
Respiratory Colonization and Short-Term Temporal Changes in the Urinary Metabolome of Children


Abstract
:1. Introduction
2. Results
2.1. Study Population Characteristics
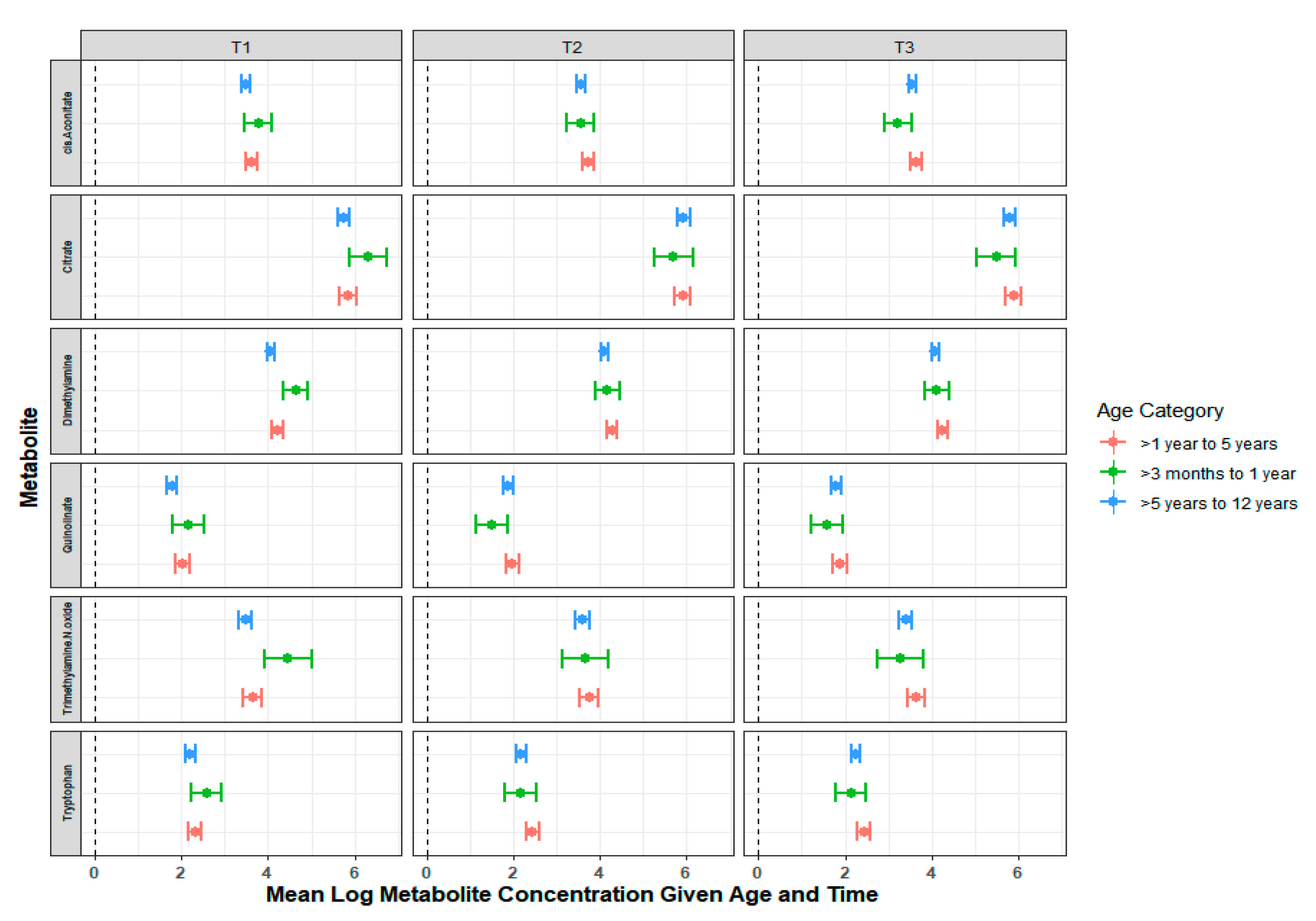
2.2. Temporal Variation in the Urinary Metabolome
2.3. Influence of Bacterial Colonization and Viral Carriage on the Urinary Metabolome
3. Discussion
4. Materials and Methods
4.1. Study Design and Study Subjects
4.2. Study Definitions
4.3. Sample Collection
4.4. Urine Metabolite NMR Measurments
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slupsky, C.M.; Rankin, K.N.; Wagner, J.; Fu, H.; Chang, D.; Weljie, A.M.; Saude, E.J.; Lix, B.; Adamko, D.J.; Shah, S.; et al. Investigations of the effects of gender, diurnal variation, and age in human urinary metabolomic profiles. Anal. Chem. 2007, 79, 6995–7004. [Google Scholar] [CrossRef]
- Kochhar, S.; Jacobs, D.M.; Ramadan, Z.; Berruex, F.; Fuerholz, A.; Fay, L.B. Probing gender-specific metabolism differences in humans by nuclear magnetic resonance-based metabonomics. Anal. Biochem. 2006, 352, 274–281. [Google Scholar] [CrossRef]
- Caterino, M.; Ruoppolo, M.; Villani, G.R.D.; Marchese, E.; Costanzo, M.; Sotgiu, G.; Dore, S.; Franconi, F.; Campesi, I. Influence of Sex on Urinary Organic Acids: A Cross-Sectional Study in Children. Int. J. Mol. Sci. 2020, 21, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, M.C.; Brennan, L.; Malthouse, J.P.; Roche, H.M.; Gibney, M.J. Effect of acute dietary standardization on the urinary, plasma, and salivary metabolomic profiles of healthy humans. Am. J. Clin. Nutr. 2006, 84, 531–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assfalg, M.; Bertini, I.; Colangiuli, D.; Luchinat, C.; Schafer, H.; Schutz, B.; Spraul, M. Evidence of different metabolic phenotypes in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 1420–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.Y.; Yeh, K.W.; Lin, G.; Chiang, M.H.; Yang, S.C.; Chao, W.J.; Yao, T.C.; Tsai, M.H.; Hua, M.C.; Liao, S.L.; et al. Metabolomics Reveals Dynamic Metabolic Changes Associated with Age in Early Childhood. PLoS ONE 2016, 11, e0149823. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rodriguez, J.A.; Fresnadillo Martinez, M.J. Dynamics of nasopharyngeal colonization by potential respiratory pathogens. J. Antimicrob. Chemother. 2002, 50 (Suppl. S2), 59–73. [Google Scholar] [CrossRef] [Green Version]
- Adegbola, R.A.; Obaro, S.K.; Biney, E.; Greenwood, B.M. Evaluation of Binax now Streptococcus pneumoniae urinary antigen test in children in a community with a high carriage rate of pneumococcus. Pediatr. Infect. Dis. J. 2001, 20, 718–719. [Google Scholar] [CrossRef]
- Tsai, M.H.; Liao, S.L.; Chiu, C.Y.; Shih, H.J.; Hua, M.C.; Yao, T.C.; Lai, S.H.; Yeh, K.W.; Chen, L.C.; Chang, Y.J.; et al. Longitudinal investigation of nasopharyngeal pneumococcal carriage in early childhood: The PATCH birth cohort study. PLoS ONE 2020, 15, e0237871. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The human urine metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Roberts, A.B.; Buffa, J.A.; Levison, B.S.; Zhu, W.; Org, E.; Gu, X.; Huang, Y.; Zamanian-Daryoush, M.; Culley, M.K.; et al. Non-lethal Inhibition of Gut Microbial Trimethylamine Production for the Treatment of Atherosclerosis. Cell 2015, 163, 1585–1595. [Google Scholar] [CrossRef] [Green Version]
- Hollister, E.B.; Riehle, K.; Luna, R.A.; Weidler, E.M.; Rubio-Gonzales, M.; Mistretta, T.A.; Raza, S.; Doddapaneni, H.V.; Metcalf, G.A.; Muzny, D.M.; et al. Structure and function of the healthy pre-adolescent pediatric gut microbiome. Microbiome 2015, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; Pan, Z.; Xi, B.; Hainline, B.E.; Shanaiah, N.; Asiago, V.; Gowda, G.A.; Raftery, D. 1H NMR metabolomics study of age profiling in children. NMR Biomed. 2009, 22, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.E.; Siskos, A.P.; Maitre, L.; Robinson, O.; Athersuch, T.J.; Want, E.J.; Urquiza, J.; Casas, M.; Vafeiadi, M.; Roumeliotaki, T.; et al. Determinants of the urinary and serum metabolome in children from six European populations. BMC Med. 2018, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Slupsky, C.M.; Cheypesh, A.; Chao, D.V.; Fu, H.; Rankin, K.N.; Marrie, T.J.; Lacy, P. Streptococcus pneumoniae and Staphylococcus aureus pneumonia induce distinct metabolic responses. J. Proteome Res. 2009, 8, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Waites, K.B.; Xiao, L.; Liu, Y.; Balish, M.F.; Atkinson, T.P. Mycoplasma pneumoniae from the Respiratory Tract and Beyond. Clin. Microbiol. Rev. 2017, 30, 747–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emwas, A.H.; Saccenti, E.; Gao, X.; McKay, R.T.; Dos Santos, V.; Roy, R.; Wishart, D.S. Recommended strategies for spectral processing and post-processing of 1D (1)H-NMR data of biofluids with a particular focus on urine. Metabolomics 2018, 14, 31. [Google Scholar] [CrossRef] [Green Version]
- Bernini, P.; Bertini, I.; Luchinat, C.; Nincheri, P.; Staderini, S.; Turano, P. Standard operating procedures for pre-analytical handling of blood and urine for metabolomic studies and biobanks. J. Biomol. NMR 2011, 49, 231–243. [Google Scholar] [CrossRef]
- Da Gloria Carvalho, M.; Pimenta, F.C.; Jackson, D.; Roundtree, A.; Ahmad, Y.; Millar, E.V.; O’Brien, K.L.; Whitney, C.G.; Cohen, A.L.; Beall, B.W. Revisiting pneumococcal carriage by use of broth enrichment and PCR techniques for enhanced detection of carriage and serotypes. J. Clin. Microbiol. 2010, 48, 1611–1618. [Google Scholar] [CrossRef] [Green Version]
- Carvalho Mda, G.; Tondella, M.L.; McCaustland, K.; Weidlich, L.; McGee, L.; Mayer, L.W.; Steigerwalt, A.; Whaley, M.; Facklam, R.R.; Fields, B.; et al. Evaluation and improvement of real-time PCR assays targeting lytA, ply, and psaA genes for detection of pneumococcal DNA. J. Clin. Microbiol. 2007, 45, 2460–2466. [Google Scholar] [CrossRef] [Green Version]
- Lacy, P.; McKay, R.T.; Finkel, M.; Karnovsky, A.; Woehler, S.; Lewis, M.J.; Chang, D.; Stringer, K.A. Signal intensities derived from different NMR probes and parameters contribute to variations in quantification of metabolites. PLoS ONE 2014, 9, e85732. [Google Scholar] [CrossRef]
- Stringer, K.A.; Serkova, N.J.; Karnovsky, A.; Guire, K.; Paine, R., 3rd; Standiford, T.J. Metabolic consequences of sepsis-induced acute lung injury revealed by plasma (1)H-nuclear magnetic resonance quantitative metabolomics and computational analysis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L4–L11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S. Quantitative metabolomics using NMR. Trends Anal. Chem. 2008, 27, 228–237. [Google Scholar] [CrossRef]
- McClay, J.L.; Adkins, D.E.; Isern, N.G.; O’Connell, T.M.; Wooten, J.B.; Zedler, B.K.; Dasika, M.S.; Webb, B.T.; Webb-Robertson, B.J.; Pounds, J.G.; et al. (1)H nuclear magnetic resonance metabolomics analysis identifies novel urinary biomarkers for lung function. J. Proteome Res. 2010, 9, 3083–3090. [Google Scholar] [CrossRef]
- Craig, A.; Cloarec, O.; Holmes, E.; Nicholson, J.K.; Lindon, J.C. Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal. Chem. 2006, 78, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- Laird, N.M.; Ware, J.H. Random-effects models for longitudinal data. Biometrics 1982, 38, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, G.M.; Laird, N.M.; Ware, J.H. Applied Longitudinal Analysis, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wickham, H. ggplot: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | All (n = 122) |
|---|---|
| Age Category | |
| >3 Months to 1 Year | 7 (5.7%) |
| >1 Year to 5 Years | 39 (32.0%) |
| >5 Years to 12 Years | 76 (62.3%) |
| Male (Sex) | 72 (59.0%) |
| Race | |
| White | 89 (73%) |
| African American | 23 (19%) |
| Other | 10 (8%) |
| Ethnicity | |
| Hispanic | 6 (5.0%) |
| Not Hispanic | 115 (95.0%) |
| Breast Fed (n = 12) | |
| Exclusively Breast-Fed | 9 (75.0%) |
| Exclusively Formula-Fed | 2 (16.7%) |
| Mixed | 1 (8.3%) |
| Sample also Obtained from Biological Sibling | 102 (83.6%) |
| Mother’s Education | |
| High School or Less | 8 (6.6%) |
| Some College or Associates | 31 (25.4%) |
| Bachelors | 26 (21.3%) |
| Masters | 43 (35.2%) |
| Professional or Doctoral | 14 (11.5%) |
| Asthma History | 11 (9.0%) |
| Seasonal Flu Vaccine Received | 68 (55.7%) |
| No Virus Detected | 93 (76.0%) |
| Virus Detected | 29 (24.0%) |
| Bocavirus | 1 (3.4%) |
| Coronavirus | 3 (10.3%) |
| Influenza | 1 (3.4%) |
| Rhinovirus/Enterovirus | 21 (72.4%) |
| Parainfluenza | 3 (10.3%) |
| Sp Load (copies/mL) * | 24,643 (3174, 496,021) |
| Sp Presence | |
| Positive | 24 (19.7%) |
| Negative ** | 98 (80.3%) |
| Metabolite | Predictor | Beta Estimate | Standard Error | % Change | p-Value | LRT p-Value | BH Corrected LRT p-Value |
|---|---|---|---|---|---|---|---|
| 1-Methylnicotinamide | (Intercept) | 2.58 | 0.33 | 1219.53 | <0.01 | 0.02 | 0.52 |
| Age (years) | −0.01 | 0.06 | −1.149 | 0.84 | |||
| Time | −0.07 | 0.14 | −7.048 | 0.61 | |||
| Asthma | −0.19 | 0.23 | −16.88 | 0.43 | |||
| Sp Presence | 0.41 | 0.15 | 50.349 | 0.01 | |||
| Rhinovirus/Enterovirus | −0.43 | 0.16 | −34.875 | 0.02 | |||
| Age–Time Interaction | −0.01 | 0.03 | −0.569 | 0.83 | |||
| 3-Hydroxybutyrate | (Intercept) | 2.15 | 0.31 | 758.009 | <0.01 | 0.03 | 0.52 |
| Age (years) | −0.06 | 0.05 | −5.681 | 0.28 | |||
| Time | −0.19 | 0.13 | −17.489 | 0.16 | |||
| Asthma | −0.28 | 0.2 | −24.681 | 0.18 | |||
| Sp Presence | 0.18 | 0.13 | 20.083 | 0.18 | |||
| Rhinovirus/Enterovirus | −0.18 | 0.15 | −16.204 | 0.24 | |||
| Age–Time Interaction | 0.01 | 0.03 | 1.024 | 0.68 | |||
| Trimethylamine-n-Oxide (TMAO) | (Intercept) | 4.21 | 0.46 | 6633.51 | <0.01 | 0.04 | 0.52 |
| Age (years) | −0.13 | 0.08 | −11.916 | 0.13 | |||
| Time | −0.35 | 0.18 | −29.622 | 0.06 | |||
| Asthma | −0.52 | 0.35 | −40.496 | 0.16 | |||
| Sp Presence | 0.61 | 0.23 | 83.097 | 0.02 | |||
| Rhinovirus/Enterovirus | 0 | 0.26 | 0.339 | 0.99 | |||
| Age–Time Interaction | 0.06 | 0.03 | 6.048 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambroggio, L.; Florin, T.A.; Williamson, K.; Pfefferman, C.; Wagner, B.D.; Yeomans, L.; Kim, J.H.; Sucharew, H.; Macaluso, M.; Ruddy, R.M.; et al. Respiratory Colonization and Short-Term Temporal Changes in the Urinary Metabolome of Children. Metabolites 2021, 11, 500. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080500
Ambroggio L, Florin TA, Williamson K, Pfefferman C, Wagner BD, Yeomans L, Kim JH, Sucharew H, Macaluso M, Ruddy RM, et al. Respiratory Colonization and Short-Term Temporal Changes in the Urinary Metabolome of Children. Metabolites. 2021; 11(8):500. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080500
Chicago/Turabian StyleAmbroggio, Lilliam, Todd A. Florin, Kayla Williamson, Cory Pfefferman, Brandie D. Wagner, Larisa Yeomans, Jae Hyun Kim, Heidi Sucharew, Maurizio Macaluso, Richard M. Ruddy, and et al. 2021. "Respiratory Colonization and Short-Term Temporal Changes in the Urinary Metabolome of Children" Metabolites 11, no. 8: 500. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11080500