Scholar] [CrossRef] [Green Version]
- Fennema, D.; Phillips, I.R.; Shephard, E.A. Trimethylamine and Trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-Mediated Host-Microbiome Metabolic Axis Implicated in Health and Disease. Drug Metab. Dispos. 2016, 44, 1839–1850. [Google Scholar] [CrossRef] [Green Version]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep. 2015, 10, 326–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthong, V.; Wang, Z.; Li, X.S.; Fan, Y.; Wu, Y.; Tang, W.H.; Hazen, S.L. Intestinal Microbiota-Generated Metabolite Trimethylamine-N-Oxide and 5-Year Mortality Risk in Stable Coronary Artery Disease: The Contributory Role of Intestinal Microbiota in a COURAGE-Like Patient Cohort. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Roncal, C.; Martinez-Aguilar, E.; Orbe, J.; Ravassa, S.; Fernandez-Montero, A.; Saenz-Pipaon, G.; Ugarte, A.; Estella-Hermoso de Mendoza, A.; Rodriguez, J.A.; Fernandez-Alonso, S.; et al. Trimethylamine-N-Oxide (TMAO) Predicts Cardiovascular Mortality in Peripheral Artery Disease. Sci. Rep. 2019, 9, 15580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koay, Y.C.; Chen, Y.C.; Wali, J.A.; Luk, A.W.S.; Li, M.; Doma, H.; Reimark, R.; Zaldivia, M.T.K.; Habtom, H.T.; Franks, A.E.; et al. Plasma levels of trimethylamine-N-oxide can be increased with ‘healthy’ and ‘unhealthy’ diets and do not correlate with the extent of atherosclerosis but with plaque instability. Cardiovasc. Res. 2021, 117, 435–449. [Google Scholar] [CrossRef]
- Sina, C.; Gavrilova, O.; Forster, M.; Till, A.; Derer, S.; Hildebrand, F.; Raabe, B.; Chalaris, A.; Scheller, J.; Rehmann, A.; et al. G protein-coupled receptor 43 is essential for neutrophil recruitment during intestinal inflammation. J. Immunol. 2009, 183, 7514–7522. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluznick, J. A novel SCFA receptor, the microbiota, and blood pressure regulation. Gut Microbes 2014, 5, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, V.; Gueimonde, M.; Pajunen, L.; Nieuwdorp, M.; Laitinen, K. How strong is the evidence that gut microbiota composition can be influenced by lifestyle interventions in a cardio-protective way? Atherosclerosis 2020, 311, 124–142. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Harris, S.C.; Bhowmik, S.; Kang, D.J.; Hylemon, P.B. Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes 2016, 7, 22–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.M.; Hazen, S.L. Microbial modulation of cardiovascular disease. Nat. Rev. Microbiol. 2018, 16, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Muzio, M.; Polntarutti, N.; Bosisio, D.; Prahladan, M.K.; Mantovani, A. Toll like receptor family (TLT) and signalling pathway. Eur. Cytokine Netw. 2000, 11, 489–490. [Google Scholar]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Wiedermann, C.J.; Kiechl, S.; Dunzendorfer, S.; Schratzberger, P.; Egger, G.; Oberhollenzer, F.; Willeit, J. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: Prospective results from the Bruneck Study. J. Am. Coll. Cardiol. 1999, 34, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Gomez-Guzman, M.; Toral, M.; Romero, M.; Jimenez, R.; Galindo, P.; Sanchez, M.; Zarzuelo, M.J.; Olivares, M.; Galvez, J.; Duarte, J. Antihypertensive effects of probiotics Lactobacillus strains in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Deledda, A.; Annunziata, G.; Tenore, G.C.; Palmas, V.; Manzin, A.; Velluzzi, F. Diet-Derived Antioxidants and Their Role in Inflammation, Obesity and Gut Microbiota Modulation. Antioxidants 2021, 10, 708. [Google Scholar] [CrossRef]
- Liu, H.; Zhuang, J.; Tang, P.; Li, J.; Xiong, X.; Deng, H. The Role of the Gut Microbiota in Coronary Heart Disease. Curr. Atheroscler. Rep. 2020, 22, 77. [Google Scholar] [CrossRef]
- Kelly, T.N.; Bazzano, L.A.; Ajami, N.J.; He, H.; Zhao, J.; Petrosino, J.F.; Correa, A.; He, J. Gut Microbiome Associates With Lifetime Cardiovascular Disease Risk Profile Among Bogalusa Heart Study Participants. Circ. Res. 2016, 119, 956–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: A cross-sectional study. Gut Pathog. 2013, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Krajmalnik-Brown, R.; Ilhan, Z.E.; Kang, D.W.; DiBaise, J.K. Effects of gut microbes on nutrient absorption and energy regulation. Nutr. Clin. Pr. 2012, 27, 201–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [Green Version]
- Tims, S.; Derom, C.; Jonkers, D.M.; Vlietinck, R.; Saris, W.H.; Kleerebezem, M.; de Vos, W.M.; Zoetendal, E.G. Microbiota conservation and BMI signatures in adult monozygotic twins. ISME J. 2013, 7, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef]
- Kalliomaki, M.; Collado, M.C.; Salminen, S.; Isolauri, E. Early differences in fecal microbiota composition in children may predict overweight. Am. J. Clin. Nutr. 2008, 87, 534–538. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Davis, S.C.; Yadav, J.S.; Barrow, S.D.; Robertson, B.K. Gut microbiome diversity influenced more by the Westernized dietary regime than the body mass index as assessed using effect size statistic. Microbiologyopen 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef]
- Palleja, A.; Kashani, A.; Allin, K.H.; Nielsen, T.; Zhang, C.; Li, Y.; Brach, T.; Liang, S.; Feng, Q.; Jorgensen, N.B.; et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome Med. 2016, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Schneeberger, M.; Everard, A.; Gomez-Valades, A.G.; Matamoros, S.; Ramirez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seck, E.H.; Senghor, B.; Merhej, V.; Bachar, D.; Cadoret, F.; Robert, C.; Azhar, E.I.; Yasir, M.; Bibi, F.; Jiman-Fatani, A.A.; et al. Salt in stools is associated with obesity, gut halophilic microbiota and Akkermansia muciniphila depletion in humans. Int. J. Obesity 2019, 43, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Medina, D.A.; Pedreros, J.P.; Turiel, D.; Quezada, N.; Pimentel, F.; Escalona, A.; Garrido, D. Distinct patterns in the gut microbiota after surgical or medical therapy in obese patients. PeerJ 2017, 5, e3443. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zhang, Y.; Wang, X.; Yang, R.; Zhu, X.; Zhang, Y.; Chen, C.; Yuan, H.; Yang, Z.; Sun, L. Gut bacteria Akkermansia is associated with reduced risk of obesity: Evidence from the American Gut Project. Nutr. Metab. 2020, 17, 90. [Google Scholar] [CrossRef] [PubMed]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20-76 years of age. NPJ Biofilms Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, G.; Wu, F.; Yang, L.; Kelly, D.; Hu, J.; Li, H.; Jasmine, F.; Kibriya, M.G.; Parvez, F.; Shaheen, I.; et al. The association between gut microbiome and anthropometric measurements in Bangladesh. Gut Microbes 2020, 11, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Companys, J.; Gosalbes, M.J.; Pla-Paga, L.; Calderon-Perez, L.; Llaurado, E.; Pedret, A.; Valls, R.M.; Jimenez-Hernandez, N.; Sandoval-Ramirez, B.A.; Del Bas, J.M.; et al. Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Guryn, K.; Hubert, N.; Frazier, K.; Urlass, S.; Musch, M.W.; Ojeda, P.; Pierre, J.F.; Miyoshi, J.; Sontag, T.J.; Cham, C.M.; et al. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe 2018, 23, 458–469. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergstrom, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Backhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Rebolledo, C.; Cuevas, A.; Zambrano, T.; Acuna, J.J.; Jorquera, M.A.; Saavedra, K.; Martinez, C.; Lanas, F.; Seron, P.; Salazar, L.A.; et al. Bacterial Community Profile of the Gut Microbiota Differs between Hypercholesterolemic Subjects and Controls. Biomed. Res. Int. 2017, 2017, 8127814. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K.; et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Jiang, F.; Cheng, R.; Luo, Y.; Wang, J.; Luo, Z.; Li, M.; Shen, X.; He, F. A high-fat diet and high-fat and high-cholesterol diet may affect glucose and lipid metabolism differentially through gut microbiota in mice. Exp. Anim. 2021, 70, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef]
- Emoto, T.; Yamashita, T.; Sasaki, N.; Hirota, Y.; Hayashi, T.; So, A.; Kasahara, K.; Yodoi, K.; Matsumoto, T.; Mizoguchi, T.; et al. Analysis of Gut Microbiota in Coronary Artery Disease Patients: A Possible Link between Gut Microbiota and Coronary Artery Disease. J. Atheroscler. Thromb. 2016, 23, 908–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.; Drautz-Moses, D.I.; Alhede, M.; Maw, M.T.; Liu, Y.; Purbojati, R.W.; Yap, Z.H.; Kushwaha, K.K.; Gheorghe, A.G.; Bjarnsholt, T.; et al. In silico analyses of metagenomes from human atherosclerotic plaque samples. Microbiome 2015, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.H.; Fak, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Backhed, F.; Nielsen, J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Gao, R.; Zhang, Y.; Pan, D.; Zhu, Y.; Zhang, X.; Yang, R.; Jiang, R.; Xu, Y.; Qin, H. Dysbiosis signatures of gut microbiota in coronary artery disease. Physiol. Genom. 2018, 50, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Bale, B.F.; Doneen, A.L.; Vigerust, D.J. High-risk periodontal pathogens contribute to the pathogenesis of atherosclerosis. Postgrad. Med. J. 2017, 93, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Spor, A.; Felin, J.; Fak, F.; Stombaugh, J.; Tremaroli, V.; Behre, C.J.; Knight, R.; Fagerberg, B.; Ley, R.E.; et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4592–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyvarinen, K.; Mantyla, P.; Buhlin, K.; Paju, S.; Nieminen, M.S.; Sinisalo, J.; Pussinen, P.J. A common periodontal pathogen has an adverse association with both acute and stable coronary artery disease. Atherosclerosis 2012, 223, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Moghadamrad, S.; McCoy, K.D.; Geuking, M.B.; Sagesser, H.; Kirundi, J.; Macpherson, A.J.; De Gottardi, A. Attenuated portal hypertension in germ-free mice: Function of bacterial flora on the development of mesenteric lymphatic and blood vessels. Hepatology 2015, 61, 1685–1695. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Callejo, M.; Mondejar-Parreno, G.; Barreira, B.; Izquierdo-Garcia, J.L.; Morales-Cano, D.; Esquivel-Ruiz, S.; Moreno, L.; Cogolludo, A.; Duarte, J.; Perez-Vizcaino, F. Pulmonary Arterial Hypertension Affects the Rat Gut Microbiome. Sci. Rep. 2018, 8, 9681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmu, J.; Salosensaari, A.; Havulinna, A.S.; Cheng, S.; Inouye, M.; Jain, M.; Salido, R.A.; Sanders, K.; Brennan, C.; Humphrey, G.C.; et al. Association Between the Gut Microbiota and Blood Pressure in a Population Cohort of 6953 Individuals. J. Am. Heart Assoc. 2020, 9, e016641. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Wen, Z.; Meng, K.; Yang, P. Metabolomic signatures for liver tissue and cecum contents in high-fat diet-induced obese mice based on UHPLC-Q-TOF/MS. Nutr. Metab. 2021, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Li, B.; Zheng, N.; Wu, G.; Ma, J.; Tao, X.; Chen, L.; Zhong, J.; Sheng, L.; Li, H. Integrated Metagenomic and Metabolomic Analyses of the Effect of Astragalus Polysaccharides on Alleviating High-Fat Diet-Induced Metabolic Disorders. Front. Pharm. 2020, 11, 833. [Google Scholar] [CrossRef]
- Medina, D.A.; Li, T.; Thomson, P.; Artacho, A.; Perez-Brocal, V.; Moya, A. Cross-Regional View of Functional and Taxonomic Microbiota Composition in Obesity and Post-obesity Treatment Shows Country Specific Microbial Contribution. Front. Microbiol. 2019, 10, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.Q.; Mills, R.H.; Peters, N.V.; Holder, M.K.; de Vries, G.J.; Knight, R.; Chassaing, B.; Gonzalez, D.J.; Gewirtz, A.T. Associations of the Fecal Microbial Proteome Composition and Proneness to Diet-induced Obesity. Mol. Cell Proteom. 2019, 18, 1864–1879. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Liu, Z.; Zhong, S.; Li, R.; Xia, H.; Jie, Z.; Wen, B.; Chen, X.; Yan, W.; Fan, Y.; et al. Integrated metabolomics and metagenomics analysis of plasma and urine identified microbial metabolites associated with coronary heart disease. Sci. Rep. 2016, 6, 22525. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, J.; Guo, J.; Geng, B.; Ji, W.; Zhao, Q.; Li, J.; Liu, X.; Liu, J.; Guo, Z.; et al. Gut-dependent microbial translocation induces inflammation and cardiovascular events after ST-elevation myocardial infarction. Microbiome 2018, 6, 66. [Google Scholar] [CrossRef]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Dong, Y.; Zuo, K.; Han, C.; Jiao, J.; Yang, X.; Li, J. Characteristics and variation of fecal bacterial communities and functions in isolated systolic and diastolic hypertensive patients. BMC Microbiol. 2021, 21, 128. [Google Scholar] [CrossRef]
- Stevens, B.R.; Pepine, C.J.; Richards, E.M.; Kim, S.; Raizada, M.K. Depressive hypertension: A proposed human endotype of brain/gut microbiome dysbiosis. Am. Heart J. 2021, 239, 27–37. [Google Scholar] [CrossRef]
- Hayashi, T.; Yamashita, T.; Watanabe, H.; Kami, K.; Yoshida, N.; Tabata, T.; Emoto, T.; Sasaki, N.; Mizoguchi, T.; Irino, Y.; et al. Gut Microbiome and Plasma Microbiome-Related Metabolites in Patients with Decompensated and Compensated Heart Failure. Circ. J. 2018, 83, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Emoto, T.; Hayashi, T.; Tabata, T.; Yamashita, T.; Watanabe, H.; Takahashi, T.; Gotoh, Y.; Kami, K.; Yoshida, N.; Saito, Y.; et al. Metagenomic analysis of gut microbiota reveals its role in trimethylamine metabolism in heart failure. Int. J. Cardiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Ye, L.; Li, J.; Jin, L.; Wang, W.; Li, S.; Bao, M.; Wu, S.; Li, L.; Geng, B.; et al. Metagenomic and metabolomic analyses unveil dysbiosis of gut microbiota in chronic heart failure patients. Sci. Rep. 2018, 8, 635. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Sharafedtinov, K.K.; Plotnikova, O.A.; Alexeeva, R.I.; Sentsova, T.B.; Songisepp, E.; Stsepetova, J.; Smidt, I.; Mikelsaar, M. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients--a randomized double-blind placebo-controlled pilot study. Nutr. J. 2013, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadooka, Y.; Sato, M.; Ogawa, A.; Miyoshi, M.; Uenishi, H.; Ogawa, H.; Ikuyama, K.; Kagoshima, M.; Tsuchida, T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. Br. J. Nutr. 2013, 110, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.; Darimont, C.; Drapeau, V.; Emady-Azar, S.; Lepage, M.; Rezzonico, E.; Ngom-Bru, C.; Berger, B.; Philippe, L.; Ammon-Zuffrey, C.; et al. Effect of Lactobacillus rhamnosus CGMCC1.3724 supplementation on weight loss and maintenance in obese men and women. Br. J. Nutr. 2014, 111, 1507–1519. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Wen, Z.; Li, X.; Meng, K.; Yang, P. Lactobacillus plantarum FRT10 alleviated high-fat diet-induced obesity in mice through regulating the PPARalpha signal pathway and gut microbiota. Appl. Microbiol. Biotechnol. 2020, 104, 5959–5972. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Lin, P.P.; Hsieh, Y.M.; Zhang, Z.Y.; Wu, H.C.; Huang, C.C. Cholesterol-lowering potentials of lactic acid bacteria based on bile-salt hydrolase activity and effect of potent strains on cholesterol metabolism in vitro and in vivo. Sci. World J. 2014, 2014, 690752. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.I.; Gibson, G.R. Cholesterol assimilation by lactic acid bacteria and bifidobacteria isolated from the human gut. Appl. Environ. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef] [Green Version]
- Joung, H.; Chu, J.; Kim, B.K.; Choi, I.S.; Kim, W.; Park, T.S. Probiotics ameliorate chronic low-grade inflammation and fat accumulation with gut microbiota composition change in diet-induced obese mice models. Appl. Microbiol. Biotechnol. 2021, 105, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, D.C.; Manzoni, M.S.; Bedani, R.; Roselino, M.N.; Celiberto, L.S.; Vendramini, R.C.; de Valdez, G.; Abdalla, D.S.; Pinto, R.A.; Rosetto, D.; et al. Probiotic Soy Product Supplemented with Isoflavones Improves the Lipid Profile of Moderately Hypercholesterolemic Men: A Randomized Controlled Trial. Nutrients 2016, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Rerksuppaphol, S.; Rerksuppaphol, L. A Randomized Double-blind Controlled Trial of Lactobacillus acidophilus Plus Bifidobacterium bifidum versus Placebo in Patients with Hypercholesterolemia. J. Clin. Diagn. Res. 2015, 9, KC01-04. [Google Scholar] [CrossRef] [PubMed]
- London, L.E.; Kumar, A.H.; Wall, R.; Casey, P.G.; O’Sullivan, O.; Shanahan, F.; Hill, C.; Cotter, P.D.; Fitzgerald, G.F.; Ross, R.P.; et al. Exopolysaccharide-producing probiotic Lactobacilli reduce serum cholesterol and modify enteric microbiota in ApoE-deficient mice. J. Nutr. 2014, 144, 1956–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, P.M.; London, L.E.; Bjorndahl, T.C.; Mandal, R.; Murphy, K.; Fitzgerald, G.F.; Shanahan, F.; Ross, R.P.; Wishart, D.S.; Caplice, N.M.; et al. Microbiome and metabolome modifying effects of several cardiovascular disease interventions in apo-E(-/-) mice. Microbiome 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Hoppu, U.; Isolauri, E.; Koskinen, P.; Laitinen, K. Maternal dietary counseling reduces total and LDL cholesterol postpartum. Nutrition 2014, 30, 159–164. [Google Scholar] [CrossRef]
- Asemi, Z.; Samimi, M.; Tabasi, Z.; Talebian, P.; Azarbad, Z.; Hydarzadeh, Z.; Esmaillzadeh, A. Effect of daily consumption of probiotic yoghurt on lipid profiles in pregnant women: A randomized controlled clinical trial. J. Matern. Fetal. Neonatal. Med. 2012, 25, 1552–1556. [Google Scholar] [CrossRef]
- Qi, D.; Nie, X.L.; Zhang, J.J. The effect of probiotics supplementation on blood pressure: A systemic review and meta-analysis. Lipids Health Dis. 2020, 19, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, P.I.; Kivimaki, A.S.; Turpeinen, A.M.; Korpela, R.; Vapaatalo, H. High blood pressure-lowering and vasoprotective effects of milk products in experimental hypertension. Br. J. Nutr. 2011, 106, 1353–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seppo, L.; Jauhiainen, T.; Poussa, T.; Korpela, R. A fermented milk high in bioactive peptides has a blood pressure-lowering effect in hypertensive subjects. Am. J. Clin. Nutr. 2003, 77, 326–330. [Google Scholar] [CrossRef]
- Ivey, K.L.; Hodgson, J.M.; Kerr, D.A.; Thompson, P.L.; Stojceski, B.; Prince, R.L. The effect of yoghurt and its probiotics on blood pressure and serum lipid profile; a randomised controlled trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Hariri, M.; Salehi, R.; Feizi, A.; Mirlohi, M.; Kamali, S.; Ghiasvand, R. The effect of probiotic soy milk and soy milk on anthropometric measures and blood pressure in patients with type II diabetes mellitus: A randomized double-blind clinical trial. ARYA Atheroscler. 2015, 11, 74–80. [Google Scholar]
- Aihara, K.; Kajimoto, O.; Hirata, H.; Takahashi, R.; Nakamura, Y. Effect of powdered fermented milk with Lactobacillus helveticus on subjects with high-normal blood pressure or mild hypertension. J. Am. Coll. Nutr. 2005, 24, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, K.S.L.; Braga, V.A.; Noronha, S.; Costa, W.; Makki, K.; Cruz, J.C.; Brandao, L.R.; Chianca, D.A., Jr.; Meugnier, E.; Leulier, F.; et al. Lactiplantibacillus plantarum WJL administration during pregnancy and lactation improves lipid profile, insulin sensitivity and gut microbiota diversity in dyslipidemic dams and protects male offspring against cardiovascular dysfunction in later life. Food Funct. 2020, 11, 8939–8950. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; del Carmen, S.; Miyoshi, A.; Azevedo, V.; Sesma, F.; Langella, P.; Bermudez-Humaran, L.G.; Watterlot, L.; Perdigon, G.; de Moreno de LeBlanc, A. Use of superoxide dismutase and catalase producing lactic acid bacteria in TNBS induced Crohn’s disease in mice. J. Biotechnol. 2011, 151, 287–293. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Zeng, D.; Wang, H.; Ni, X.; Yi, D.; Pan, K.; Jing, B. Preventing non-alcoholic fatty liver disease through Lactobacillus johnsonii BS15 by attenuating inflammation and mitochondrial injury and improving gut environment in obese mice. Appl. Microbiol. Biotechnol. 2014, 98, 6817–6829. [Google Scholar] [CrossRef]
- Friques, A.G.; Arpini, C.M.; Kalil, I.C.; Gava, A.L.; Leal, M.A.; Porto, M.L.; Nogueira, B.V.; Dias, A.T.; Andrade, T.U.; Pereira, T.M.; et al. Chronic administration of the probiotic kefir improves the endothelial function in spontaneously hypertensive rats. J. Transl. Med. 2015, 13, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakerian, M.; Razavi, S.H.; Ziai, S.A.; Khodaiyan, F.; Yarmand, M.S.; Moayedi, A. Proteolytic and ACE-inhibitory activities of probiotic yogurt containing non-viable bacteria as affected by different levels of fat, inulin and starter culture. J. Food Sci. Technol. 2015, 52, 2428–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripolt, N.J.; Leber, B.; Triebl, A.; Kofeler, H.; Stadlbauer, V.; Sourij, H. Effect of Lactobacillus casei Shirota supplementation on trimethylamine-N-oxide levels in patients with metabolic syndrome: An open-label, randomized study. Atherosclerosis 2015, 242, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C.; Caruso, D.; Buonomo, G.; D’Avino, M.; Ciampaglia, R.; Maisto, M.; Schisano, C.; Bocchino, B.; Novellino, E. Lactofermented Annurca Apple Puree as a Functional Food Indicated for the Control of Plasma Lipid and Oxidative Amine Levels: Results from a Randomised Clinical Trial. Nutrients 2019, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Suboc, T.M.; Tyagi, S.; Salzman, N.; Wang, J.; Ying, R.; Tanner, M.J.; Kakarla, M.; Baker, J.E.; Widlansky, M.E. Lactobacillus plantarum 299v Supplementation Improves Vascular Endothelial Function and Reduces Inflammatory Biomarkers in Men with Stable Coronary Artery Disease. Circ. Res. 2018, 123, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Tao, X.; Xiong, H.; Yu, J.; Wei, H. Lactobacillus plantarum ZDY04 exhibits a strain-specific property of lowering TMAO via the modulation of gut microbiota in mice. Food Funct. 2018, 9, 4299–4309. [Google Scholar] [CrossRef]
- Boutagy, N.E.; Neilson, A.P.; Osterberg, K.L.; Smithson, A.T.; Englund, T.R.; Davy, B.M.; Hulver, M.W.; Davy, K.P. Probiotic supplementation and trimethylamine-N-oxide production following a high-fat diet. Obesity 2015, 23, 2357–2363. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kitada, Y.; Shimomura, Y.; Naito, Y. Bifidobacterium animalis subsp. lactis LKM512 reduces levels of intestinal trimethylamine produced by intestinal microbiota in healthy volunteers: A double-blind, placebo-controlled study. J. Funct. Foods 2017, 36, 94–101. [Google Scholar] [CrossRef]
- Borges, N.A.; Stenvinkel, P.; Bergman, P.; Qureshi, A.R.; Lindholm, B.; Moraes, C.; Stockler-Pinto, M.B.; Mafra, D. Effects of Probiotic Supplementation on Trimethylamine-N-Oxide Plasma Levels in Hemodialysis Patients: A Pilot Study. Probiotics Antimicrob. Proteins 2019, 11, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Buys, N. Effects of probiotics consumption on lowering lipids and CVD risk factors: A systematic review and meta-analysis of randomized controlled trials. Ann. Med. 2015, 47, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Zhang, X.; Yang, Y. Effect of probiotics on lipid profiles in hypercholesterolaemic adults: A meta-analysis of randomized controlled trials. Med. Clin. 2019, 152, 473–481. [Google Scholar] [CrossRef]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Cox, T.; Robertson, D.; Chi, C.; Sun, J. Efficacy of Probiotics in Patients of Cardiovascular Disease Risk: A Systematic Review and Meta-analysis. Curr. Hypertens. Rep. 2020, 22, 74. [Google Scholar] [CrossRef]
- Eiseman, B.; Silen, W.; Bascom, G.S.; Kauvar, A.J. Fecal enema as an adjunct in the treatment of pseudomembranous enterocolitis. Surgery 1958, 44, 854–859. [Google Scholar] [PubMed]
- van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; de Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.; Tijssen, J.G.; et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.Y.; Dirweesh, A.; Khurshid, T.; Siddiqui, W.J. Comparing fecal microbiota transplantation to standard-of-care treatment for recurrent Clostridium difficile infection: A systematic review and meta-analysis. Eur. J. Gastroenterol. Hepatol. 2018, 30, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Kootte, R.S.; Levin, E.; Salojarvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, E.W.; Gao, L.; Stastka, P.; Cheney, M.C.; Mahabamunuge, J.; Torres Soto, M.; Ford, C.B.; Bryant, J.A.; Henn, M.R.; Hohmann, E.L. Fecal microbiota transplantation for the improvement of metabolism in obesity: The FMT-TRIM double-blind placebo-controlled pilot trial. PLoS Med. 2020, 17, e1003051. [Google Scholar] [CrossRef] [PubMed]
- Allegretti, J.R.; Kassam, Z.; Mullish, B.H.; Chiang, A.; Carrellas, M.; Hurtado, J.; Marchesi, J.R.; McDonald, J.A.K.; Pechlivanis, A.; Barker, G.F.; et al. Effects of Fecal Microbiota Transplantation With Oral Capsules in Obese Patients. Clin. Gastroenterol. Hepatol. 2020, 18, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Mell, B.; Jala, V.R.; Mathew, A.V.; Byun, J.; Waghulde, H.; Zhang, Y.; Haribabu, B.; Vijay-Kumar, M.; Pennathur, S.; Joe, B. Evidence for a link between gut microbiota and hypertension in the Dahl rat. Physiol. Genom. 2015, 47, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durgan, D.J.; Ganesh, B.P.; Cope, J.L.; Ajami, N.J.; Phillips, S.C.; Petrosino, J.F.; Hollister, E.B.; Bryan, R.M., Jr. Role of the Gut Microbiome in Obstructive Sleep Apnea-Induced Hypertension. Hypertension 2016, 67, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Gregory, J.C.; Buffa, J.A.; Org, E.; Wang, Z.; Levison, B.S.; Zhu, W.; Wagner, M.A.; Bennett, B.J.; Li, L.; DiDonato, J.A.; et al. Transmission of atherosclerosis susceptibility with gut microbial transplantation. J. Biol. Chem. 2015, 290, 5647–5660. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.F.; Zhang, W.Y.; Wen, Q.; Chen, W.J.; Wang, Z.M.; Chen, J.; Zhu, F.; Liu, K.; Cheng, L.X.; Yang, J.; et al. Fecal microbiota transplantation alleviates myocardial damage in myocarditis by restoring the microbiota composition. Pharm. Res. 2019, 139, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Toral, M.; Robles-Vera, I.; de la Visitacion, N.; Romero, M.; Yang, T.; Sanchez, M.; Gomez-Guzman, M.; Jimenez, R.; Raizada, M.K.; Duarte, J. Critical Role of the Interaction Gut Microbiota—Sympathetic Nervous System in the Regulation of Blood Pressure. Front. Physiol. 2019, 10, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, L.P.; Kootte, R.S.; Levin, E.; Prodan, A.; Fuentes, S.; Zoetendal, E.G.; Wang, Z.; Levison, B.S.; Cleophas, M.C.P.; Kemper, E.M.; et al. Effect of Vegan Fecal Microbiota Transplantation on Carnitine- and Choline-Derived Trimethylamine-N-Oxide Production and Vascular Inflammation in Patients With Metabolic Syndrome. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef]
- Koeth, R.A.; Lam-Galvez, B.R.; Kirsop, J.; Wang, Z.; Levison, B.S.; Gu, X.; Copeland, M.F.; Bartlett, D.; Cody, D.B.; Dai, H.J.; et al. l-Carnitine in omnivorous diets induces an atherogenic gut microbial pathway in humans. J. Clin. Investig. 2019, 129, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.T.; Parajuli, N.; Sung, M.M.; Bairwa, S.C.; Levasseur, J.; Soltys, C.M.; Wishart, D.S.; Madsen, K.; Schertzer, J.D.; Dyck, J.R.B. Fecal transplant from resveratrol-fed donors improves glycaemia and cardiovascular features of the metabolic syndrome in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E511–E519. [Google Scholar] [CrossRef] [PubMed]

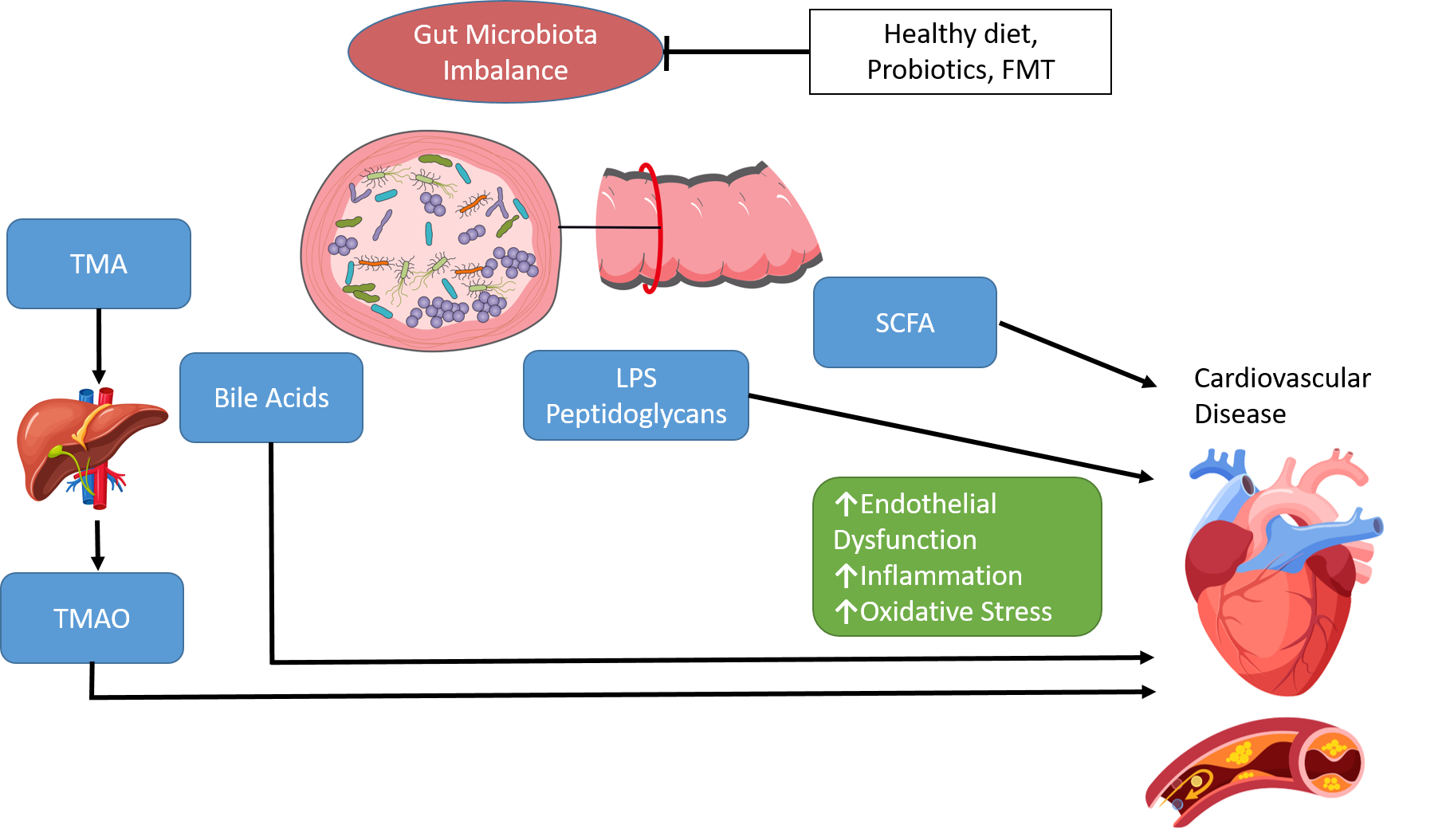{kind=link}
Table 1.
Comparison of published studies of gut microbiome in CVD.
| Study | Year | Cohort | Main Findings | PMID |
|---|---|---|---|---|
| Kelly et al. | 2016 | Bogalusa Heart Study subjects | Prevotella, Tyzzerella ↑ in subjects with high CVD risk. Alpha diversity inversely associated with CVD risk. | 27507222 |
| Bäckhed et al. | 2004 | Germ-free (GF) C57BL/6 mice | FMT from conventionalized mice ↑ body fat and insulin resistance. | 15505215 |
| Bäckhed et al. | 2007 | GF C57BL/6J mice | GF mice protected against diet-induced obesity through AMPK and Fiaf. | 17210919 |
| Ley at al. | 2006 | 12 obese subjects | Obesity ↓ Bacteroidetes ↑ Firmicutes. | 17183309 |
| Ley at al. | 2005 | ob/ob mice, lean ob/+ mice | Obesity ↓ Bacteroidetes ↑ Firmicutes. | 16033867 |
| Turnbaugh et al. | 2009 | Obese and lean twin pairs | Core gut microbiome at level of metabolic functions. | 19043404 |
| Bervoets et al. | 2013 | 26 obese and 27 lean children | Obesity ↑ Firmicutes/Bacteroidetes ratio. | 23631345 |
| Schwiertz et al. | 2010 | 30 lean, 35 overweight and 33 obese subjects. | ↑ Bacteroides from lean to obese. | 19498350 |
| Jumpertz et al. | 2011 | 12 lean and 9 obese subjects | ↑ Firmicutes ↓ Bacteroidetes in lean subjects with an increased energy harvest. | 21543530 |
| Zhang et al. | 2009 | 3 normal weight, 3 morbidly obese, and 3 post-gastric-bypass subjects | Firmicutes dominant in normal-weight and obese subjects. ↓ Firmicutes ↑ Gammaproteobacteria in post-gastric-bypass. | 19164560 |
| Duncan et al. | 2008 | Obese and non-obese subjects | No evidence that Firmicutes/Bacteroidetes have a function in obesity. | 18779823 |
| Tims et al. | 2013 | 40 monozygotic twin pairs | Inverse correlation between Clostridium cluster IV diversity and BMI. Eubacterium ventriosum and Roseburia intestinalis positively correlated to BMI differences. Oscillospira guillermondii negatively correlated to BMI differences. | 23190729 |
| Collado et al. | 2008 | 18 overweight and 36 normal-weight pregnant women | Overweight ↑Bacteroides and Staphylococcus. | 18842773 |
| Kalliomäki et al. | 2008 | 25 overweight and obese children, 24 normal-weight | ↑ Bifidobacteria during infancy in normal-weight. ↑ Staphylococcus aureus during infancy in overweight. | 18326589 |
| Cotillard et al. | 2013 | 38 obese and 11 overweight subjects | Dietary intervention improves low gene richness | 23985875 |
| Davis et al. | 2017 | 81 random Alabama residents | Westernized diet type had a greater impact upon gut microbiota diversity than ↑ BMI | 28677210 |
| Stanislawski et al. | 2019 |
| Gut microbiota phenotypes of obesity may differ with race/ethnicity | 31285833 |
| Palleja et al. | 2016 | 13 morbidly obese patients who underwent Roux-en-Y gastric bypass (RYGB) | ↑ microbial diversity and altered composition post RYGB | 27306058 |
| Schneeberger et al. | 2015 | C57BL/6J mice | Rapid decline of Akkermansia muciniphila during HFD and aging | 26563823 |
| Seck et al. | 2018 | 1326 subjects with variable geo-graphical origin, diet, age, and gender | High fecal salinity linked to ↓diversity and ↓Akkermansia muciniphila and Bifidobacterium | 30206336 |
| Medina et al. | 2017 | 9 obese subjects following medical dietary treatment, 5 following RYGB, 5 following sleeve gastrectomy | ↑Proteobacteria in surgical patients | 28649469 |
| Zhou et al. | 2020 | 10,534 subjects from American Gut Project | Akkermansia is associated with ↓risk of obesity | 33110437 |
| Ozato et al. | 2019 | 1001 Japanese subjects | Blautia associated with visceral fat accumulation | 31602309 |
| Osborne et al. | 2020 | 248 subjects from the Health Effects of Arsenic Longitudinal Study | Correlation between Oscillospira and leanness | 31138061 |
| Companys et al. | 2021 | 96 overweight/obese subjects and 32 lean subjects | Dorea formicigenerans, Dorea longicatena and Collinsella aerofaciens obesity biomarkers. Lachnospiraceae associated with lipid CVD risk factors. | 34199239 |
| Martinez-Guryn et al. | 2018 | C57Bl/6 mice | HFD ↑ Clostridiaceae, ↓Bifidobacteriaceae and Bacteroidaceae GF mice ↓ plasma triglyceride and LDL-C | 29649441 |
| Karlsson et al. | 2013 | 53 women who had Type 2 Diabetes, 49 with impaired glucose tolerance, 43 with normal glucose tolerance | Positive correlation between HDL-C and Clostridium, negative correlation between serum triglycerides and Clostridum | 23719380 |
| Rebolledo et al. | 2017 | 30 hypercholesterolemia subjects, 27 normocholesterolemic controls | ↓ microbial richness and diversity in hypercholesterolemic subjects. | 28698878 |
| Fu et al. | 2015 | 893 subjects from the Life-Lines-DEEP cohort | 34 taxa associated with BMI and blood lipids | 26358192 |
| Liang et al. | 2021 | C57BL/6 mice; normal chow, high-fat diet (HFD), or high-fat and high-cholesterol diet (HFHCD) | ↑ Oscillospira, Odoribacter, Bacteroides, and Prevotella, ↓ Ruminococcus and Akkermansia in HFD compared with HFHCD | 32999215 |
| Zhang et al. | 2021 |
| HFHCD associated with ↑ Mucispirillum, Desulfovibrio, Anaerotruncus and Desulfovibrionaceae; ↓ Bifidobacterium and Bacteroides | 32694178 |
| Emoto et al. | 2016 | 39 coronary artery disease (CAD) patients, 30 controls with coronary risk factors and 50 healthy volunteers | CAD associated with ↑ Lactobacillales, ↓ Bacteroidetes | 26947598 |
| Mira et al. | 2015 | 22 symptomatic and asymptomatic atherosclerotic plaques | Asymptomatic plaques ↑ Porphyromonadaceae, Bacteroidaceae, Micrococcaceae, and Streptococcaceae | 26334731 |
| Karlsson et al. | 2012 | 12 patients with symptomatic atherosclerotic plaques | ↑ Collinsella in patients with symptomatic atherosclerosis | 23212374 |
| Zhu et al. | 2018 | 70 patients with CAD and 98 healthy controls | CAD associated with ↑ Escherichia-Shigella, Enterococcus ↓ Faecalibacterium, Subdoligranulum, Roseburia, Eubacterium rectale | 30192713 |
| Jie et al. | 2017 | 218 individuals with atherosclerotic CVD and 187 healthy controls. | Atherosclerotic CVD associated with ↑ Enterobacteriaceae, Streptococcus | 29018189 |
| Koren et al. | 2011 | 15 patients with atherosclerosis and healthy controls. | Chryseomonas in all atherosclerotic plaque samples, and Veillonella and Streptococcus in the majority. | 20937873 |
| Hyvärinen et al. | 2012 | 179 patients with stable CAD, 166 with acute coronary syndrome, and 119 healthy controls. | ↑ salivary levels of A. actinomycetemcomitans associated with ↑ risk for CAD. | 22704805 |
| Moghadamrad et al. | 2015 | GF male C57BL/6 mice, altered Schaedler flora (ASF) mice, and specific pathogen-free (SPF) mice | GF mice had significantly lower BP than conventional mice | 25643846 |
| Yang et al. | 2015 |
| SHR ↓ microbial richness, diversity, and evenness, ↑ Firmicutes/Bacteroidetes ratio | 25870193 |
| Marques et al. | 2017 | C57Bl/6 mice; control diet, high fiber or acetate | Fiber and acetate ↓ Firmicutes/Bacteroidetes ratio, ↑ Bacteroides acidifaciens | 27927713 |
| Callejo et al. | 2018 | Pulmonary arterial hypertension (PAH)-exposed wistar rats | PAH rats 3X ↑ Firmicutes/Bacteroidetes ratio. | 29946072 |
| Li et al. | 2017 | 41 healthy controls, 56 pre-hypertension, 99 primary hypertension | Pre-hypertensive and hypertensive associated with ↓ microbial richness and diversity, ↑ Prevotella and Klebsiella | 28143587 |
| Palmu et al. | 2020 | 6953 Finns aged 25 to 74 years | α and β diversities strongly related to BP indexes Positive, associations between BP indexes and 45 microbial genera, majority Firmicutes. Negative associations between 19 Lactobacillus species and BP indexes | 32691653 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murphy, K.; O’Donovan, A.N.; Caplice, N.M.; Ross, R.P.; Stanton, C. Exploring the Gut Microbiota and Cardiovascular Disease. Metabolites 2021, 11, 493. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080493
AMA Style
Murphy K, O’Donovan AN, Caplice NM, Ross RP, Stanton C. Exploring the Gut Microbiota and Cardiovascular Disease. Metabolites. 2021; 11(8):493. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080493
Chicago/Turabian StyleMurphy, Kiera, Aoife N. O’Donovan, Noel M. Caplice, R. Paul Ross, and Catherine Stanton. 2021. "Exploring the Gut Microbiota and Cardiovascular Disease" Metabolites 11, no. 8: 493. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.